











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introduction
Naidí or açai, Euterpe oleracea Mart. (Arecaceae), is a palm that grows in South America (Jones, 1995). This palm is widely distributed in Brazil (Gallori et al., 2004), Ecuador, the Guianas, Panama, Venezuela, and Colombia (Galeano & Bernal, 2018). It is found in higher numbers in periodically flooded soils, although it also grows in lower densities in non-flooded soils (Cavalcante, 1974). In Colombia, E. oleracea has grown in the Amazon and Pacific regions, as well as in the Magdalena valley (Galeano & Bernal, 2018). The naidí palm fruit is a purple-black berry (naidí) (Gallori et al., 2004). It is massively consumed due to its high nutritional value and elevated levels of minerals, lipids, and proteins (Silva et al., 2014), representing a source of economic income to the families that sell it in local markets (Montenegro-Gómez & Rosales-Escarria, 2015). Corpocampo, a business organization based on sustainable use strategies, has approximately 5,000 hectares planted between Buenaventura (Valle del Cauca) and Putumayo and three production plants in Guapi (Cauca), Tumaco (Nariño), and Puerto Asís (Putumayo), where it processes five million tons of naidí per year, of which 98 % is exported and only 2 % is for national sale (Llano, 2017).
Arbuscular mycorrhizal fungi (AMF) of the Glomeromycota phylum, are known for their beneficial associations with plants and have been reported to colonize more than 80 % of plant roots (Ambrosino et al., 2018; Rivera et al., 2016). However, little information about AMF related to tropical palms has been published in the literature (Molineros Hurtado et al., 2013). Nonetheless, the result found suggests that palms are dependent on AMF colonization, as it has been described for Euterpe edulis (Bonfim et al., 2015; Sgrott et al., 2012), Euterpe precatoria (Garcés-Ruíz et al., 2017), and Bactris gasipaes (Garcés-Ruiz et al., 2017; Riascos-Ortiz et al., 2011). AMF have been observed developing associations with E. oleracea as reported by Chu (1999), who found benefits in growth and nutrient acquisition in pre-nursery and nursery stages. The establishment of E. oleracea seeds is a complex process as plants show slow growth and high mortality (Bovi et al., 1987). However, a significant increase in growth and nutrient acquisition in seedlings was observed after being inoculated with seven AMF species, demonstrating that E. oleracea is a mycorrhizal-dependent plant (Chu, 1999).
When this research was conducted, no literature information about AMF related to the rhizosphere of E. oleracea, and if the soil chemical properties –mainly phosphorous– can affect the natural populations of AMF to obtain the maximum benefit of symbiosis with the naidí palm, was found for Colombia. Accordingly, the aim of this research was to contribute with information related to some environmental factors that may affect the occurrence and distribution of native AMF so that it can be used in other locations.
Materials and methods
Study area
The research was conducted in two areas located in Bajo Calima, Buenaventura, Colombia, with coordinates 03°54' and 04°00' North latitude and 77°05' and 77°30' West longitude. The climate of the region is classified as Af (tropical rain forest) by Köppen, with a mean annual rainfall of 7,328 mm, and a mean annual temperature is 26.1 °C. The occurrence of regular floods characterizes the region. Soil samples were collected from two different sites with the presence of E. oleracea in an area under the influence of rice cropping (L1), and the other two sites were located in a rainforest area (L2). Three soil samples were extracted from the rhizosphere of E. oleracea from 0 to 10 cm of depth. Fine roots in E. oleracea (< 1 mm) are more abundant in this soil layer compared to other depths. The three soil samples were mixed into one composite sample until completing 1,000 g. The soil and root samples were placed in plastic bags and stored at 4 °C (before preparation and analysis), and transported to the laboratory.
Soil chemical analyses
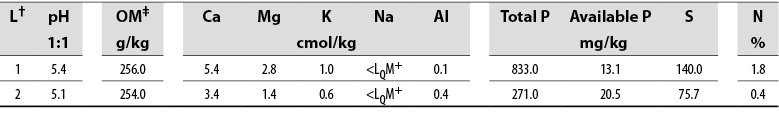
Soil chemical properties were determined in 150 g of all soil the samples. pH was measured in a 1:1 soil: water ratio using a pH meter (Lab 855 - SI Analytics, GmbH, Mainz, Germany), and organic matter content was determined by the Wakley Black K2Cr2O7-H2SO4 colorimetric method. Available and total phosphorous were established following the Bray II and acid digestion methods, respectively. Aluminium was extracted with KCl; calcium, magnesium, potassium, and sodium were determined by atomic absorption spectrometry. Soil samples were analyzed at the International Center of Tropical Agriculture (CIAT).
Root and spore sampling
AMF spores were extracted from 100 g soil subsamples by the wet sieving and decanting method (Brundrett & Juniper,1995). Spores were counted and identified by mounting on glass slides with PVLG mixed with Melzer’s reagent (1:1 v/v) (Morton, 1991). The spore number is given in AMF/100 g of dry soil (Sánchez De Práger et al., 2010; Sieverding, 1983). For estimating the colonization rate of AMF, fine roots in E. oleracea (< 1 mm) were sampled from the soil mixtures, cleared in KOH 10 %, and stained in Trypan Blue (0.05 %). Stained roots were washed with water, and dye excesses were removed with lactoglycerol; root fragments of 1 cm long were cut and mounted in parallel on plates according to Deguchi et al. (2017), Rivera et al. (2016), and Sieverding (1983).
Statistical analysis
Mycorrhizal colonization and spore number between two areas were compared using the t-test in the SigmaPlot software version 11.0 (SYSTAT Software Inc., California, USA). Available P level, spore abundance, and mycorrhizal colonization were analyzed through Pearson’s product-moment correlation in SigmaPlot. Spore abundance and mycorrhizal colonization were transformed using log10 in Pearson’s product-moment correlation.
Results and discussion
Segundo título Soil chemical analyses
Soil pH was low in both evaluated areas, ranging from 5.12 to 5.44. On the other hand, high levels of total and available P were observed in L1 and L2, respectively. Similar organic matter content was observed in L1 and L2 (table 1). The detected level of Ca, Mg, K, P, S, and N in the soil varied between L1 and L2.
Mycorrhizal colonization and spore abundance
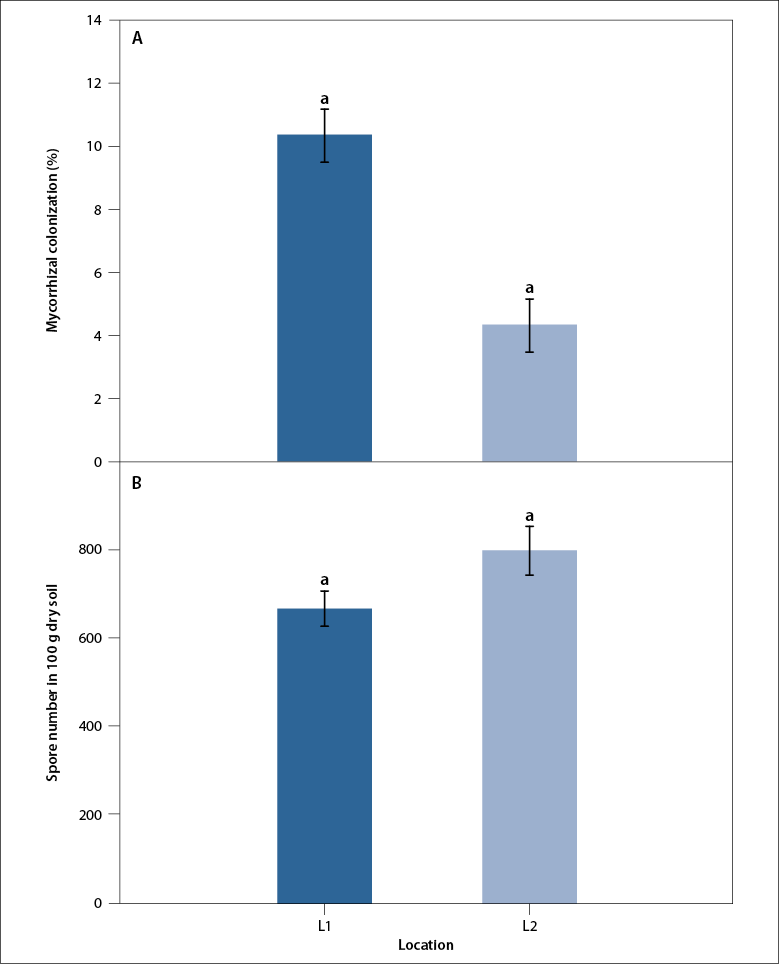
Mycorrhizal colonization in L1 (10.2 ± 0.9 %) was significantly higher than L2 (4.3 ± 0.9 %). According to Pearson’s product-moment correlation, a negative correlation between available P levels and mycorrhizal colonization in L1 (p = 0.006; r = -1.0) and L2 (p = 0.02; r = -1.0) was observed. Spore number ranged from 660 to 791 spores per 100 g of dry soil (figure 1) although L2 showed the highest spore abundance.
Source: Elaborated by the authors
Figure 1 A. Mycorrhizal colonization (%); B. Spore abundance (spore number/100 g of dry soil) associated with the rhizosphere of E. oleracea in an area under the influence of rice cropping (L1) and in a rainforest area (L2) in the Colombian Pacific region. Bars represent mean values and standard error (SE). Different letters show a significant difference between the areas sampled (p < 0.05).
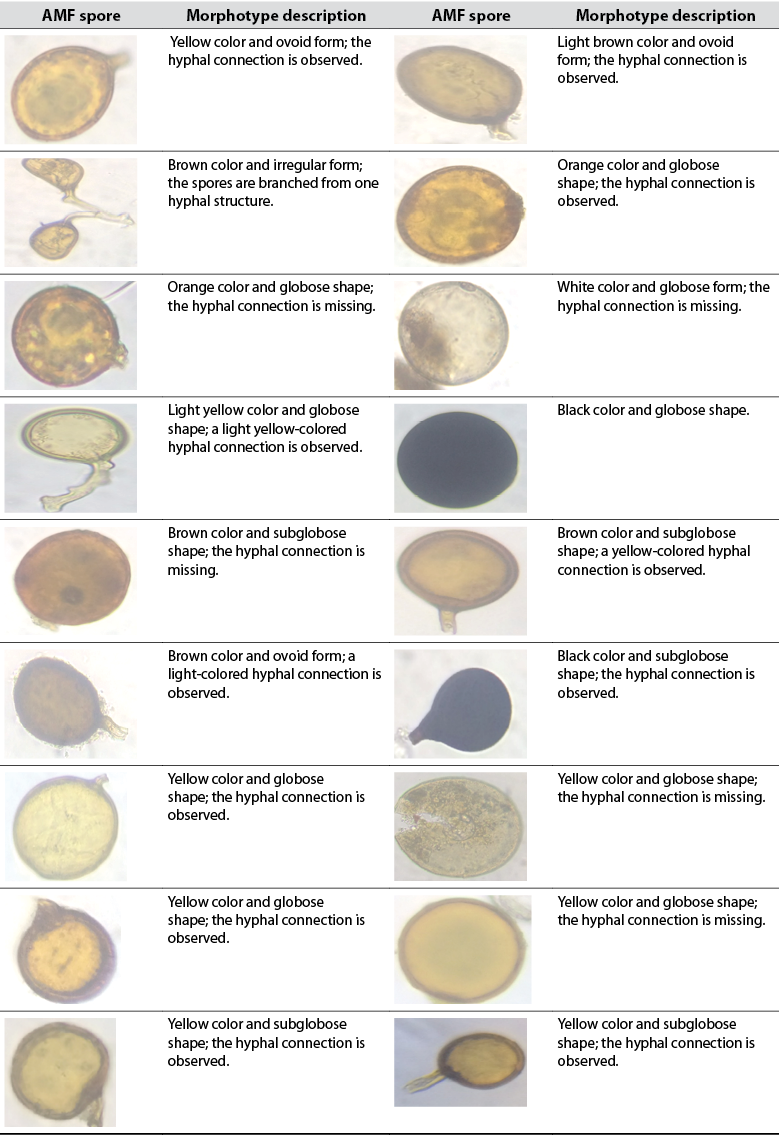
Ovoid and globose spores represented in several morphotypes were recorded (tables 2 and 3). Spores observed in table 2 belong to L1, while spores shown in table 3 were found in L2. Based on the images displayed in tables 2 and 3, the bulk of spores are glomoid (Glomus morphotype) and can be classified as Claroideoglomeraceae, Glomeraceae, Diversisporaceae, or Paraglomeraceae; there are also dimorphic species that can be classified as Ambisporaceae. Within Glomeromycota, species with glomoid spore formation represent the largest group (Oehl et al., 2011). Black colored spores represent the same morphotype and possibly belong to the same species.
Table 2 Arbuscular mycorrhizal fungi (AMF) spores isolated from the rhizosphere of E. oleracea in an area under the influence of rice cropping (L1)
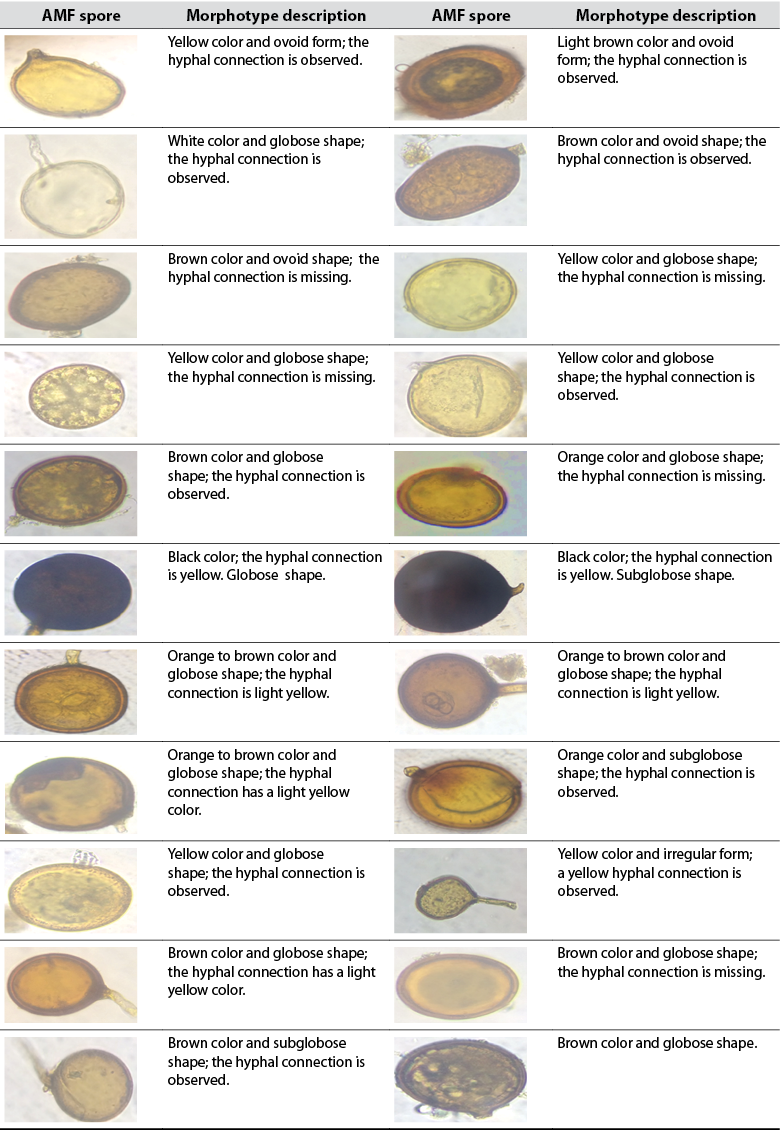
Source: Elaborated by the authors. Photographs taken by Maité Rosales
Discussion
In this study, the mycorrhizal colonization and spore abundance associated with the rhizosphere of E. oleracea was evaluated in two areas, the first one under the influence of a flooded rice field, and the second in a rainforest zone, both located in Bajo Calima, Buenaventura, in the Colombian Pacific region, to understand how the level of available P affects the mycorrhizal colonization and spore abundance. In the current study, soil analysis showed high levels of total P, mainly in L1, where large amounts of fertilizers are applied in the rice fields located near the area where E. oleracea plants are found. According to Casas (2012), available P showed adequate and high levels in the L1 and L2 areas, respectively (table 1).
However, several studies indicate that soils rich in natural forms of P or high P levels due to fertilization decrease root colonization by AMF (Maia et al., 2009; Moreira & Siqueira, 2006). This situation was observed in our study, where levels of available P were 13.10 mg/kg in L1 and 20.50 mg/kg in L2, while mycorrhizal colonization was low (L1: 10 % and L2: 4 %). This agrees with the findings on mycorrhizal dependence dynamics reported by Ramos-Zapata et al. (2006) in Desmoncus orthacanthos, a native palm from Yucatan (Mexico), where lower available P levels in the soil (12 ppm) showed higher mycorrhizal colonization (25 %), while higher available P levels (24 ppm) recorded lower values of mycorrhizal colonization (7 %). Therefore, the E. oleracea relation observed between available P and mycorrhizal colonization would indicate that the symbiotic association could be related to the absorption of this element.
Nevertheless, other factors could be associated with low colonization rates since there are not enough studies that relate ecological factors to AMF dynamics in tropical rainforests (Eom et al., 2000). For instance, correlations between AMF abundance or mycorrhizal colonization and pH values or organic matter content were reported in the palm E. edulis in the Brazilian Atlantic rainforest (Bonfim et al., 2015). However, no correlation with pH or organic matter content was evaluated in our data. In spite of having carried out the study in tropical rainforest soils characterized by extremely low pH values and fertility (Martins et al., 2015; Pereira et al., 2010), AMF may be essential in the establishment and survival of forest plants due to their crucial roles in soils (Zangaro & Moreira, 2010).
On the other hand, although individual soil characteristics have been reported to play major roles in AMF communities in natural and agricultural ecosystems (Jansa et al., 2014), the driving effect of soil properties could not be attributed to a single soil characteristic, but to the interactions among all of them along with host plant and environmental factors not related to soil properties (Alguacil et al., 2016). Although, the role of pH in mycorrhizal colonization is not well known, it is important, considering the acid condition in the soils evaluated in this work (Clark, 1997; Coughlan et al., 2000; Duarte et al., 2018; Siqueira et al., 1984). Conversely, the low mycorrhizal colonization (< 20 %) observed in this study could be associated with the effect of soil flooding, as it was reported in studies carried out under the same conditions (Miller, 2000; Pérez et al., 2011).
According to Zangaro et al. (2008, 2012), the maturity of E. oleracea plants growing in secondary forests on a late succession (Mazuera & Ladrach, 1981) is also a factor that can decrease the mycorrhizal colonization in mature forests. In this type of forest, low mycorrhizal colonization values have been recorded, indicating that trees with lower metabolic activity could not withstand the symbiosis association due to decreasing carbohydrates availability (Zangaro et al., 2008; 2012). Furthermore, comparing several vegetal covers (soil covered with native grasses, and secondary and mature forests) at depths of 0 - 10 cm and 10 - 20 cm, a negative relation among soil depth and vegetation maturity with mycorrhizal colonization was observed. Moreover, the lowest values were found in the mature forest and in soil layers deeper than 10 cm (Zangaro et al., 2014); this has also been reported by other authors (Brown & Bledsoe, 1996; Cardoso et al., 2003; Ingleby et al., 1997; Muthukumar et al., 2003; Powers et al., 2005).
Considering the relevance of this palm in Colombia, as well as the environmental factors affecting the symbiotic association in the Pacific region, we compared our spore abundance results with the one found in E. edulis, a palm growing in Brazilian tropical rainforests. In our study, we recorded higher spore abundance compared to those stated in similar rainforest areas in Brazil. Medina et al. (2012) reported a low spore abundance (120 - 400 spores in 100 g of dry soil) in five forest areas in Minas Gerais, while Bonfim et al. (2015) found between 8 and 22 spores in 1 g of dry soil in the Atlantic rainforest in Sao Paulo. Still, available P levels could also affect spore abundance in our study, considering that high levels of P also affect AMF sporulation (Lin et al., 2012; Maia et al., 2009).
Several AMF morphotypes associated with E. oleracea that can be classified as Claroideoglomeraceae, Glomeraceae, Diversisporaceae, Paraglomeraceae, and Ambisporaceae were found. Fifty-eight AMF taxa associated with the rhizosphere of E. edulis were classified using molecular techniques, into Ambisporaceae, Diversisporaceae, Acaulosporaceae, Entrophosporaceae, Glomeraceae, Dentiscutataceae, Gigasporaceae, Intraornatosporaceae, Racocetraceae, Scutellosporaceae, and Praglomeraceae along an altitudinal gradient in the Brazilian Atlantic forest. Further, this region was considered as an AMF biodiversity hotspot (Bonfim et al., 2015). In this sense, the region where this study was conducted shows interesting results on AMF diversity due to the similar climatic conditions to the Atlantic rainforest. Similarly, using molecular tools, genera assigned to Acaulosporaceae, Archaeosporaceae, and Glomeraceae were reported as associated with E. precatoria in polluted soils in the Amazon forest in Ecuador (Garcés-Ruiz et al., 2017). However, it is important to consider that samples collected in the field are exceedingly variable since spores are highly exposed to parasitism and deterioration by several edaphic agents (Leal et al., 2009).
This article is the first report on AMF associated with the rhizosphere of E. oleracea and to the Euterpe genus in Colombia. Nonetheless, AMF association in the rhizosphere of E. oleracea under greenhouse conditions has already been revealed (Chu, 1999). In any case, further studies with robust identification are needed to evaluate the AMF structure, composition, and diversity related to E. oleracea in the Colombian Pacific region, considering the latter as a source of native AMF species, which would be a good alternative for agricultural and agroforestry systems with limited nutrient conditions.
Conclusions
In this study, low mycorrhizal colonization and high spore abundance were found in two areas with E. oleracea palms. The spore abundance found in the rhizosphere of E. oleracea suggests a potential to explore native AMF in the Colombian Pacific region, which can be articulated with a sustainable palm production. An available P level in the soil negatively affected AMF root colonization, but the influence of other edaphic parameters along with the plant effect could also be related to ecological factors on the AMF dynamics in tropical forests. Studies about E. oleracea in Colombia are required since it is vital to carry out bioprospecting works, including AMF, in the sustainable production of E. oleracea palms under different environmental conditions.

