











 texto em
texto em  Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroducción
El lulo, Solanum quitoense Lam. (Solanaceae) cultivar Castilla, es una de las frutas andinas con mayor potencial comercial, dada su amplia aceptación en los mercados nacionales, y por ser considerada un producto promisorio para los internacionales. Sin embargo, la demanda se ha reducido debido al sistema de producción que se utiliza, caracterizado por un alto consumo de pesticidas, que ocasionan que se detecten trazas de ingredientes activos no permitidos en los mercados externos (Muñoz, 2011).
Durante el año 2014 en Colombia se produjeron 33.522 toneladas de lulo, con rendimientos promedio de 9,6 t/ha al año. El departamento de Antioquia es el principal productor, con 10.477 t, que corres ponden al 31,25 % de la producción total, seguido por Huila, Cundinamarca y Santander.
En el periodo de enero a julio de 2014, los principales destinos de esta fruta fueron Francia, que participa con el 49 % de la demanda, con un valor de USD38.990,50, seguido de Panamá (14 %), España (11 %) y Aruba (4 %) (Departamento Administrativo Nacional de Estadística [DANE], 2015).
Algunos autores han reconocido dos variedades botánicas: Solanum quitoense var. septentrionale, que presenta espinas, y Solanum quitoense var. quitoense, que no las tiene (Fory et al., 2010; Morton, 1987; Schultes & Cuatrecasas, 1953; Whalen, Costich, & Heiser, 1981), aspecto sobre el cual hay controversia.
El lulo cv. La Selva es producto de dos retro-cruzamientos encaminados a una población sin espinas del híbrido obtenido por el profesor C. Heiser (Universidad de Indiana, EE. UU.) de Solanum quitoense χ Solanum hirtum (lulo de perro) (Bernal, Lobo, & Londoño, 1998), y se caracteriza por no poseer espinas ni en sus hojas y ni en el tallo. Es un cultivar precoz, que comienza su producción entre seis y siete meses después de la siembra.
Esa producción puede llegar a las 20 t/ha, y se destina a la agroindustria, debido a que tiene una vida muy corta en poscosecha, ya que su pulpa se oxida rápidamente y tiene una alta dehiscencia, que causa que el fruto se raje incluso antes de alcanzar su madurez total. Sin embargo, posee tolerancia a nematodos y al pasador del fruto (Medina, Lobo, Martínez, & Riaños, 2004).
El lulo de los cultivares Castilla y Castilla Larga Vida presenta espinas en tallos y hojas, inicia su producción entre siete y nueve meses después del trasplante, y alcanza producciones de 8 a 10 t/ha, que se destinan al mercado fresco. Entre las plagas que más afectan el cultivo se encuentran el pasador del fruto Neoleucinodes elegantalis (Guenée) (Lepidoptera: Crambidae), que constituye la principal, y el nematodo Meloidogyne incognita (Kofoid & White) (Nematoda: Secernentea: Tylenchida: Heteroderidae) (Bernal & Díaz, 2006).
Neoleucinodes elegantalis es una polilla que en su estado larval perfora frutos y los barrena, alimen tándose de ellos hasta llegar a su adultez. La larva ingresa al fruto a partir del tercer instar y permanece ahí hasta el quinto, luego se deja caer al suelo, donde empupa. Al salir, la larva deja un orificio que permite la entrada de otros insectos y microorganismos, deteriorando completamente el fruto (Instituto Colombiano Agropecuario [ICA], 2011; Marcano, 1991; Obando, 2011; Parra, Barona, & Vallejo, 1994).
Debido a sus particulares hábitos en los que la larva vive dentro del fruto, el control químico resulta inefectivo (Díaz-Montilla, 2013). Esta plaga es considerada cuarentenaria en Chile, EE. UU. y Perú, lo cual limita las exportaciones de fruta (Obando, 2011).
En cuanto al nematodo M. incognita, de acuerdo con Tamayo (2001) , se considera una plaga que ataca al lulo en todas las zonas productoras de Colombia. El ataque inicia en el semillero, produ ciendo agallas y nudos en las raíces, que impiden la absorción de agua y nutrientes, lo que conduce a plantas débiles, con síntomas como hojas de menor tamaño, clorosis de hojas bajeras y marchitamiento temporal en días soleados. Puede llegar a reducir la producción en más del 50 % (Franco et al., 2002).
El control a base de agroquímicos es el más usado por los agricultores, pero tiene una gran desventaja, que radica en que se utilizan productos extremada mente tóxicos y residuales, que contaminan el medioambiente y que, al ser aplicados de manera repetida, ocasionan resistencia de este organismo al producto, agravando aún más el problema (Andrés, 2002).
Por lo anterior, el mejor método de control para estas dos plagas limitantes del cultivo es el uso de cultivares y variedades resistentes. Se ha reportado que el lulo cv. La Selva es resistente a los nematodos y al insecto pasador del fruto N. elegantalis (Lobo & Medina, 2000). La definición de la resistencia varietal a las plagas es muy variada, pero, en términos agrícolas prácticos, se puede afirmar que un cultivar resistente es aquel que rinde más que uno susceptible cuando se enfrenta a la invasión de una plaga (Maxwell & Jennings, 1980).
Por consiguiente, la resistencia de las plantas es relativa, y se basa en la comparación con aquellas que carecen de esa cualidad, es decir, las susceptibles. Según Panda y Khush (1995) , la resistencia varietal tiene las siguientes características fundamentales: a) heredable y controlada por uno o más genes; b) relativa y solo puede ser medida al compararla con un cultivar susceptible de la misma especie; c) medible de forma cualitativa o cuantitativa; d) variable y con la posibilidad de ser modificada por factores bióticos y abióticos.
En su primera fase, este proyecto de investigación pretendió evaluar la resistencia o tolerancia varietal, a nematodos fitopatógenos y al pasador del fruto, de los materiales Fi obtenidos del cruzamiento recíproco entre el lulo cv. Castilla, selección natural Larga Vida, y el cv. La Selva.
Materiales y métodos
Colecta del material vegetal empleado como parental
El material vegetal del lulo cv. La Selva se colectó en la vereda Caucayá del municipio de Belén de Umbría, Risaralda (Colombia), con coordenadas geográficas 05°13' N, 75°48' O, a una altitud de 1.320 m s. n. m., con una temperatura promedio de 26 °C y una precipitación anual de 2.150 mm.
Se cortaron estacas de chupones basales de plantas seleccionadas por su excelente producción y buen estado fitosanitario. Las estacas fueron sembradas en bolsas plásticas, con un sustrato compuesto por tres partes de suelo más una de materia orgánica. Una vez sembradas, se realizó una aplicación en drench con Trichoderma spp., con el fin de prevenir el ataque del complejo damping-off (Fusarium oxysporum, Phytophthora infestans, Sclerotinia sclerotiorum) (ICA, 2011).
Para el caso de la selección natural de lulo cv. Castilla Larga Vida, se colectaron frutos de las mejores plantas de un cultivo comercial, aislado de otros, en la vereda Andica de Belén de Umbría, Risaralda, con una altitud de 1.370 m s. n. m., una temperatura promedio de 25 °C, una precipitación promedio anual de 2.030 mm, en las coordenadas geográficas 05°14' N, 75°52' O.
Las semillas se extrajeron de frutos con un peso mayor a 150 g y un diámetro superior a 8 centímetros, se mezclaron como una selección masal y se sembraron en germinador. 28 días después de la siembra, se observaron las primeras plántulas que emergieron, y se logró una germinación del 70,3 %. Cuando las plántulas emitieron el tercer par de hojas, fueron trasplantadas a bolsas plásticas, en las cuales permanecieron durante cuatro meses, hasta tener tres hojas verdaderas.
Manejo de plantas parentales
Las plántulas de ambos parentales se sembraron en un lote de la Universidad Nacional Abierta y a Distancia (UNAD) en el municipio de Dosquebradas, Risaralda, ubicado a una altitud de 1.460 m s. n. m., con una temperatura promedio de 21 °C, una preci pitación media anual de 2.200 mm y una humedad relativa promedio del 85 %.
Para la siembra se hicieron hoyos de 30 χ 30 χ 30 cm, en un arreglo en cuadro, con distancias de 2 m entre plantas y 3 m entre surcos. A cada hoyo se le adicionaron 1,5 kg de materia orgánica compostada y 150 g de micorriza comercial, con el fin de favorecer una mejor absorción de fósforo.
Se llevaron a cabo todas las labores culturales típicas del cultivo, como control de las arvenses, fertilización del suelo con NPK y elementos secundarios, y fertilización foliar de elementos menores. También se hicieron podas de formación, para eliminar los brotes o chupones del tallo que estuvieran por debajo de los 30 cm de altura.
Cruzamientos
Cuatro meses después del trasplante, ocurrió la floración. En ese momento, se seleccionaron las flores de pistilo largo en ambos materiales, y se realizaron cruces recíprocos, es decir, se emplearon los dos materiales como plantas madres y padres. Se emascularon las flores, retirando los estambres, y se procedió a polinizar las flores escogidas como hembras. En la figura 1 se pueden apreciar los frutos que fueron producto de estos cruzamientos, en proceso de desarrollo.

Figura 1 Frutos en desarrollo, obtenidos de los cruzamientos de lulo cv. Castilla Larga Vida x cv. La Selva.
De estos cruzamientos se obtuvieron frutos de los que se extrajo la semilla Fi (figura 2). Las semillas de todos los frutos Fi de cada cruce recíproco se mantuvieron separadas como una selección masal. Se sembraron 344 semillas de cada uno de estos cruces y el porcentaje de germinación fue del 73 %. Se seleccionaron 100 plantas Fi de cada cruce, las cuales se pasaron a bolsas plásticas, y luego una parte de ellas fue llevada a campo, para realizar el proceso de evaluación de caracteres morfoagronómicos.

Figura 2 Frutos de lulo Fi. a. cv. Castilla Larga Vida x cv. La Selva; b. cv. La Selva x cv. Castilla Larga Vida.
Las plantas Fi de los cruces del lulo Castilla χ La Selva, La Selva χ Castilla, y La Selva χ Larga Vida se sembraron en un lote ubicado en terreno de la uñad, en el municipio de Dosquebradas, Risaralda, a una altitud de 1.460 m s. n. m., con una temperatura promedio de 21 °C, una precipitación anual de 2.200 mm y una humedad relativa del 80 %. La zona corresponde a un bosque muy húmedo premontano (bmh-PM), según la clasificación de zonas de vida de Holdridge.
La siembra se hizo con un diseño completamente al azar, con 25 plantas de cada uno de los tratamientos, para un total de 100. Cada siete días se tomaron datos de los siguientes descriptores morfoagronómicos cuantitativos: altura de la planta, diámetro del tallo, volumen del fruto, número de hojas por semana, número de botones por cojín, número de cojines, número de frutos por racimo, número de racimos, y área foliar. De cada uno de los tratamientos, se arrancaron 10 plantas con raíz a los seis meses de sembradas, para evaluar el nivel de infestación de las raíces por nematodo.
Evaluación de la resistencia a los nematodos
Aislamiento de nematodos
Los cultivos puros de M. incognita se obtuvieron de raíces infectadas de lulo cv. Castilla y de la tierra circundante a ellas, mediante el método del embudo de Baermann.
Montaje del experimento para evaluación de resistencia a Meloidogyne incognita
Para evaluar la resistencia de las plantas F1 a los nematodos, se tomaron plántulas con cuatro o cinco hojas bien formadas, y se sembraron en tarros plásticos con capacidad de 2 kg, en un sustrato compuesto por tres partes de suelo y una de materia orgánica, sin desinfectar.
Se utilizó un diseño completamente al azar. La unidad experimental correspondió a 10 plantas, y la de muestreo fue una planta completa por tratamiento y por repetición. Los tratamientos correspondieron a la interacción de los cruzamientos por la inoculación con nematodos, así:
T1: Plantas Fi Castilla Larga Vida x La Selva, sin inoculación de nematodos
T2: Plantas Fi Castilla Larga Vida x La Selva, inoculadas con nematodos
T3: Plantas Castilla Larga Vida, inoculadas con nematodos (testigo)
T4: Plantas Fi La Selva x Castilla Larga Vida, inoculadas con nematodos
T5: Plantas Fi La Selva x Castilla Larga Vida, sin inoculación de nematodos
Para la inoculación de las plantas se hicieron cuatro orificios en el suelo dentro de cada maceta y alrededor del cuello de la raíz, donde se inocularon 50 cm3 de agua destilada que contenía aproximadamente 10.000 nematodos, entre adultos y huevos. Estas plantas se mantuvieron dentro del invernadero durante 60 días, a una temperatura promedio de 23 °C y un 85 % de humedad relativa.
Al final de este periodo, las plantas se extrajeron de sus macetas, y se evaluó la nodulación del sistema radicular, de acuerdo con una escala de 0 a 10, donde 0 correspondió a una planta sana sin nodulación, y 10 a una con un sistema radicular completamente nodulado. Las plantas se consideraron resistentes cuando el índice medio de nodulación resultó inferior a 2, y susceptibles cuando se superó ese valor. Las variables de respuesta fueron: nivel de infestación, largo de la raíz y peso fresco de la raíz.
Evaluación de resistencia al pasador del fruto
Multiplicación del Neoleucinodes elegantalis
Los adultos de N. elegantalis se obtuvieron de frutos de lulo (de un cultivar no determinado) recolectados en un cultivo de la zona, que poseían orificios de entrada y no de salida. Fueron llevados al laboratorio y allí se dispusieron de cuatro a cinco frutos dentro de tarros de plástico transparente con arena húmeda y pedazos de papel tela en el fondo (figura 3), para que, una vez que las larvas salieran de los frutos, iniciaran el proceso de empupamiento en las telas de papel y los adultos emergieran posteriormente. Los adultos obtenidos fueron sexados por el color de sus alas y la abertura genital.

Montaje del experimento de evaluación de tolerancia a Neoleucinodes elegantalis
Para evaluar la tolerancia de las generaciones Fi al pasador del fruto, se emplearon mangas entomológicas (40 x 15 cm), seleccionando una rama del tercio medio de la planta, con frutos en su primera fase de formación, en los cuales se introdujeron dos polillas macho y dos hembras de N. elegantalis.
Se realizaron observaciones diarias con el fin de registrar la ovoposición. Para la evaluación de tolerancia al pasador del fruto, se utilizó un diseño completamente al azar de las progenies de los cruces recíprocos de lulo cv. Castilla Larga Vida x cv. La Selva, y como testigo se emplearon plantas de Castilla Larga Vida. La unidad experimental correspondió a una planta, con su respectiva manga entomológica, y la de muestreo fue de 10 plantas por tratamiento.
Se emplearon cuatro descriptores cuantitativos: porcentaje de frutos afectados, número promedio de orificios de salida por fruto, porcentaje de frutos con orificios de salida y número promedio de larvas por fruto. Los datos obtenidos de la experimentación, tanto de la evaluación de resistencia a nematodos como aquella al pasador del fruto, se sometieron a un análisis de varianza (Anova) y pruebas de comparación de medias (Duncan) utilizando el programa estadístico SPSS V15.0.
Resultados
Evaluación de resistencia de los materiales F1 al nematodo Meloidogyne incognita
El análisis de varianza para las variables nivel de infestación, largo de la raíz y peso fresco de la raíz, en plantas en etapa de almácigo (60 días después de inoculación), en relación con la resistencia al nematodo M. incognita de las plantas Fi de los cruces recíprocos de lulo cv. La Selva x cv. Castilla Larga Vida, presentó diferencias altamente significativas entre los tratamientos.
Nivel de infestación de Meloidogyne incognita en fase de almácigo
Al analizar el nivel de infestación de la raíz por los nematodos en fase de almácigo, 60 días después de la inoculación, el tratamiento 1 (plantas F1 Castilla Larga Vida x La Selva, sin inocular) presentó un nivel de infestación de nematodos de 0. Los tratamientos 2 (plantas F1 Castilla Larga Vida x La Selva, inoculadas con nematodos) y 5 (plantas F1 La Selva x Castilla Larga Vida, sin inocular) mostraron un porcentaje bajo (2,5 %).
Por su parte, los tratamientos 3 (plantas cv. Castilla, inoculadas con nematodos [testigo]) y 4 (plantas F1 La Selva x Castilla Larga Vida, inoculadas con nematodos), fueron los más susceptibles, con un nivel de infestación del 4,2 % y el 4,7 %, respectivamente. El 50 % de las plantas del tratamiento 2 mostraron niveles entre 0 y 2 de infestación, y pudieron constituirse como líneas resistentes (figura 4).
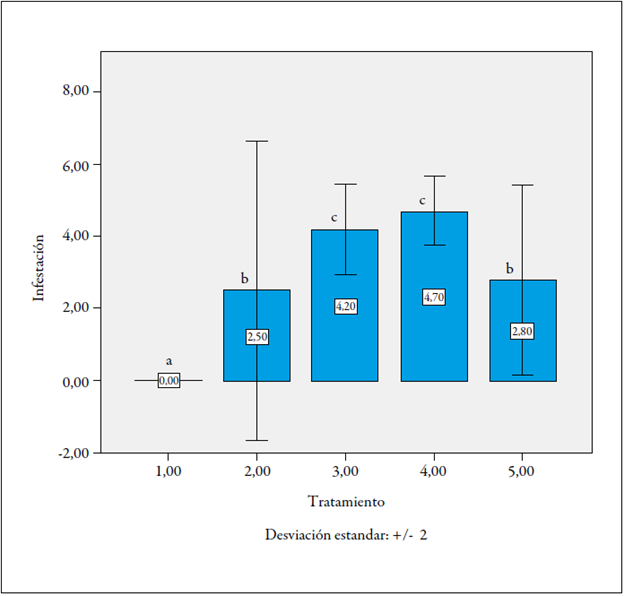
Largo de la raíz
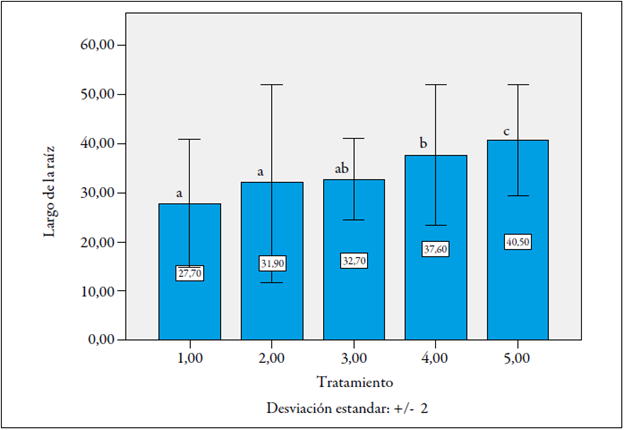
Se presentaron diferencias significativas entre los tratamientos, y las raíces del tratamiento 5 (F1 La Selva x Castilla Larga Vida, sin inocular) fueron las más largas, con un promedio de 40,5 cm de longitud, seguidas por las de este mismo material, pero inoculado (figura 5).
Fuente: Elaboracion propia
Figura 5 Afectación de la elongación de la raíz de las plantas F1 de lulo por la infestación de Meloidogyne incognita en fase de almácigo. Letras iguales sobre las barras significan que no difieren estadísticamente (p > 0,05).
Aunque para este caso se puede decir que los nematodos afectaron el alargamiento de la raíz, no fue así para el material Fi Castilla Larga Vida x La Selva, lo que puede deberse a un efecto oligogénico, aportado por el parental Castilla Larga Vida cuando se utiliza como macho (figura 5).
Peso fresco de la raíz
Por sí sola, la variable correspondiente al peso fresco de la raíz no resultó ser un buen indicador de la susceptibilidad o tolerancia al nematodo M. incognita, aunque en el tratamiento 4 (plantas La Selva x Castilla Larga Vida, inoculadas con nematodos) se encontró una diferencia altamente significativa, de 39,3 g respecto a todos los demás (figura 6).
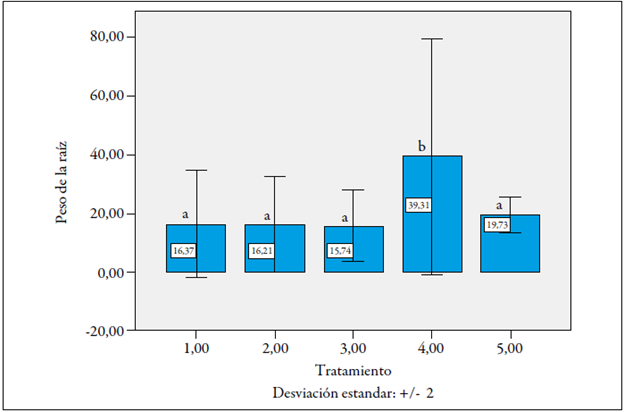
El mayor peso de las raíces de este tratamiento puede corresponder al efecto hiperplástico de la infección de M. incognita, que aumentó el volumen de la raíz frente a las que no estaban infectadas, como lo ha reportado la literatura (Hafez & Sundararaj, 2000; Hashmi, Huettel, Hammerschlag, & Krusberg, 1993; Perrotta, Magnano, & Bassi, 1980).
La figura 7 muestra el desarrollo de las plantas 60 días después de aplicado el tratamiento. En primer lugar, se evidencia un menor vigor de las plantas obtenidas de los cruces cuando se empleó el parental Castilla Larga Vida como madre; mientras que las progenies del cruce que tuvo a La Selva como madre presentaron un desarrollo más vigoroso.

Figura 7 Plantas de lulo creciendo en recipientes plásticos, representativas de cada uno de los cinco tratamientos, 60 días después de la inoculación.
El testigo, Castilla Larga Vida inoculado con M. incognita, además de un pobre desarrollo presentó amarillamiento, síntoma de la afectación por el nematodo. Aparentemente, los materiales F1 inoculados con M. incognita no mostraron sintomatología de tal afectación en el momento en que se evaluó la infestación de sus plantas.
De los resultados anteriores, resultantes de la comparación de los tratamientos T1 y T5, podemos concluir que no existe un efecto citoplasmático en la resistencia o tolerancia al nematodo, sino que es producto del efecto oligogénico, ya que se observa una mayor resistencia cuando se emplea el lulo cv. La Selva como parental macho.
Sin embargo, como lo afirma Dahms (1972) , es necesario tener en cuenta que este tipo de experimentos, en los que se ejercen altas presiones de inóculo, mediante la inoculación artificial de los tratamientos, pueden enmascarar los resultados, de manera que no se pueda detectar si estos materiales F1 realmente son resistentes.
Además, se debe considerar que, por tratarse de plantas normalmente alógamas, y al ser el cultivar La Selva el resultado de un retrocruzamiento, ambos parentales son heterocigotos, y se puede estar presentando una segregación en la progenie obtenida de los cruzamientos, lo que hace que cada planta presente niveles diferenciales de resistencia a estas plagas del cultivo.
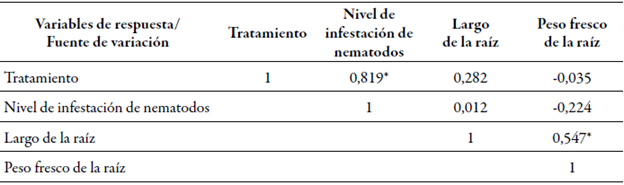
Al realizar un análisis de correlación entre las variables, se observa una relación inversa no significativa entre el nivel de infestación y el peso de la raíz, y una directa, no significativa, entre tal nivel y el largo de la raíz. Por su parte, la correlación entre el peso fresco de la raíz y su largo muestra una relación directa y altamente significativa, como es de esperarse según la lógica biológica del desarrollo de plantas. Sin embargo, es un dato que no da cuenta de la tolerancia o susceptibilidad a los nematodos (tabla 1).
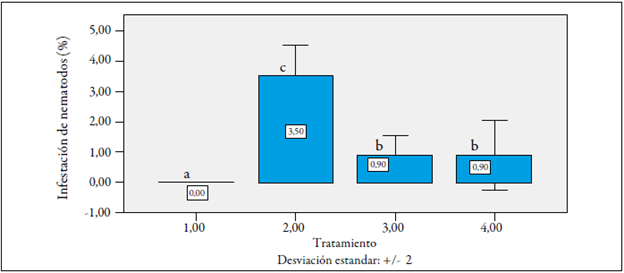
Nivel de infestación de Meloidogyne incognita en condiciones normales en campo
Las plantas F1 fueron evaluadas en condiciones normales en campo durante la fase de fructificación (seis meses después del trasplante en campo). Debido a que se trataba de una prueba destructiva, y a que se continuaba evaluando otras variables fenológicas de las plantas, se arrancaron al azar solo diez plantas de cada cruce recíproco y de los cultivares evaluados, y se encontraron diferencias estadísticas altamente significativas entre los materiales en lo que respecta a la infestación de M. incognita.
En función de las comparaciones entre las medias de los materiales evaluados en lo que respecta a la infestación de las raíces con M. incognita, se encontró que el grado de infestación de ambos cruces recí procos fue inferior a 1 %, lo que nos indica un alto nivel de resistencia de estos materiales a los nematodos, al igual que cv. La Selva (0). Por otro lado, cv. Castilla Larga Vida presentó una alta susceptibilidad (3,5 %) (figura 8).
Evaluación de la resistencia a Neoleucinodes elegantalis
Los materiales F1 de ambos cruces recíprocos de lulo cv. La Selva x cv. Castilla Larga Vida no presentaron larvas de N. elegantalis dentro de los frutos, ni orificios de salida de estas, mientras que aquellos de las plantas del cv. Castilla Larga Vida, utilizado como testigo, fueron afectados en un 60 %, y mostraron, en promedio, dos orificios de salida y dos larvas por fruto (tabla 2).
Tabla 2 Valores promedio de los descriptores evaluados para determinar la resistencia al pasador del fruto, Neoleucinodes elegantalis, de las plantas F1 de cruces de lulo cv. La Selva × cv. Castilla Larga Vida
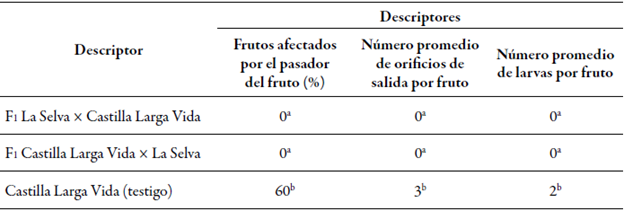
Promedios con letras distintas en la misma columna son significativamente diferentes.
Fuente: Elaboración propia
El alto porcentaje de frutos afectados y el elevado promedio de larvas que atacaron cada fruto en el tratamiento testigo indican que, utilizando las mangas entomológicas, el nivel de infestación fue adecuado para evaluar la resistencia al pasador en los materiales Fi de los cruces recíprocos de cv. La Selva x cv. Castilla Larga Vida.
Los materiales F1 presentaron resistencia al pasador del fruto, probablemente debido a un efecto antixe-nótico en el insecto adulto de N. elegantalis, basado sobre todo en factores físicos y químicos, causados por la presencia de tricomas en toda la planta y en especial en los frutos, que secretan terpenoides, alcaloides, sustancias fenólicas y otras, que pueden ser repelentes olfatorios o gustativos para el insecto (Aragão, Dantas, & Gandolfi, 2000; Cardona & Mesa, 2011; Simmons & Gurr, 2005). Sin embargo, se hace necesario continuar con las observaciones del cultivo en condiciones comerciales.
Comportamiento morfoagronómico de los híbridos
Las plantas híbridas de los cruces recíprocos resultaron ser vigorosas, con una altura promedio de 145 cm y un diámetro de tallo de 4,5 cm, una buena estructura, hojas grandes, un largo de 40,6 cm, un ancho de 41,5 cm, y un peciolo de 14 cm, y adaptadas a plena exposición solar.
La primera floración se inició 60 días después del trasplante, con un promedio de cuatro inflorescencias por planta, que formaron unos 13 frutos, con un diámetro promedio de 5,6 cm, semejantes al lulo cv. Castilla Larga Vida. La maduración y cosecha de los frutos sucedió 155 días después del inicio de la floración.
La epidermis de los frutos fue de color amarillo y la pulpa de color verde, sin dehiscencia (rajamiento), y con aroma y sabor agradables. Sin embargo, presentaron abundantes espinas pronunciadas, en tallos, ramas y hojas (figura 9), una condición que es catalogada como desfavorable para los productores, ya que dificulta las labores de cultivo.

Figura 9 Plantas F1, cuatro meses y medio después del trasplante, a libre exposición solar. a. Cruce de lulo cv. La Selva x cv. Castilla Larga Vida: se observan tallo y hojas con espinas muy fuertes, en gran cantidad y con ramificaciones basales; b. Cruce de lulo cv. Castilla Larga Vida x cv. La Selva: presenta espinas en tallo y hojas.
Conclusiones
Al ser inoculadas con un alto nivel de infestación del nematodo M. incognita, las plantas Fi obtenidas de los cruces recíprocos entre los parentales de lulo cv. Castilla Larga Vida y cv. La Selva resultaron ser susceptibles. Sin embargo, al evaluar estos mismos materiales en condiciones de campo, donde mediante la toma de muestras previas se había detectado la presencia del nematodo, las plantas presentaron una nodulación inferior al 1 %, lo que indica que los materiales son altamente tolerantes al nematodo, y que se presenta una posible segregación de este carácter de resistencia.
En la fase de almácigo, el peso fresco de la raíz de las plantas inoculadas fue afectado por la infestación de M. incognita, causando una reducción de su peso, lo que repercute en el desarrollo normal de la planta.
Aunque el crecimiento de la raíz se vio afectado significativamente en las plantas sometidas a una alta presión de inóculo de M. incognita, se evidenciaron un vigor superior y una mayor tolerancia en las plantas Fi, sobre todo cuando el material cv. Castilla Larga Vida se empleó como madre.
Aparentemente, las plantas Fi de los cruces recíprocos de cv. La Selva x cv. Castilla Larga Vida mostraron resistencia, debido a un efecto antixenótico en el pasador del fruto, N. elegantalis.
En ensayos posteriores, es necesario analizar por separado el peso fresco de raíz y el de nódulos de M. incognita, con el fin de determinar las reales diferencias en peso atribuibles a la nodulación causada por los nematodos.
Se requiere necesario continuar con las evaluaciones tanto de la tolerancia a nematodos como de la calidad de los frutos (grados Brix, sólidos totales, ácido ascórbico, etc.), en condiciones de cultivo comercial, con el objetivo de obtener materiales sin espinas, que conserven estas características agronómicas sobresalientes y que tengan una gran aceptación de los consumidores.

