










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCiencia y Tecnología Agropecuaria
Print version ISSN 0122-8706
Corpoica cienc. tecnol. agropecu. vol.19 no.3 Mosquera Sep./Dec. 2018
https://doi.org/10.21930/rcta.vol19_num3_art:566
Sistemas de información, zonificación y georreferenciación (agricultura de precisión)
Meloidogyne exigua (Meloidogynidae) em clones de seringueira em duas épocas do ano
1 Doutor, Universidade Estadual Paulista Júlio de Mesquita Filho (Unesp), Faculdade de Ciências Agronômicas (FCA). Botucatu, Brasil. E-mail: jpezzoni@gmail.com.
2 Doutora, Universidade Estadual Paulista Júlio de Mesquita Filho (Unesp), Faculdade de Ciências Agronômicas (FCA). Botucatu, Brasil. E-mail: andressa_brida23@hotmail.com.
3 Doutor, Universidade Estadual Paulista Júlio de Mesquita Filho (Unesp), Faculdade de Ciências Agronômicas (FCA). Botucatu, Brasil. E-mail: die.gomoraes45@gmail.com.
4 Professora Adjunta, Universidade Estadual Paulista Júlio de Mesquita Filho (Unesp), Faculdade de Ciências Agronômicas (FCA). Botucatu, Brasil. E-mail: srenata@fca.unesp.br.
5 Professora Adjunta III, Universidade Estadual Paulista Júlio de Mesquita Filho (Unesp), Faculdade de Ciências Agronômicas (FCA). Botucatu, Brasil. E-mail: czimback@gmail.com.
6 Livre-docente, Universidade Estadual Paulista Júlio de Mesquita Filho (Unesp), Faculdade de Ciências Agronômicas (FCA). Botucatu, Brasil. E-mail: elfurtado@fca.unesp.br.
Meloidogyne exigua Goeldi, raça 3 (Nematoda: Tylenchida: Meloidogynidae), tem se tornado importante para plantios de seringueira. Assim o objetivo deste trabalho foi monitorar sua infestação em plantio de seringueira e posteriormente realizar a distribuição espacial da população nas raízes dos clones GT 1 e RRIM 600, sem identificação do porta-enxerto, em duas épocas de 2013, no município de Itiquira, MT, Brasil. Em maio foram coletadas 264 e em novembro 227 amostras na área total de 6.646 hectares plantados com 22 clones, utilizados para a interpolação pelo método da krigagem, para elaboração do mapa de infestação na área total. Baseado neste mapa do plantio, foram recortados os talhões dos dois clones avaliados, representados com 55 amostras em maio (clone GT 1 = 29 e RRIM 600 = 26 amostras) e 52 em novembro (clone GT 1 = 27 e RRIM 600 = 25 amostras), em área total de 2.409 hectares. Com um aparelho GNSS foi identificado cada ponto amostral, coletando-se raízes da seringueira a 0,10 m de profundidade. As infestações em maio, nos dois clones, foram maiores do que em novembro. Os Índices de Dependência Espacial (IDE) entre as coletas realizadas em maio e novembro de 2013 foram moderados. A validação cruzada dos dados não foi satisfatória, devido a alta variação de infestações.
Palavras-chave: geoestatística; Hevea brasiliensis; interpolação; krigagem; nematoide das galhas
Meloidogyne exigua Goeldi, race 3 (Nematoda: Tylenchida: Meloidogynidae), has become an important pest for rubber tree plantations. Thus, the objective of this work was to monitor its infestation in rubber tree plantations and to determine the spatial distribution of the population in the roots of clones GT 1 and RRIM 600, without identification of the rootstock, in two periods of 2013, in the municipality of Itiquira, MT, Brazil. In May, 264 samples were collected, and in November, 227 samples were collected in the total area of 6,646 hectares planted with 22 clones, which were used for the interpolation by the kriging method for mapping infestations in the total area. Based on this planting map, the plots of the two evaluated clones were cut, resulting in 55 samples in May (clone GT 1 = 29 and RRIM 600 = 26 samples) and 52 in November (clone GT 1 = 27 and RRIM 600 = 25 samples), in a total area of 2,409 hectares. With a GNSS device, each sampling point was identified, and roots of the rubber tree were collected at 0.10 m depth. For the two clones, the infestations in May were larger than those recorded in November. The Spatial Dependency Index (IDE) between collections collected in May and November of 2013 were moderate. The cross-validation of the data was not satisfactory, due to the high variation of infestations.
Keywords: geostatistics; Hevea brasiliensis; interpolation; kriging; root-knot nematode
Introdução
A seringueira Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg. (Euphorbiaceae) é uma planta de ciclo perene, de origem tropical, cultivada e utilizada de modo extrativo, com a finalidade de produção de borracha natural (Campelo-Júnior, 2000). Sua exploração econômica é baseada na sangria para a extração do látex, onde os vasos laticíferos da casca são atingidos, liberando a borracha natural (Rajagopal, Vijayakumar, Thomas, & Karunaichamy, 2004). Em plantios comerciais, de forma geral, a enxertia é o método mais utilizado na propagação de mudas, visando manter a integridade genotípica dos clones (Cardinal, Gonçalves, & Martins, 2007a).
A interação entre os porta-enxertos GT 1, IAN 873, PB 235, RRIM 600, RRIM 701 e sementes não selecionadas influenciaram na produção de borracha seca dos enxertos/ clones de copa GT 1, IAN 873, PB 235, PR 107, RRIM 600 e RRIM 701 (Cardinal, Gonçalves, & Martins, 2007b). Pesquisas detalhando esta relação do ponto de vista de metabolismo radicular até o momento não foram realizadas. Desta forma, baseado nesta interação citada, as raízes do porta-enxerto provavelmente podem ser influenciadas pelo enxerto e consequentemente refletir na infestação de patógenos de solo.
A planta cultivada em monocultura tornou-se uma espécie vulnerável a diferentes microorganismos, tais como os fungos Microcyclus ulei, Colletotrichum gloeosporioides e C. acutatum, Oidium hevea e Tanatephorus cucumeris (Furtado, 2010), e o nematoide das galhas - Meloidogyne exigua (Santos, 1995).
Espécies do gênero Meloidogyne são caracterizadas pela indução do engrossamento nas raízes denominadas de galhas, resultando em baixa produção de sementes, crescimento reduzido, murchamento e aumento da susceptibilidade a outros patógenos (Randig, Bongiovanni, Carneiro, Sarah, & Castagnone-Sereno, 2002). No Brasil, a espécie com maior importância para a seringueira é o M. exigua Goeldi (Santos, Matos, Barré, & Ferraz, 1992). Muniz et al. (2008) classificaram este nematoide em raça 1: infecta o pimentão e o cafeeiro; raça 2: infecta o tomateiro, o pimentão e o cafeeiro; e a raça 3: parasita somente a seringueira.
Devido à população do nematoide apresentar distribuição aglomerada (Ferris & Wilson, 1987), a metodologia da geoestatística pode ser utilizada, na qual o princípio fundamental é que, teoricamente, amostras próximas no tempo e espaço sejam mais similares entre si, do que as que estiverem distantes (Isaaks & Srivastava, 1989). A geoestatística apresenta as seguintes etapas: 1º - levantamento do variograma experimental; 2º - ajuste dos variogramas e validação cruzada do modelo a ser utilizado na Krigagem; 3º - interpolação pela Krigagem (Landim, 2006).
A distribuição espacial de nematoides ocorre influenciada pela presença de alimento, determinada pela fisiologia vegetal do hospedeiro (Yeates, 1987). Análises espaciais de nematoides foram realizadas por Webster e Boag (1992), Wallace e Hawkins (1994) e Farias, Barbosa, Vieira, Sánchez-Vila e Ferraz (2002), onde os resultados demonstraram relação espacial entre populações. O presente trabalho objetivou realizar a análise da infestação de M. exigua raça 3 em plantio comercial de seringueira e posteriormente encontrar a distribuição espacial da população nas raízes dos clones de copa GT 1 e RRIM 600, em duas épocas do ano.
Material e métodos
Área de estudo
A área de estudo encontra-se no município de Itiquira, MT, Brasil, em área total de 6.646 ha plantados com 22 clones de seringueira, com idades entre 12 e 34 anos no ano de 2013, nas coordenadas geográficas de 17°17'40,29" e 17°26'17,73" de latitude sul e 54°44'28,6" e 54°44'43,55" de longitude oeste, na bacia do Rio Paraguai. Posterior ao mapeamento da população de nematoides em todos os 22 clones, o mapa total foi recortado baseado nos talhões dos clones GT 1 e RRIM 600. O clone/ enxerto GT 1, apresentou área total de 1.529 ha (maio) e 1.441 ha (novembro); e o RRIM 600 com 880 ha nos dois meses. O local apresenta solo classificado como Latossolo Vermelho Distrófico típico - LVd, de acordo com a Empresa Brasileira de Pesquisa Agropecuária (Embrapa) (2013). Segundo Köppen (1923), o clima da região é classificado como Aw com inverno seco, entre junho e agosto. Devido ao histórico de infestação pelo nematoide M. exigua raça 3, o plantio foi avaliado para verificar seu nível populacional nos clones estudados.
Coleta de amostras e análises nematológicas
As coletas amostrais em maio e novembro foram definidas baseadas na fenologia da planta durante o ano. Em maio a planta está no fim do período de máximo potencial produtivo e este mês precede a desfolha natural anual das plantas, que regionalmente ocorre entre maio e julho. O período chuvoso e quente do ano anterior a maio pode favorecer o aumento da população de nematoides. Em novembro, período posterior à desfolha anual, a planta está totalmente reenfolhada e a população de nematoides possivelmente é reduzida.
Nos meses de maio e novembro de 2013 foram coletados 264 e 227 pontos amostrais, respectivamente, para os 22 enxertos, sendo que todos estes dados foram utilizados para a interpolação pela krigagem. O número de pontos amostrais do clone GT 1 foi de 29 em maio e 27 pontos em novembro e para o RRIM 600 foram 26 em maio e 25 amostras em novembro. Os porta-enxerto dos clones não foram identificados. Os pontos amostrais foram distribuídos em toda a área experimental em grade irregular, devido a infestação ocorrer em toda área de plantio. Em cada ponto amostral foram coletadas raízes da seringueira a uma profundidade de 0,10 m. Para registro, localização e definição dos pontos amostrais, foi utilizado um aparelho GNSS com precisão de 4 a 5 m, datum WGS 84, zona 21 Sul e projeção UTM.
A análise nematológica das amostras de raízes foi executada conforme a metodologia proposta por Coolen e D’Herde (1972), considerando-se o número de indivíduos em 10 g de raízes. Após o processamento das amostras, foram efetuadas as contagens dos nematoides na lâmina de Peters e estimada a população na amostra proporcionalmente ao volume da sus-pensão da amostra total.
Análise estatística e geoestatística
A estatística descritiva dos dados de infestação de nematoides foi realizada, onde a média, desvio padrão, variância amostral, coeficiente de assimetria e curtose foram calculados. Castrignanò (2011) citou que em caso de os dados apresentarem coeficiente de assimetria positivo e maior ou igual a 0,5, estes devem ser transformados.
Em seguida, realizou-se a análise variográfica dos dados baseada na hipótese intrínseca e apresentada na Equação 1, segundo Vieira, Hatfield, Nielsen e Biggar (1983). Para tanto foi efetuado o ajuste do modelo experimental e teórico, obtendo-se os valores dos componentes do variograma: alcance (a), patamar (C0 + C), componente estrutural (C) e efeito pepita (C0).

Onde: γ (h) - semivariância estimada entre pares de pontos; N(h) - número de pares de valores medidos; Z(xi), Z(xi + h) - são os pares de pontos separados por uma distância h.
A interpolação, por meio da krigagem ordinária, foi realizada para a área total (6.646 ha em maio e 5.604 ha em novembro) desconsiderando as diferenças de clones entre talhões. Landim e Sturaro (2002) citaram que a krigagem ordinária utiliza informações a partir do variograma para encontrar os pesos ótimos a serem associados às amostras com valores conhecidos que irão estimar pontos desconhecidos. Para a krigagem ordinária, foram realizados os cálculos descritos nas equações 2 e 3, conforme Thompson (1992):
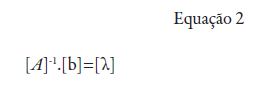
[A]-1: matriz inversa de semivariância entre as localidades da vizinhança de um ponto, determinada pelo modelo de variograma com base nas distâncias euclidianas entre as localidades; [b]: matriz de semivariância entre as localidades vizinhas (com a variável estimada) e o ponto para o qual a variável será interpolada, também determinado pelo modelo de variograma, com base nas distâncias entre as localidades vizinhas e o ponto a ser interpolado; [λ]: matriz de pesos de krigagem.
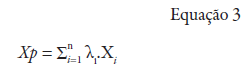
Xp: variável interpolada; λ1: peso da i-ésima localidade vizinha; Xi: valor da variável para a i-ésima localidade; n: número de localidades vizinhas empregadas para interpolação do ponto.
Posteriormente, foi elaborado o mapa de distribuição espacial de M. exigua raça 3 na área total e, realizado o recorte dos talhões com os clones de interesse. As infestações de nematoides no presente estudo foram divididas em cinco classes, sendo que a maior abrangência de infestações foi de 0 até 100.000, dividindo-se em 4 classes e deste valor até 200.000 nematoides foi a última classe. Devido ao alto nível populacional encontrado em todos os clones e nas duas épocas do ano, as diferentes classes populacionais foram distinguidas nas seguintes classes: I (0 a 25.000 nematoides), II (25.001 a 50.000 nematoides), III (50.001 a 75.000 nematoides), IV (75.001 a 100.000 nematoides) e V (100.001 a 200.000 nematoides), com unidade utilizada de nematoides em 10 g de raízes.
Resultados e discussão
Análise descritiva
Os resultados referentes à estatística descritiva para M. exigua raça 3 nos meses de maio e novembro de 2013 encontram-se na tabela 1. A média (m) de infestação encontrada em maio foi de 41.401,21 nematoides, enquanto que no mês de novembro foi de 5.566,737 nematoides (tabela 1), demonstrando queda acentuada nas populações, fato explicado pela desfolha anual da seringueira, chamado de hibernação por Moraes (1982). A deficiência hídrica ocorrida no período de hibernação também influencia a infestação, pois interfere no metabolismo do nematoide e no sistema radicular da seringueira.
Tabela 1 Resultados da estatística descritiva para as infestações de M. exigua raça 3 nos meses de maio (05) e novembro (11) de 2013
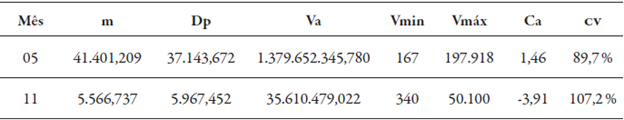
m = média; Dp = desvio padrão; Va = variância amostral; Vmin = valores mínimos e Vmáx = valores máximos de nematoides em 10 g de raízes; Ca = coeficiente de assimetria; CV = coeficiente de variação.
Fonte: Elaboração própria
O coeficiente de variação (CV) encontrado em maio foi de 89,7 % e em novembro foi de 107,2 %, valores que demonstram a elevada amplitude das infestações pontuais encontradas nestes meses. O coeficiente de assimetria dos dados em maio apresentou valor positivo, maior que 0,5, sendo aplicada a transformação log de x. Em novembro, o coeficiente de assimetria apresentou valor de - 3,91, portanto, não ocorreu a necessidade de transformação dos dados. Nos dois meses utilizou-se a metodologia de Castrignanò (2011).
Validação cruzada das estimativas
Esta análise dos dados foi realizada para posteriormente escolher o melhor ajuste e efetuar a interpolação pela krigagem. Segundo Isaaks e Srivastava (1989), a validação cruzada (VC) tem por função a avaliação de erros de estimativa, onde são comparados valores estimados com os amostrados (coeficiente de determinação - R2). Desta forma, certo valor em determinado ponto geográfico é temporariamente descartado do conjunto de dados e neste local se estima seu valor pela krigagem, utilizando o restante das amostras.
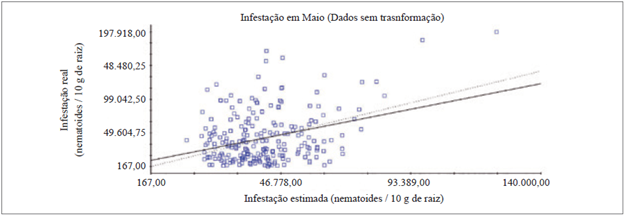
Em maio foram utilizados os dados brutos, pois com a transformação em log de x não foi possível efetuar o ajuste do variograma. Com os dados brutos, ocorreu muita discrepância entre as estimativas, devido a amplitude alta das infestações, sendo o fator responsável pelo baixo coeficiente de determinação - R2 = 0,104 (figura 1) da VC e com valor de coeficiente de regressão igual a 0,805.
Em novembro, os dados utilizados para a VC foram transformados em log de x para o ajuste do variograma, pois com os dados brutos não foi possível realizar o ajuste. Na VC foi obtido um valor de R2 = 0,109, sendo considerado baixo, devido a grande quantidade dos valores estimados que foram discrepantes em relação à infestação real (figura 2) e o valor de coeficiente de regressão foi igual a 0,618.
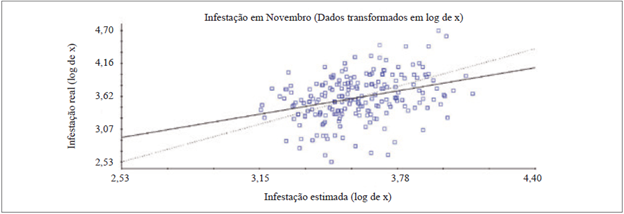
A validação cruzada dos dados de maio e novembro demonstraram a discrepância entre os dados de infestações estimadas e reais, devido ao fato de determinada porcentagem dos dados estimados pela krigagem serem inferiores ou superiores ao valor real de infestação (figuras 1 e 2). Este fato pode ser correlacionado com os valores altos de CV, com 89,7 % e 107,2 %, em maio e novembro, respectivamente (tabela 1), pois dentro do mesmo clone de copa houve intervalos de infestação muito altos, que consequentemente dificultam na validação cruzada.
Análise variográfica
Os modelos utilizados foram o exponencial e esférico, para os meses de maio e novembro, respectivamente. Apesar de o coeficiente de assimetria (Ca) para maio ter apresentado um valor de 1,46 (tabela 1) que culminou na transformação dos dados, a transformação não foi considerada, uma vez que o variograma não apresentou ajuste adequado quando aplicada a transformação. Portanto, o variograma de maio foi obtido considerando os dados originais (tabela 2).
Tabela 2 Análise variográfica da infestação de M. exigua em maio e novembro de 2013
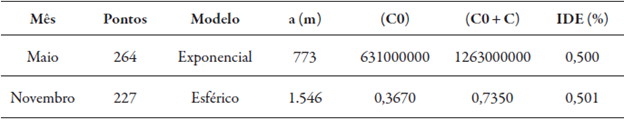
Maio - dados não foram transformados para log de x; Novembro - transformou-se para logaritmo. A = alcance; C0 = efeito pepita; C0 + C = patamar; IDE (%) = Índice de Dependência Espacial
Fonte: Elaboração própria
Dentre os componentes dos variogramas, o alcance (a) de maio e novembro foi de 773 e 1.546 metros, respectivamente. O menor alcance foi em maio (tabela 2), onde, a explicação mais aceita, foi devido ao amplo intervalo de infestação em pontos próximos, ou seja, locais geograficamente próximos tiveram populações com variações altas, superior a 10.000 indivíduos em 10 g de raízes entre duas amostras vizinhas, o que se refletiu nos dados da estatística descritiva relacionados na tabela 1.
Em novembro, a diferença de infestação entre pontos próximos foi menor do que em maio, pois porcentagem considerável das amostras, valor de 87,23 % das infestações encontradas em novembro foram de 0 até 10.000 nematoides em 10 g de raízes. Para as condições experimentais encontradas na avaliação, as amostragens realizadas a mais de 773 metros, em maio, e 1.546 metros, em novembro (tabela 2), não tiveram influência da distância entre si e, consequentemente, na confecção do mapa final da infestação.
Na análise variográfica, os alcances encontrados (773 m em maio e 1.546 m em novembro) demonstraram que, quanto maior a população de M. exigua raça 3, maior a influência na análise espacial da infestação, sendo que este componente do variograma (alcance) tem significativa importância, conforme citaram Journel e Huijbregts (1978). Estes autores descreveram que na interpolação por krigagem a grande influência no grau de confiança estatística da modelagem variográfica é a distância entre os pontos amostrais.
No presente estudo, o efeito pepita (C0) foi alto, o qual pode ser explicado devido à distância entre os pontos coletados apresentarem metragem alta, onde algumas foram maiores que 10 km. Este fato ocorreu devido à extensão do plantio, onde a distância entre os extremos dos talhões dos clones GT 1 e RRIM 600, chegou a 13,05 km. Outro fator que pode ter influenciado advém da alta amplitude dos dados de infestações.
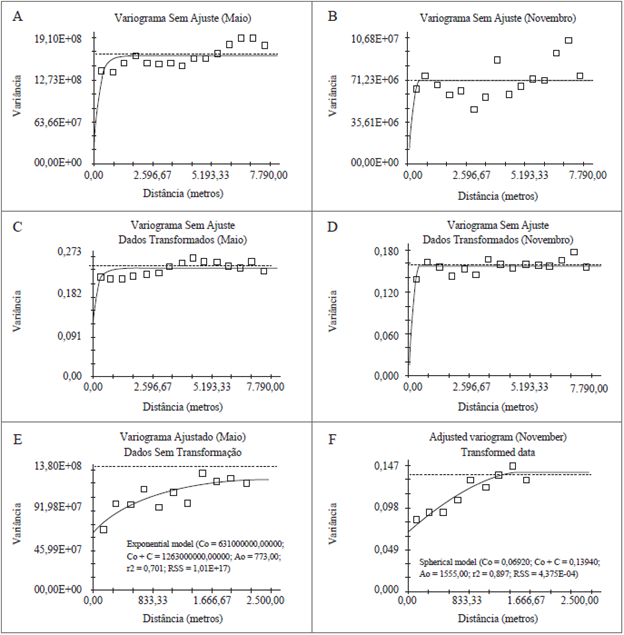
O efeito pepita (C0) de 45,45 % e 54,57 %, em maio e novembro respectivamente, demonstraram que o efeito de local foi praticamente igual à aleatoriedade dos dados (figura 3E e 3F). Estas porcentagens encontradas demonstram que em futuras coletas amostrais na mesma área, recomenda-se realizar a grade amostral com distâncias menores entre pontos, para ocorrer a mensuração mais aproximada da infestação real, obtendo assim maior acurácia. O efeito pepita ocorre devido a fatores incontroláveis, conforme Landim (2003), os quais são erros de mensuração e também devido à coleta dos dados não terem sido realizadas em espaçamentos menores, sem ocorrer o diagnóstico real do comportamento espacial do fenômeno.
Fonte: Elaboração própria
Figura 3 Variogramas sem ajuste com os dados sem transformação: Variograma - Maio (A) e Novembro (B); Variograma com dados transformados em log de x - Maio (C) e Novembro (D); e Variogramas ajustados em Maio e Novembro (F).
Em novembro, a transformação logarítmica dos dados enquadrou-se em um intervalo satisfatório, ajustando-se o variograma com os dados em log de x (figura 3F). Os modelos utilizados foram o exponencial e esférico, para os meses de maio e novembro, respectivamente, sendo que o modelo esférico apresentou uso adequado, conforme Trangmar, Yost, Wade, Uehara e Sudjadi (1987) e Paz, Taboada e Gómez (1996), sendo este o mais recomendado para demonstrar o comportamento de variogramas de atributos de plantas e de solos, onde foi encontrado na presente pesquisa (tabela 2). Webster e Boag (1992) verificaram que o modelo esférico foi o que melhor representou a distribuição espacial dos nematoides Heterodera avenae e Globodera rostochiensis. Wallace e Hawkins (1994) também utilizaram o modelo esférico para infestações do nematoide Tylenchorhynchus spp. em gramínea forrageira.
O patamar (C0 + C) de novembro apresentou estabilização do modelo variográfico mais próximo a variância amostral (s2) do que em maio, devido ao fato de os dados não serem transformados para log de x e apresentarem efeito pepita alto. O patamar demonstra a distância entre os pontos amostrais que tem influência entre si, conforme citou Vieira (2000), onde quando aumenta a distância entre os pontos amostrais, a variância também aumenta até um valor máximo no qual se estabiliza.
Cambardella et al. (1994) citaram que o patamar (C0 + C) têm importância na determinação do alcance (a), ou seja, componente que limita a dependência e a independência espacial entre as amostras (amostras com e sem influência entre si). O raio utilizado no procedimento de krigagem foi de 2.319 e 1.546 metros para a infestação em maio e novembro, respectivamente. Neste cálculo o número máximo de amostras vizinhas utilizadas para a interpolação foi de 16, podendo ocorrer menor número de amostras conforme a região geográfica do cálculo geoestatístico. Desta forma, ocorreu o máximo de abrangência do efeito estacionário do fenômeno de infestação por M. exigua raça 3.
O Índice de Dependência Espacial (IDE) apresentou em maio e novembro valores de 0,500 % e 0,501 % (tabela 2), respectivamente, onde, segundo a classificação de Zimback (2001), os IDE encontrados foram moderados. Os variogramas sem ajuste, com os dados sem transformação e transformados para log de x (figuras 3A, 3B, 3C, 3D) foram obtidos com a utilização de distancias entre pontos amostrais iguais a 7.790,00 m entre amostras e intervalo de cálculo entre pontos amostrais de 519,00 m, para os meses de maio e novembro.
Distribuição espacial da infestação
O mapa de distribuição espacial da infestação no clone GT 1 apresentou a máxima população com 159.840 nematoides (cor vermelha no mapa) e a menor população foi 2.640 nematoides M. exigua raça 3 (cor azul), sendo ambas no mês de maio. Em novembro a população foi classificada somente na primeira classe de infestação, cor azul, entre 0 e 25.000 nematoides, variando de 300 até 13.860 nematoides (figura 4).
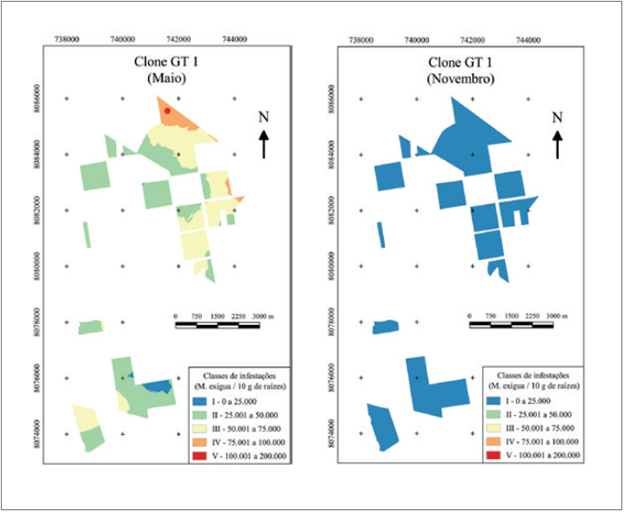
O clone GT 1 demonstrou dados na avaliação de maio semelhantes ao exposto por Bernardo et al. (2003), onde encontraram infestação de 61.824 juvenis de M. exigua em 5 g de raízes em clone não identificado, em levantamento realizado no município de São José do Rio Claro, MT. Entre maio e novembro houve uma queda de 91,41 % na média de infestação, sendo que a menor população foi em novembro, onde a explicação mais aceitável deve-se a hibernação da seringueira, quando esta cessa ou reduz seu metabolismo radicular. Soong (1976), em experimento na Malásia, demonstrou que o desenvolvimento radicular variou sazonalmente nas diferentes profundidades, sendo que nas camadas superficiais, o maior crescimento foi no período de reenfolha natural das plantas, posterior a hibernação (fevereiro e março) e o menor desenvolvimento ocorreu antes da desfolha natural anual (agosto a dezembro). Para as condições do presente experimento, pode ter ocorrido situação semelhante, somente mudando os meses de hibernação (maio a julho), o que pode ter favorecido a redução da infestação devido ao menor sistema radicular.
No período de hibernação da seringueira, ocorre a morte de parte das raízes das plantas, sendo acompanhada pela redução da população de M. exigua raça 3. Após o reenfolhamento das plantas, as raízes que permaneceram vivas, infestadas de nematoides e M. exigua em estágio latente no solo são fonte de inóculo para novas infestações. Juntamente a novos juvenis que se desenvolveram a partir de ovos, estimulados pela presença de condições favoráveis (umidade e exsudatos radiculares), iniciam infestações nas raízes que crescem depois da hibernação. Desta forma, a população de nematoides volta a aumentar. O nível de infestação predominante em maio foi dentro das classes II e III. Já para novembro, ocorreu somente a classe I de população (figura 4). A distância entre os pontos amostrais foi de 34,3 a 15.576,78 metros e área média de 25,17 ha por ponto amostral em maio e de 24,69 ha em novembro.
Como esse nematoide é nativo da região de Itiquira (Santos, 1995), certamente, antes do plantio a área apresentava razoável população, porém, com o passar dos anos, a infestação ficou próxima da encontrada no ano de 2013, dentre os talhões do mesmo clone. Este fato ocorreu devido aos tratos culturais realizados para o manejo do plantio, onde ocorreu o permanente trânsito de maquinários na área plantada, disseminando M. exigua raça 3 em locais onde estavam ausentes. Conforme o histórico desta área, o uso de grades para a limpeza das entre linhas era uma atividade rotineira.
Para o clone RRIM 600, as infestações máximas foram de 197.918 nematoides e 27.690 nematoides, em maio e em novembro, respectivamente, demonstrando queda acentuada, da ordem de 86 %, período que de acordo com a média pluviométrica, apresenta menor e maior precipitação, respectivamente. No entanto, em novembro, ocorreu enquadramento da infestação pontual com 27.690 nematoides/ 10 g de raízes para a classe I de infestação, devido à suavização dos valores utilizados no interpolador da krigagem (figura 5). Estes dados concordam com Souza, Souza, Santos e Silva (1998), que constataram que M. exigua no cafeeiro (provavelmente raças 1 e 2) apresentou maiores infestações e número de galhas nas raízes no período chuvoso e no solo ocorreu queda na população de juvenis.
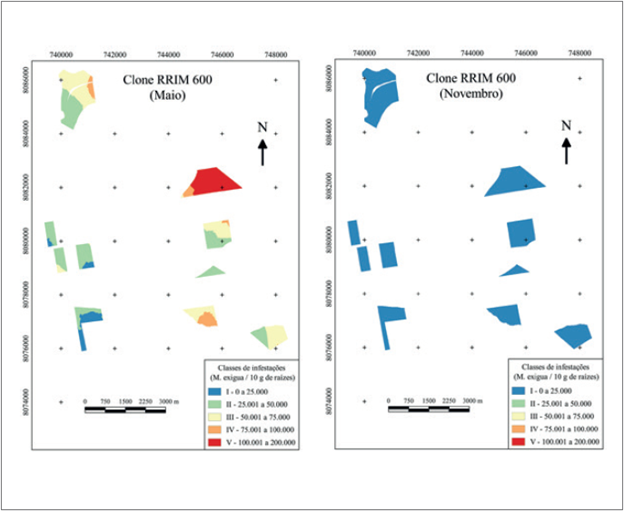
Para as médias de infestações, os valores encontrados em maio e novembro foram de 46.284 e 6.170 nematoides, respectivamente, havendo redução de 86,67 % na infestação. A suscetibilidade do clone RRIM 600 ao nematoide foi demonstrada por Fonseca, Ferraz e Machado (2003), que em casa de vegetação inocularam 1.000 juvenis J2 em mudas e, após 10 dias, verificaram presença de células gigantes, sendo as responsáveis pela nutrição do nematoide.
A mancha com maior população do nematoide ocorreu nas coordenadas UTM 21S 8082000 L/ 744000 N, intervalo de 100.001 a 200.000 nematoides, predominando em mais de 90 % do talhão. A infestação com maior área foi a classe II, sendo que foi ausente somente em dois talhões dentre os dez talhões estudados. No plantio total, em novembro, as infestações enquadraram-se somente na classe I (cor azul no mapa), sendo que a maior infestação pontual foi de 50.100 nematoides (figura 5), mas devido a suavização ocasionada pela krigagem, este ponto amostral encaixou-se na referida classe.
Silveira (1992) citou que o clone RRIM 600 é altamente suscetível aos nematoides das galhas. Santos et al. (1992) e Fonseca, Jaehn e Silva (1999) citaram que este clone é suscetível a M. exigua raça 3 e resistente a Meloidogyne javanica (Treub) Chitwood. Esta suscetibilidade foi reafirmada com a análise no presente experimento, com 197.918 nematoides/ 10 g de raízes.
As infestações em maio e novembro demonstraram suscetibilidade a M. exigua raça 3, fato destacado por Fonseca et al. (1999) no clone RRIM 600. As plantas dos clones GT 1 e RRIM 600 avaliadas no plantio em Itiquira, exibiam severos sintomas da doença, com grande quantidade de galhas nas raízes (figura 6), variando a intensidade da sua presença entre os clones, mas com presença em todos, demonstrando desta forma que o patógeno apresenta alta agressividade, dificultando o controle deste fitoparasita.

Fonte: Elaboração própria
Figura 6 Raízes de seringueira com presença de galhas, sintomas do Meloidogyne exigua raça 3.
A ocorrência da maior quantidade de nematoides foi constatada em maio, pois este mês ocorre posteriormente a época de altas temperaturas e período chuvoso, o que é favorável à reprodução do nematoide. Com isso, a população fica alta, devido à ausência de inimigos naturais e de controle químico, onde a monocultura de seringueira selecionou poucos nematoides daqueles que estavam no solo, dentre eles M. exigua raça 3, cuja população passou a crescer.
A ausência de identificação do porta-enxerto tem grande influência na avaliação da infestação. Porém, este nematoide é altamente agressivo à seringueira, conforme avaliaram Paes-Takahashi et al. (2015), relatando intolerância e susceptibilidade dos porta-enxertos GT 1, PB 235, PB 217, RRIM 501, PR 255, IAN 873, RRIM 600 e TJ 1 a M. exigua e Pratylenchus brachyurus Filipjev & S. Stekhoven.
Supondo-se que no plantio da presente avaliação foi utilizado somente um clone como porta-enxerto, a alta variabilidade de infestações dentre os diferentes clones pode ser devido a troca de diferentes metabolitos entre a interação com o enxerto; a população anteriormente estabelecida na área, as quais parasitavam plantas nativas; diferenças de umidades do solo dentre os talhões, dentre outros fatores.
Conclusões
As infestações de M. exigua raça 3 no mês de maio, nos clones GT 1 e RRIM 600, são maiores quando comparadas com as infestações no mês de novembro;
As variações espaciais das infestações dentro do mesmo clone são altas, apresentando IDE em maio e novembro considerados moderados;
A aleatoriedade da infestação foi alta em ambos os meses e clones avaliados, devido à alta amplitude populacional presente nestes clones, refletindo na validação cruzada dos dados;
Em maio, os clones RRIM 600 e GT 1 demonstraram 197.918 e 159.840 nematoides / 10 g de raiz, respectivamente, onde ambos os clones de copa apresentaram infestações altas.
REFERÊNCIAS
Bernardo, E. R. A., Santos, J. M., Silva, R. A., Cassetari-Neto, D., Santos, S. S., Delmadi, L., & Rocha, V. F. (2003). Levantamento de Meloidogyne exigua na cultura da seringueira em São José do Rio Claro, MT. Ciência Rural, 33(1), 157-159. [ Links ]
Cambardella, C. A., Moorman, T. B., Novak, J. M., Parkin, T. B., Karlen, D. L., Turco, R. F., & Konopka, A. E. (1994). Field-scale variability of soil properties in central Iowa soils. Soil Science Society of America Journal, 58(5), 1501-1511. [ Links ]
Campelo-Júnior, J. H. (2000). Estimativa da transpiração em seringueira. Revista Brasileira de Agrometeorologia, 8(1), 35-42. [ Links ]
Cardinal, A. B. B., Gonçalves, P. S., & Martins, A. L. M. (2007a). Influência de seis portaenxertos sobre a produção de clones superiores de seringueira. Bragantia, 66(2), 277-284. [ Links ]
Cardinal, A. B. B., Gonçalves, P. S., & Martins, A. L. M. (2007b). Stockscion interactions on growth and rubber yield of Hevea brasiliensis. Scientia Agricola, 64(3), 235-240. [ Links ]
Castrignanò, A. (2011). Introduction to spatial data processing. Rome, Italy: Aracne. [ Links ]
Coolen, W. A., & D’Herde, C. J. (1972). A method for the quantitative extraction of nematodes from plant tissue. Merelbeke, Belgium: State Nematology and Entomology Research Station. [ Links ]
Empresa Brasileira de Pesquisa Agropecuária (Embrapa). (2013). Sistema brasileiro de classificação de solos (3ª ed.). Brasilia, Brazil: Embrapa. [ Links ]
Farias, P. R., Barbosa, J. C., Vieira, S. R., Sánchez-Vila, X., & Ferraz, L. C. C. B. (2002). Geostatistical analysis of the spatial distribution of Rotylenchus reniformis on cotton cultivated under crop rotation. Russian Journal of Nematology, 10(1), 1-10. [ Links ]
Ferris, H., & Wilson, L. T. (1987). Concepts and principles of population dynamics. In J. A. Veech & D. W. Dickson (Eds.), Vistas on nematology: a commemoration of the twenty-fifth anniversary of the Society of Nematologists (pp 372-376). Hyattsville, USA: Society of Nematologists. [ Links ]
Fonseca, H. S., Jaehn, A., & Silva, M. F. A. (1999). Reações de porta-enxertos de seringueira (Hevea brasiliensis) a Meloidogyne javanica e M. exigua. Nematologia Brasileira, 23(2), 9-14. [ Links ]
Fonseca, H. S., Ferraz, L. C. C. B., & Machado, S. R. (2003). Ultra-estrutura comparada de raízes de seringueira parasitadas por Meloidogyne exigua e M. javanica. Nematologia Brasileira, 27(2), 199-206. [ Links ]
Furtado, E. L. (2010). Doenças da seringueira e seu controle. II Encontro Brasileiro de Heveicultura [CD-ROM]. Ilhéus, Brasil. [ Links ]
Isaaks, E. H., & Srivastava, R. M. (1989). An Introduction to Applied Geostatistics. New York, USA: Oxford University Press. [ Links ]
Journel, A. G., & Huijbregts, C. J. (1978). Mining geostatistics. London, United Kingdom: Academic Press. [ Links ]
Köppen, W. (1923). Die klimate der Erde. Berlin, Germany: W. Guyter. [ Links ]
Landim, P. M. B. (2006). Sobre geoestatística e mapas. Terrae Didatica, 2(1), 19-33. [ Links ]
Landim, P. M. B. (2003). Análise estatística de dados geológicos. São Paulo, Brazil: Universidade Estadual Paulista (Unesp). [ Links ]
Landim, P. M. B., & Sturaro, J. R. (2002). Krigagem indicativa aplicada à elaboração de mapas probabilísticos de riscos. Rio Claro, Brazil: Universidade Estadual Paulista (Unesp). [ Links ]
Moraes, V. H. F. (1982). Sugestões para uniformização da metodologia de estudo da fenologia foliar da seringueira. Manaus, Brazil: Empresa Brasileira de Pesquisa Agropecuária (Embrapa). [ Links ]
Muniz, M. F. S., Campos, V. P., Castagnone-Sereno, P., Castro, J. M. C., Almeida, M. R. A., & Carneiro, R. M. D. G. (2008). Diversity of Meloidogyne exigua (Tylenchida: Meloidogynidae) populations from coffee and rubber tree. Nematology, 10(6), 897-910. [ Links ]
Paz, A., Taboada, M. T., & Gómez, M. J. (1996). Spatial variability in topsoil micronutrient contents in one-hectare cropland plot. Communications in Soil Science and Plant Analysis, 27(3/4), 479-503. [ Links ]
Paes-Takahashi, V. S., Soares, P. L. M., Guiducci, E. P., Brito, P. F., Carneiro, F. A., & Ferreira-Junior, R. (2015). Reação de porta-enxertos de seringueira a Meloidogyne exigua e a Pratylenchus brachyurus. Nematropica, 45(2), 242-251. [ Links ]
Rajagopal, R., Vijayakumar, K. R., Thomas, K. U., & Karunaichamy, K. (2004). Effect of judicious ethephon application on yield response of Hevea brasiliensis (Clone RRII 105) under ½S d/3 6d/7 tapping system. Journal of Rubber Research, 7(2), 138-147. [ Links ]
Randig, O., Bongiovanni, M., Carneiro, R. M., Sarah, J. L., & Castagnone-Sereno, P. (2002). A species-specific satellite DNA family in the genome of the coffee root-knot nematode Meloidogyne exigua: application to molecular diagnostics of the parasite. Molecular Plant Pathology, 3(6), 431-437. [ Links ]
Santos, J. M. (1995). Meloidogyne exigua e Botryodiplodia theobromae, principais componentes bióticos de uma doença complexa da seringueira em Mato Grosso. Fitopatologia Brasileira, 28, 341. [ Links ]
Santos, J. M., Matos, C., Barré, L., & Ferraz, S. (1992). Meloidogyne exigua, sério patógeno da seringueira nas plantações Michelin, em Rondonópolis, MT. Em Anais do Congresso Brasileiro de Nematologia. Lavras, Brazil: Sociedade Brasileira de Fitopatologia. [ Links ]
Silveira, S. G. P. (1992). Nematoides associados à cultura da seringueira (Hevea spp.). In M. J. S. Medrado, M. S. Bernardes, J. D. Costa, & A. N. Martins (Eds.), Formação de mudas e plantio de seringueira (pp. 65-77). Piracicaba, Brasil: Escola Superior de Agricultura Luiz de Queiroz (Esalq). [ Links ]
Soong, N. K. (1976). Feeder root development of Hevea brasiliensis in relation to clones and environment. Journal of the Rubber Research Institute of Malaysia, 24, 283-298. [ Links ]
Souza, S. E., Souza, L. H., Santos, F. S., & Silva, R. V. (1998). Flutuação populacional de Meloidogyne exigua (Goeldi, 1887) em cafeeiros no município da Barra do Choça, BA. Bahia Agrícola, 2(3), 77-80. [ Links ]
Thompson, S. K. (1992). Sampling. New York, USA: John Wiley & Sons. [ Links ]
Trangmar, B. B., Yost, R. S., Wade, M. K., Uehara, G., & Sudjadi, M. (1987). Spatial variation of soil properties and rice yield on recently cleared land. Soil Science Society of America Journal, 51(3), 668-674. [ Links ]
Vieira, S. R., Hatfield, J. L., Nielsen, D. R., & Biggar, J. W. (1983). Geostatistical theory and application to variability of some agronomical properties. Hilgardia, 51(3), 1-75. [ Links ]
Vieira, S. R. (2000). Geoestatística em estudos de variabilidade espacial do solo. In R. F. Novais, V. H. Alvarez & C. E. G. R. Schaefer (Eds.), Tópicos em ciência do solo (pp. 1-54). Viçosa, Brazil: Sociedade Brasileira de Ciência do Solo. [ Links ]
Wallace, M. K., & Hawkins, D. M. (1994). Applications of geostatistics in plant nematology. Journal of Nematology, 26(4S), 626-634. [ Links ]
Webster, R., & Boag, B. (1992). Geostatistical analysis of cyst nematodes in soil. European Journal of Soil Science, 43(3), 583-595. [ Links ]
Yeates, G. W. (1987). How plants affect nematodes. Advances in Ecological Research, 17, 61-113. [ Links ]
Zimback, C. R. L. (2001). Análise espacial de atributos químicos de solos para fins de mapeamento da fertilidade do solo (Tese de docência). Universidade Estadual Paulista, Botucatu, Brasil. [ Links ]
Recebido: 24 de Janeiro de 2017; Aceito: 03 de Abril de 2018
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
