











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
Introduction
The Colombian bovine herd’s reproductive efficiency (RE) does not exceed 53% (Santana et al., 2009). This low RE is due to various factors that affect fertility and result in a wide calving interval (CI), such as nutrition, imposed environmental management (Granja et al., 2012; Ossa et al., 2007; Vergara et al., 2008), and the use of breeds and crosses not adapted to the environmental supply (Galeano & Manrique, 2010; Perdomo et al., 2017). Even though Colombia has seven native breeds, including Romosinuano, which have shown higher RE than zebu (De Alba, 2011), the reproductive behavior exhibited by the former at the age of the first calving (1,133 ± 200 days) and CI (422 ± 131 days) (Ossa et al., 2013) stand out.
The CI is a determining component of RE that expresses the female’s ability to produce one calf and one lactation per year; it is also related to the number of successive births that the animal is capable of giving during its reproductive life (López et al., 2006). The RE of a female in particular and the population to which she belongs in general is the sum of fertility and fecundity (Ossa, 2003) and can be evaluated by the number of services per conception, age at the first calving, length of calving-mating intervals and CIs. The calving-mating interval directly affects the calving period length and, consequently, the CI (Ossa et al., 2007; Ríos-Utrera et al., 2013; Vergara et al., 2008). The CI is influenced by animal genotype, month and year of delivery, the mother’s age (Ossa, 2017a), and year-season of the year or sex of the calf-year interactions (Cardellino & Brazil, 1987). Wilcox (1957) proposed an equation based on the CI to calculate RE.
RE is a trait of economic importance in bovine production systems because a change in behavior results in a variation in the production system profitability (Casas & Tewolde, 2001; Vergara et al., 2008); therefore, increasing the RE is one of the most critical livestock breeding challenges (Galvão et al., 2013). The CI is the trait that has the most significant influence on reproductive behavior and has a different intensity of expression (Cardellino & Brasil, 1987). Additionally, since parturitions occur several times during the bovine female’s reproductive life, and the date each time a parturition occurs is recorded, it is valid to say that the trait is repeatable.
The variance of a trait at different stages of the animal’s life can be analyzed under two components: variance within individuals, which measures temporary differences in the same individual’s performance, and variance between individuals, which is partially genetic and environmental, since the environmental portion is caused by environmental circumstances that permanently affect the individual’s performance (Ossa, 2003).
The repeatability of a particular trait measures the correlation between the repeatable measurements of the same trait in the same individual, considering the constant environmental conditions over time (Rúales et al., 2007, as cited in Galeano & Manrique, 2010). It has a series of considerations, namely, it marks the limit of heritability, which can be equal to or less than repeatability, but never greater; when calculated for the average of a high number of production records, it offers greater assurance than when calculated for few records; it is used to estimate the most probable producing ability (MPPA) of the animal, indicating the animal’s production in the next parturition (Ossa, 2003). The repeatability values of the CI, reported in the literature by Cardellino and Brasil (1987), Galeano and Manrique (2010), and Yagüe et al. (2009), are low and varied between 0.05-0.10.
Repeatability, heritability, and genetic correlations of populations are necessary to estimate individuals’ genetic value (GV) (Quijano et al., 2011) and establish genetic improvement programs (Galeano & Manrique, 2010). Due to the low RE (Vergara et al., 2008), which in turn decreases livestock profitability (Martínez-Rocha et al., 2012), and the few studies on beef cattle related to the genetic evaluation of reproductive parameters (Montes et al., 2009), this paper aims to estimate CI repeatability and establish its relationship with RE in Romosinuano females of the Nation’s Germplasm Bank for Food and Agriculture (SBGNAA, for its acronym in Spanish), AGROSAVIA Turipaná Research Center, using records for 81 years (1936-2016).
Materials and methods
The information used in this study was obtained from the SBGNAA reproductive records of cows of the Colombian native breed Romosinuano, kept by AGROSAVIA in the Turipaná research center located in the Valle del Sinú, northeastern Colombia.
The database includes the informative record of the CI trait for 1936-2016. For this information to be truthful and consistent, we eliminated the days between parturitions that were outside the range considered normal (300-1,278 days) using the dplyr package (Wickham et al., 2019) of R 3.6.0 (R Core Team, 2019), specialized in statistical data analysis. After editing the information, 86.9% of the total number of records was useful.
Four classes were formed with the calving month variable. Similarly, the bovine female’s calving year variable was grouped into ten-year periods, comprising eight classes, to have a better representation of the CI variable in each calving month and year class since the information was scarce when considering each variable separately. Except for the first to sixth parturitions, it was necessary to group births seven and eight in the same class (Calving 7) and births equal to or greater than nine in a class called Calving 8. The variables considered in the study are described in table 1.
Table 1 Description of analyzed variable CI, calving month class, calving year class, and calving number
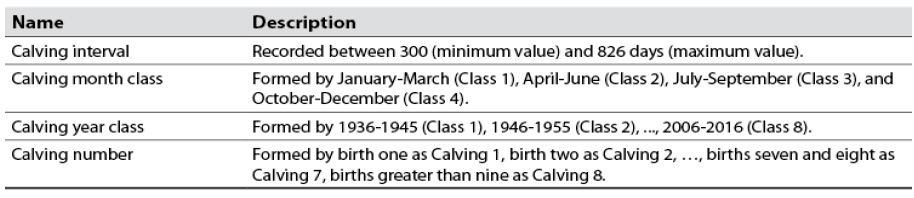
Source: Elaborated by the authors
The model used to calculate the variance components, necessary to estimate the repeatability value of the CI characteristic, was an animal model of repeated measures over time, whose scalar form can be represented as follows:
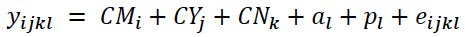 Equation 1
Equation 1
Where yijkl is the Bovine Female l’s CI, considering the fixed effects of calving month class (CM), calving year class (CY), and calving number (CN). The random-effects considered include the Bovine Female l’s additive GV (al) for the CI, the Bovine Female l’s permanent environment effect (pl) on CI in its various births, and the residual effect (eijkl). This model assumes that the effect of genes on the same bovine female’s trait does not change with calving number and that the permanent environment affects each measure taken in the same animal equally.
The matrix form of the repeatability model can be represented as follows:
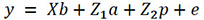 Equation 2
Equation 2
Where y: CI vector, b: fixed effects solution vector; a: unknown additive genetic random effects vector; p: unknown permanent environmental effects vector; X, Z1, and Z2: the unknown incidence matrices that relate the records to fixed (b), additive genetic (a), and permanent environment (p) effects, respectively; e: the residual effects vector.
The expected values (E) and variances (v) for the model’s random effects are as follows:
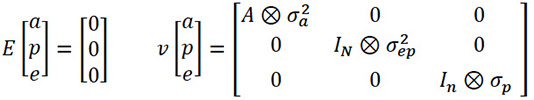 Equation 3
Equation 3
Where A: kinship matrix among bovines in the pedigree; ⊗: Kronecker product between matrices; 𝜎2a: additive genetic variance; IN: identity matrix whose size is equal to the number of cows; 𝜎2ep: permanent environment variance; In: identity matrix whose size is equal to the number of observations; 𝜎2a: residual variance.
The repeatability value of the CI trait was estimated using the rptR package (Stoffel et al., 2019) of R, specialized in statistical data analysis. In addition to estimating this value, the package can quantify the uncertainty in the estimated repeatability values (Stoffel et al., 2017) through the bootstrap method.
Each bovine’s GV was calculated based on Equation 4 using the variance components estimated by this statistical package (Quijano & Echeverri, 2015; Ossa, 2017b):
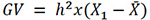 Equation 4
Equation 4
Where GV: genetic value; X1: each bovine’s average CI;
After estimating the repeatability value, the MPPA was calculated for each bovine female, whose formula is described in Equation 5.
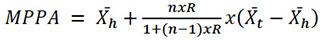 Equation 5
Equation 5
Where MPPA: most probable producing ability;
The estimation reliability or accuracy of the MPPA (AMPPA) is given by the following expression:
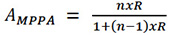 Equation 6
Equation 6
We established the linear correlation between the MPPA and the GV by initially classifying the bovines according to the value obtained for both parameters (from highest to lowest) and then calculating the Spearman’s correlation. Finally, the Romosinuano bovine females’ RE was calculated using Wilcox’s formula (1957), as stated in Equation 7.
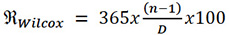 Equation 7
Equation 7
Where REWilcox: RE according to Wilcox; n: total number of births of the female bovine; D: age (in days) between the last and the first calving.
Results and discussion
The repeatability for the CI of Romosinuano bovine females belonging to the SBGNAA, Turipaná research center, was 0.02 ± 0.01 (table 2). The estimated value was low, indicating a low correlation between the females’ CIs; furthermore, as the standard error was less than the estimated repeatability, the study results can be used reliably.
Since the value of the estimated repeatability was low to improve the CI based on it, environmental effects would have to be considered; therefore, only females with CIs greater than 730 days should be discarded.
Table 2 Estimated repeatability and uncertainty associated with the estimated value for the CI variable of Colombian Romosinuano bovine females

1Confidence interval for repeatability, estimated using 1,000 bootstrap iterations and assuming the Gaussian distribution of the CI.
Source: Elaborated by the authors
The estimated repeatability value of the CI in this paper coincides in its low category with those found by other authors in other breeds, as follows: 0.05 in Brahman herd (Peña et al., 1979); 0.06 ± 0.02 in zebu (Magaña et al., 2002); 0.09 in Japanese black females (Oyama et al., 2002); 0.082 in Rubia Gallega females (Yagüe et al., 2009); 0.082 ± 0.07 in Brahman cattle (Osorio & Segura, 2010); 0.08 ± 0.01 in females used in a dual-purpose system (Galeano & Manrique, 2010); 0.0448 ± 0.0434, in beef cattle of the Angus × Nelore, Caracú × Nelore, and Valdostana × Nelore genetic groups (Silva et al., 2015).
Repeatability is a handy parameter for our bovine production systems’ circumstances since its major flaw is a lack of records. For its estimation, it is unnecessary to have a structured population relying on paternal half-sib groups, an essential mathematical trick to estimate heritability and then estimate animals’ GV for a particular trait since repeatability marks the limit of heritability. The foregoing helps estimate the animals’ most probable GV, whose expression is
As repeatability marks the limit of heritability, the latter can be less than or equal to the former, but never greater. Therefore, from the study, we can infer that heritability may have a maximum value of 0.02 or less, being very similar to the estimated value (0.04 ± 0.02) for this trait in the same breed, in a shorter period, and with fewer records (Ossa et al., 2007). Moreover, we can affirm that CI heritability in evaluated Romosinuano cows is equal to 0.02, meaning that the trait variation in the population depends on 2% of the genotype variations between individuals and 98% of other variations. It should be considered which environmental factors (e.g., calving year, month, or number) affect the trait significantly and, thus, propose management programs that induce its improvement.
The classification of the ten best and worst females of the herd, considering their MPPA and GV is shown in table 3. Note that the best cows have a CI less than 379,74 ± 47.74, which is the population’s average and, consequently, a negative GV. Instead, those included in the group of the worst ones have better MPPA, given by a CI greater than the population’s average and, consequently, a positive GV.
Table 3 Classification of the ten best and worst Romosinuano bovine females, according to their MPPA relative to the CI variable and relevant GV
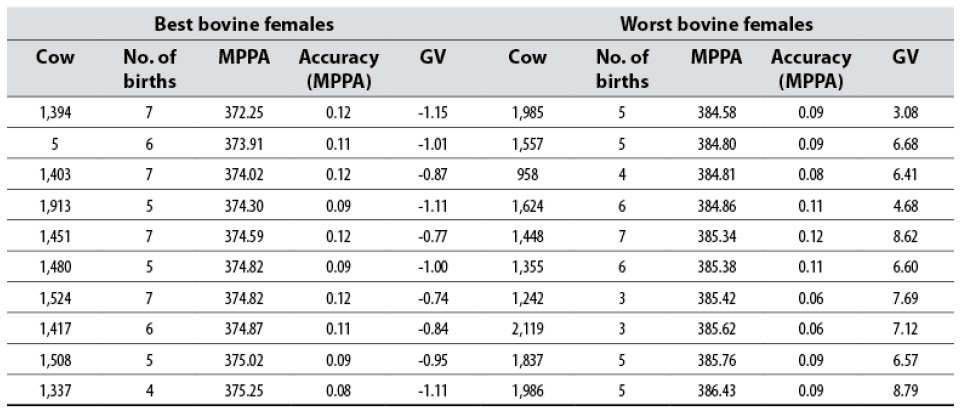
Source: Elaborated by the authors
For selection, we could use the MPPA or GV parameters due to their high positive correlation (~ 0.94 with p-value < 2.2 × 10-16), as shown in figure 1, coinciding with the results obtained by Quijano and Echeverri (2015).
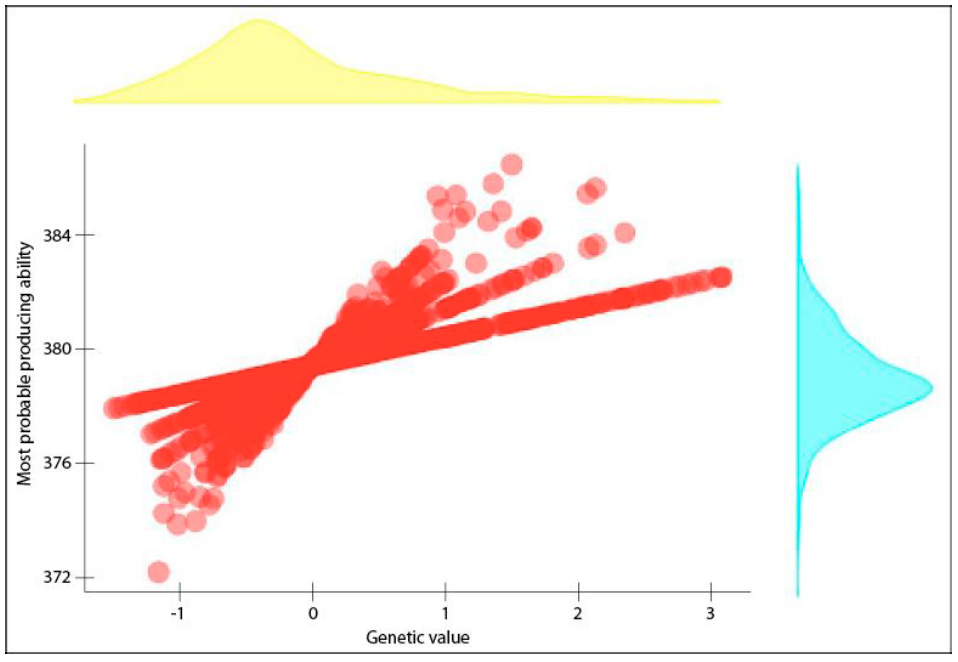
Source: Elaborated by the authors
Figure 1 Correlation between the MPPA and GV values for the CI variable of Colombian Romosinuano bovine females. The density plots show the distribution of both parameters
The MPPA makes it possible to predict the female bovine’s CI in this case regarding the next calving. Therefore, it can be a good indication for discarding females with CIs greater than the population’s average to improve this trait until achieving a good record structure that allows for more rigorous analysis.
The classification of 871 females analyzed according to their RE using fourteen classes with a class interval of four units is shown in table 4. The first class ranged from 53 to 57%, suggesting an average RE of 55%, i.e., a CI of 663.636 days (result obtained by calculating 365/0.55). Two cows had an RE between 118 and 122%, giving an average CI of 304.17 days (365/1.20). The highest concentration of RE was between 88 and 107%, i.e., 75% of the females whose CIs varied between 414.77 and 341.12 days. The general RE in the analyzed period was 96% (365/379.74); that is to say, of 100 cows exposed to the bull, 96 calves were produced.
Table 4 RE classes in which the Colombian Romosinuano bovine females were grouped with their relevant frequencies
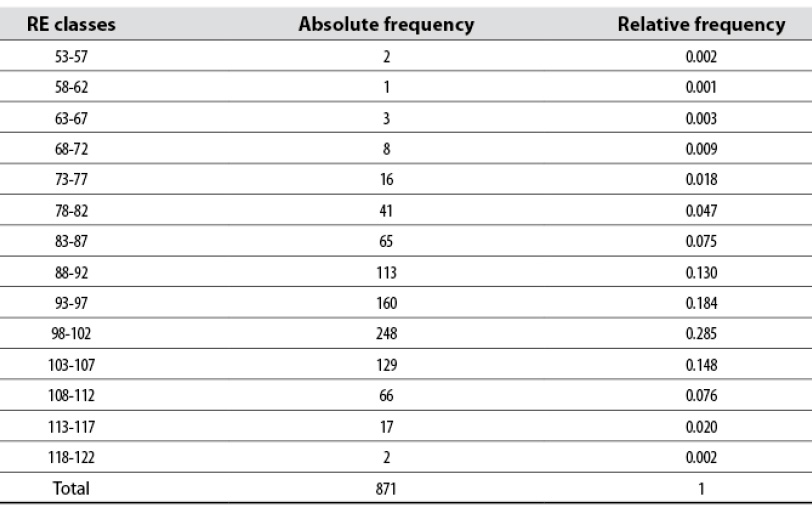
Source: Elaborated by the authors
One of the most outstanding traits of the Romosinuano breed is its degree of adaptation to the place of origin, Valle del Sinú, due to its high reproductive ability measured by the RE in this study. It is worth mentioning that its adaptation is not only reproductive-related but also in its ability to survive in this environment.
Conclusions
According to the analysis carried out in this paper, we can draw the following conclusions:
The CI repeatability in the Romosinuano females of the evaluated herd was low, indicating a low correlation between the cows’ individual records.
In the evaluations carried out in SBGNAA’s females, we noted that the MPPA becomes more reliable as the female produces more calves.
The SBGNAA’s females in Turipaná exhibited a high RE of 96% and an average CI value of
379.74 days, demonstrating Romosinuano females’ high RE and, therefore, adaptation to Valle del Sinú. So, it becomes an alternative for improving this characteristic in the national herd if used in crossbreeding.
Females can be selected for their RE or MPPA, given the high correlation.

