










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCiencia y Tecnología Agropecuaria
Print version ISSN 0122-8706On-line version ISSN 2500-5308
Cienc. Tecnol. Agropecuaria vol.22 no.2 Mosquera May/Aug. 2021 Epub May 01, 2021
https://doi.org/10.21930/rcta.vol22_num2_art:1814
Genética vegetal y biodiversidad
Evaluación fenotípica de poblaciones de Festuca arundinacea Schreber naturalizadas en ambientes subhúmedos semiáridos
*
1
 http://orcid.org/0000-0001-8564-1154
http://orcid.org/0000-0001-8564-1154
2
http://orcid.org/0000-0001-9358-468X
3
http://orcid.org/0000-0002-0609-332X
4
http://orcid.org/0000-0002-5830-5136
1Universidad Nacional de Río Cuarto. Río Cuarto
2Universidad Nacional de Río Cuarto. Río Cuarto
3Universidad Nacional de Río Cuarto. Río Cuarto
4Universidad Nacional de Río Cuarto. Río Cuarto
5Universidad Nacional de Río Cuarto. Río Cuarto
6Universidad Nacional de Río Cuarto. Río Cuarto
La festuca alta (Festuca arundinacea Schreber) es una forrajera perenne de regiones templadas importante en los sistemas de producción pecuarios debido a su elevado rendimiento en lugares con restricciones ambientales y edáficas. El objetivo del presente estudio fue evaluar el rendimiento fenotípico de diez poblaciones de festuca alta naturalizada en ambientes semiáridos subhúmedos de Argentina. Los caracteres fenotípicos altura de planta, diámetro de corona, número de macollos . producción de biomasa seca, se estimaron durante tres años. Los datos se analizaron mediante análisis de varianza, análisis de correlación, análisis de componentes principales y análisis de interacción población × ambiente. Los resultados sugieren que existe una elevada variabilidad fenotípica entre las diez poblaciones de festuca alta. Se registró interacción población × ambiente para todos los caracteres (p < 0,05). El análisis de correlación mostró que el carácter altura de planta puede ser útil para seleccionar indirectamente plantas superiores en producción de biomasa seca en cortes individuales y anuales. Las poblaciones 3243-645 y 3250-BAI se destacaron en los caracteres analizados y presentaron estabilidad a lo largo de los años. La alta variabilidad fenotípica encontrada resulta útil para ser explorada a futuro en el programa de mejoramiento y para lograr la identificación y selección de genotipos superiores dentro de poblaciones.
Palabras clave biomasa; caracteres; forrajera perenne; festuca alta; programa de mejoramiento
Tall fescue (Festuca arundinacea Schreber) is a temperate perennial forage important in livestock production systems due to its high yield in places with environmental and edaphic restrictions. The aim of this study was to evaluate the phenotypic performance of ten naturalized tall fescue populations in sub-humid semi-arid environments of Argentina. The phenotypic traits plant height, crown diameter, number of tillers, and dry biomass production were estimated during three years. The data were examined through analysis of variance, correlation analysis, principal component analysis, and population × environment interaction analysis. The results suggest that there is high phenotypic variability among the ten naturalized tall fescue populations. A genotype × environment interaction was recorded for all traits (p < 0.05). The correlation analysis showed that the plant height trait could be useful to select indirectly higher plants in dry biomass production in individual and annual cuts. Populations 3243-645 and 3250-BAI stood out in the analyzed traits presenting stability throughout the years. The high phenotypic variability found is valuable to be explored in the future in the breeding program and to identify and select superior genotypes within populations.
Keywords biomass; breeding program; forage; tall fescue; traits
Introducción
La producción agropecuaria en todo el mundo se encuentra fuertemente influenciada por la amplia variación de condiciones ambientales locales, lo que produce gran incertidumbre económica debido a los rendimientos inestables de los diferentes cultivos año tras año (Dehghani et al., 2016). Las regiones semiáridas son vulnerables a las variaciones climáticas debido a que tienen una distribución irregular de las precipitaciones que se concentran en unos pocos meses del año y varían entre y dentro de años. Los sistemas de producción agropecuaria deben adecuarse a las variaciones climáticas para aprovechar el recurso hídrico de manera eficiente (Ramos et al., 2016). En Argentina las regiones semiáridas tienen promedios de precipitaciones anuales que no reflejan su realidad de ocurrencia; años húmedos y secos se alternan de manera no previsible (Brendel et al., 2017). La región central de Argentina presenta un habitual déficit hídrico en los meses de invierno que, sumado a las limitaciones edáficas de los ambientes ganaderos, justifican la búsqueda de especies adaptadas a tales condiciones (Jauregui et al., 2017).
Resulta importante considerar que, además de las fluctuaciones climáticas propias de los ambientes semiáridos, la evaluación fenotípica de cultivos a campo es compleja, ya que siempre se analizan caracteres cuantitativos que se encuentran fuertemente influenciados por el ambiente. Los ensayos multiambientales (EMA) desempeñan un papel importante en el mejoramiento genético de los cultivos agronómicos debido a su utilidad para interpretar la interacción genotipo × ambiente (GxE). El diseño experimental y los análisis estadísticos de los ensayos EMA permiten eliminar la variabilidad contenida en los datos por causas que no se pueden explicar (Crossa, 1990). La metodología GGE biplot se desarrolló para analizar las bases de datos generadas por los EMA (Yan et al., 2000).
Las siglas GGE hacen referencia al efecto que los diferentes genotipos (G) tienen sobre los datos de producción, más el efecto que corresponde a la interacción genotipo × ambiente (GxE). Por otro lado, los análisis mediante el modelo de efecto aditivo e interacción multiplicativa (AMMI) permiten describir la respuesta adaptativa de los genotipos a través de diferentes ambientes (Gauch et al., 2008).
Festuca alta, Festuca arundinacea Schreber (Poaceae), es una especie forrajera hexaploide, con 2n = 6x = 42 cromosomas, perenne, de reproducción alógama y polinización anemófila (Hand et al., 2010). Estudios previos afirman que la amplia distribución geográfica de la festuca alta es consecuencia de su excelente adaptabilidad, conferida por características agronómicas favorables, tales como la resistencia al pastoreo, tolerancia a enfermedades e insectos (Petigrosso et al., 2019) y abióticos como la sequía (Dehghani et al., 2016). Por tales motivos, ocupa un lugar significativo en los sistemas de producción de carne y de leche de todo el mundo, debido a su rendimiento en lugares con restricciones ambientales y edáficas (Cougnon et al., 2018).
La especie forrajera perenne F. arundinacea se encuentra naturalizada en diferentes ambientes de la región central subhúmeda semiárida de la Argentina (Scheneiter et al., 2016). La cátedra de genética perteneciente al departamento de Biología Agrícola de la Facultad de Agronomía y Veterinaria de la Universidad Nacional de Río Cuarto comenzó un programa de mejora de festuca alta, mediante la colecta de plantas adultas naturalizadas en la región. A partir de dicha colecta, se formaron diez poblaciones que se estudian con fines de mejoramiento. Estas se caracterizaron en campo por sus caracteres vegetativos y reproductivos (Vega et al., 2018). El objetivo del presente estudio fue evaluar el desempeño fenotípico de las diez poblaciones de festuca alta naturalizada en la región central subhúmeda semiárida de la Argentina.
Materiales y métodos
Material vegetal y condiciones de crecimiento
El estudio se desarrolló en el Campo Experimental de la Facultad de Agronomía y Veterinaria de la Universidad Nacional de Río Cuarto (33°06′22″S, 64°17′52″O, 443 m s.n.m.), localizada al sur de la provincia de Córdoba, Argentina, durante los años 2014, 2015 y 2016. El suelo del campo experimental es un Haplustol típico con un contenido promedio de materia orgánica de 1,6%, 27 ppm de nitratos y 24 ppm de fósforo en los primeros 20 cm de suelo. El clima de la región es subhúmedo-semiárido, con un valor medio anual de precipitaciones de 800 mm y el 80% de estas concentrado en el semestre más cálido, desde octubre hasta marzo. El área cuenta con una temperatura media anual de 16,5 °C, una temperatura máxima media anual de 22,8 °C y una temperatura mínima media anual de 10,2 °C (periodo 1981-2010). De 2014 a 2016 se presentaron valores de temperatura media mensual similares a los valores de periodo 1981-2010, pero las precipitaciones presentaron mayor variabilidad en comparación con dicho periodo (figura 1).
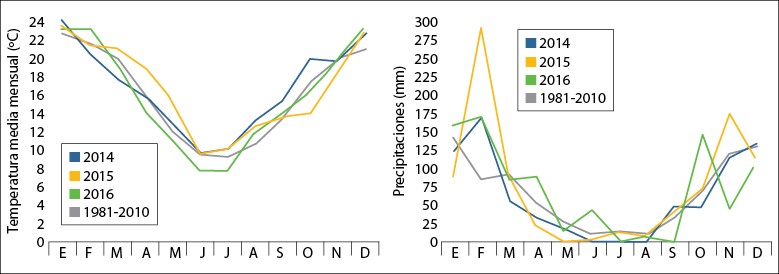
Fuente: Elaboración propia
Figura 1. Temperaturas (izquierda) y precipitaciones (derecha) medias mensuales registradas en los años 2014, 2015 y 2016 y de la serie 1981-2010 en Río Cuarto, Córdoba, Argentina.
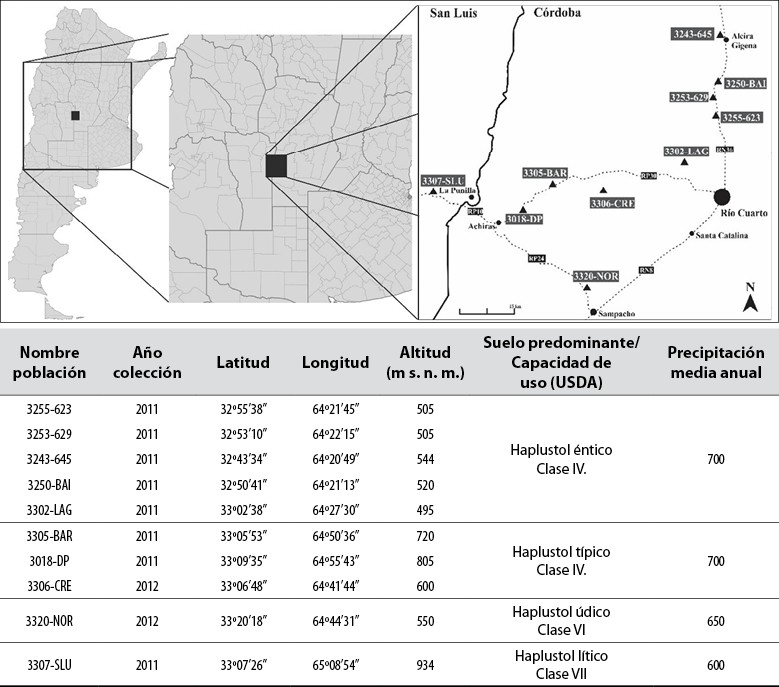
El material vegetal consistió en diez poblaciones naturalizadas colectadas en el sur de la provincia de Córdoba y norte de San Luis, Argentina (figura 2), a las orillas de caminos y rutas, así como en pasturas de prolongado tiempo bajo pastoreo. Además, se utilizaron dos cultivares comerciales: Pergamino El Palenque MAG (Palenque-INTA), cultivar de amplia utilización y bien adaptado a la región, y Balerón- Forratec (Balerón-Forr), cultivar nuevo de rendimientos promisorios, y dos colecciones (017 y 509) de comportamiento diferencial en cuanto a acumulación de biomasa provenientes del Banco de Germoplasma del Instituto Nacional de Tecnología Agropecuaria, ubicado en la Estación Experimental Pergamino (Rosso et al., 2007). El ensayo en campo se implantó en marzo de 2014, en condiciones de secano y sin aplicación de fertilizantes en ningún momento del ciclo. El control de malezas se realizó de forma manual. Al final de cada ciclo del cultivo se llevaron a cabo cortes de limpieza. Se utilizó un diseño en bloques completos aleatorizados. Además, las plantas de cada población se clonaron para conformar cuatro repeticiones, donde cada población fue dispuesta al azar dentro de cada bloque, en parcelas con 24 plantas cada una. Cada parcela consistió en tres surcos distanciados entre sí por 25 cm, con ocho plantas por surco distanciadas por 50 cm entre plantas.
Caracteres fenotípicos
Los caracteres se estimaron a través de cortes sucesivos de forraje. Para distanciarlos durante el ciclo de crecimiento, se consideró el tiempo térmico de senescencia foliar (cada 550 °C días) y una temperatura base de 5 °C (Chapman & Lemaire, 1993). Se realizaron tres cortes en 2014 y cuatro en los años 2015 y 2016, en los meses de marzo (primer corte), abril-mayo (segundo corte), julio-agosto (tercer corte), octubre-noviembre (cuarto corte). Los caracteres fenotípicos considerados en cada corte por planta fueron altura de planta (AP), diámetro de corona (DC), número de macollos (NM) y producción de biomasa seca promedio por corte por planta (BSC). Además, se calculó la biomasa seca total producida en el año mediante la suma de los cortes (BST). La AP se midió antes de cada corte y el DC después, en centímetros (cm) con regla graduada sin modificación de la estructura de la planta (medidas no destructivas). El corte de biomasa aérea de cada planta se realizó con una tijera mecánica, dejando una altura de 5 cm de rastrojo remanente. La biomasa verde se utilizó para determinar el NM en número de macollos por planta. Luego se pesó y se secó en estufa de aire forzado hasta tener peso constante, para obtener la BSC en gramos/planta (medidas destructivas).
Análisis estadísticos
Los caracteres fenotípicos se estudiaron mediante análisis de varianza (ANOVA), utilizando el siguiente modelo: Υ𝑖𝑗𝑘𝑙 = 𝜇 + 𝛼𝑖 + 𝛽𝑗 + 𝛾𝑘 + 𝛿𝑙 + 𝛼𝑖 × 𝛿𝑙 + 𝜀𝑖𝑗𝑘𝑙, donde 𝛶ijkl es el valor promedio de la población i en la repetición j en el corte k en el año l, µ es la media general del ensayo; αi, βj, γk y δl son los efectos principales debido a la población, repetición, corte y año, respectivamente; αi x δl corresponde al efecto de la interacción población × año (GA) y εijkl es el término del error (Ospina et al., 2012). Las poblaciones (G), el año (A) y la interacción población × año (GA) se consideraron como efectos fijos, mientras que las variables repetición y corte se consideraron como aleatorias. La correlación lineal entre caracteres se analizó utilizando coeficientes de correlación de Pearson (Pearson, 1920).
Un análisis de componentes principales (ACP) se realizó incluyendo los tres años de evaluación. Los caracteres fenotípicos además se analizaron mediante el modelo de regresión por sitio (SREG) con gráficos biplot GGE. Los biplot GGE se generaron a partir de la técnica de descomposición del valor singular (SVD) de la matriz de residuos del siguiente modelo: Υ𝑖𝑗 − Υ𝑗 = 𝜆1 𝜉𝑖1𝜂𝑗1 + 𝜆2𝜉𝑖2𝜂𝑗2 + 𝜀𝑖𝑗, donde Υij es el rendimiento medio de la población i en el año j; Υj es la media de las poblaciones en el año j; λ1 y λ2 son los autovalores para la CP1 y CP2, respectivamente; ξi1 y ξi2 son los scores de la población i en la CP1 y CP2, respectivamente; ηj1 y ηj2 son los scores del año j en la CP1 y CP2, respectivamente, y εij es el término residual asociado a la observación promedio de la población i en el año j centrado por el efecto del año j.
El carácter BSC se analizó mediante el modelo de efecto aditivo e interacción multiplicativa (AMMI): Υ𝑖𝑗𝑟 = 𝜇 + 𝛼𝑖 + 𝛽𝑗 + ∑ 𝜆𝑛 𝛾𝑖𝑛 𝛿𝑗𝑛 + 𝜀𝑖𝑗𝑟, donde Υijr es el rendimiento de la población i en el año j para la repetición r. Los parámetros aditivos son .(media general del ensayo combinado), αi y βj son los efectos principales debido a la población y el año, respectivamente, mientras que λn corresponde al autovalor para el eje . del ACP; γin, al autovector de las poblaciones para el eje n; δjn, al autovector de años y, finalmente, εijr es el término del error (Gauch et al., 2008). La metodología se basó en el ajuste de los efectos aditivos del modelo (población y año) mediante un ANOVA, del que se obtuvieron los residuos que contienen solo los efectos de la interacción (población × año). Luego, dicha matriz de residuos se analizó mediante un ACP. El componente principal uno (CP1) del ACP se utilizó para generar el gráfico resultado del modelo AMMI1. Se trabajó con los software estadísticos Infostat (Di Rienzo et al., 2018) y su vinculación con R (R Core Team, 2017) e Infogen (Balzarini & Di Rienzo, 2017).
Resultados y discusión
El ANOVA reveló que las poblaciones difirieron significativamente y que se encontró interacción GA (p < 0,05) en todos los caracteres. La interacción puede explicarse por el comportamiento desigual de las poblaciones ante las diferentes condiciones ambientales ocurridas durante los tres años de evaluación (figura 1). Estudios previos realizados sobre el mismo material vegetal en un año de evaluación confirman la presencia de diferencias entre poblaciones (Vega et al., 2018). Chtourou-Ghorbel et al. (2011), al igual que en el presente estudio, reportan la existencia de diferencias estadísticamente significativas entre poblaciones e interacción GA, significativa en producción de biomasa seca promedio por corte sobre un grupo de 35 poblaciones de festuca alta, recolectadas en diferentes ambientes de Túnez, en donde también se observa una elevada variabilidad de características edafoclimáticas.
Los valores promedio de cada corte para los tres años de evaluación se resumen en la tabla 1. El carácter DC presentó valores promedio que oscilaron entre 13,09 ± 0,15 cm y 15,34 ± 0,15 cm en los diferentes cortes realizados. En Isfahan, Irán, región donde la precipitación anual es de 140 mm, Ebrahimiyan et al. (2012) observaron en secano valores promedio inferiores a los hallados en el presente estudio (10 ± 1,8 cm), mientras que con irrigación los valores (13,70 ± 1,4) fueron similares a los informados en los datos que se discuten, donde las precipitaciones fueron de alrededor de 800 mm anuales en secano.
Tabla 1. Valor promedio y error estándar (EE) de los caracteres fenotípicos medidos en cada corte de forraje en el periodo 2014-2016, valores F y p de la interacción población × año (GA) del análisis de la varianza de las poblaciones de Festuca arundinacea evaluadas en Río Cuarto, Córdoba, Argentina

Nota.EE: error estándar; F: estadístico de análisis de varianza; p: valor de probabilidad; AP: altura de planta (cm); DC: diámetro de la corona (cm); NM: número de macollos (macollo/planta); BSC: producción de biomasa seca promedio por corte (g/planta); sd: sin datos.
Fuente: Elaboración propia
Por otro lado, el número de macollos por planta (NM) promedio del presente trabajo osciló entre 38,45 ± 0,62 en la población 3250-BAI y 46,46 ± 0,69 en la población 3243-645 (promedios de todos los cortes y años de evaluación). Dichos valores resultan inferiores a los hallados por Ebrahimiyan et al. (2012), quienes informaron valores promedio de 63,40 ± 10,3 macollos por planta bajo riego y de 51,60 ± 13 en condiciones de secano. Estas diferencias quizá se deban, al menos en parte, al uso de genotipos más macolladores en Isfahan, Irán. Además, dado que las precipitaciones anuales representan solo un 17% (140 mm anuales) en relación con las que ocurren en la región central subhúmeda semiárida de Argentina (800 mm anuales), probablemente las plantas recurran a la estrategia de producir mayor cantidad de macollos en desmedro de su tamaño, para asegurar una mayor exploración horizontal del suelo en búsqueda de agua.
Respecto a la biomasa seca por planta (BSC), en el 2015 fue superior al segundo corte del 2014 y al primero y segundo del 2016 (tabla 1). Este hecho puede explicarse por las abundantes precipitaciones ocurridas durante febrero del 2015 (figura 2). La BSC del 3.º y 4.º corte fue superior en el 2014 (tabla 1), probablemente atribuible a las temperaturas superiores a la media, ocurridas durante la segunda mitad del año, y a la ocurrencia de mayores precipitaciones durante el mes de septiembre (50% superiores a la media histórica de la región) (figura 1).
La biomasa seca total producida por planta por año (BST) presentó valores promedio de 82,36 ± 1,38 g en 2014, 122,13 ± 2,47 g en 2015 y 63,91 ± 1,06 g en 2016. El carácter presentó mayor EE en 2014 y 2015, lo que pudo estar asociado a la mayor variación de la temperatura y precipitaciones, respectivamente (figura 1).
Pirnajmedin et al. (2016) en Isfahan, Irán, encontraron que el rango de producción de biomasa osciló entre 30,58 g y 248,58 g en un ensayo ya citado. Por su parte, los valores del ensayo que se discute se sitúan entre los observados en el ensayo anterior (desde 74,81 ± 2,59 g en la población 3307-SLU a 101,06 ± 4,74 g en la población 3250-BAI). Los valores elevados de BST encontrados por Pirnajmedin et al. (2016) probablemente se deban a las fertilizaciones anuales del ensayo con fósforo y nitrógeno; sin embargo, las condiciones edafoclimáticas más benignas de Río Cuarto habrían permitido obtener rendimientos más estables que en Isfahan, Irán, aún con las diferencias fenotípicas entre las poblaciones evaluadas en el presente trabajo.
Correlación entre caracteres fenotípicos y comportamiento agronómico de las poblaciones de festuca alta
Este enfoque puede proporcionar información importante para mejorar el rendimiento en forraje de festuca alta (Ebrahimiyan et al., 2012). Se encontraron valores de correlación positiva entre varios caracteres estimados (tabla 2). Los valores más altos y significativos ocurrieron entre NM vs. DC, AP vs. BST y AP vs. BSC. Estas correlaciones sugieren que el carácter altura de planta (AP) puede ser útil para seleccionar plantas superiores en producción de biomasa (BSC y BST). Probablemente, debido a que la AP presenta una alta correlación directa positiva con BSC, se produce una disminución del efecto de correlación positiva directa entre NM y BSC. Otros estudios también informan la presencia de correlaciones positivas entre producción de biomasa seca y altura de planta (Chtourou-Ghorbel et al., 2011), así como entre número de macollos (NM) y diámetro de corona (DC) (Pirnajmedin et al., 2016).
Tabla 2. Coeficientes de correlación de Pearson entre los caracteres fenotípicos medidos en el conjunto de los once cortes de forraje de Festuca arundinacea durante los años 2014 2015 y 2016 Río Cuarto Córdoba Argentina

El ACP proporciona un resumen gráfico del conjunto de caracteres medidos, la relación entre ellos y el comportamiento de las poblaciones en relación con ellos. El biplot presentado en la figura 3 muestra que las CP1 y CP2 explicaron un 92,4% de la variabilidad entre las poblaciones, colecciones y cultivares (correlación cofenética: 0,994). Las poblaciones 3202-LAG, 3250-BAI y los cultivares comerciales Palenque-INTA y Balerón-Forr fueron los que mejor se asociaron con los caracteres AP, BSC, y BST. Por otro lado, la población 3243-645 y la colección 509 se asociaron en forma positiva con NM y DC.
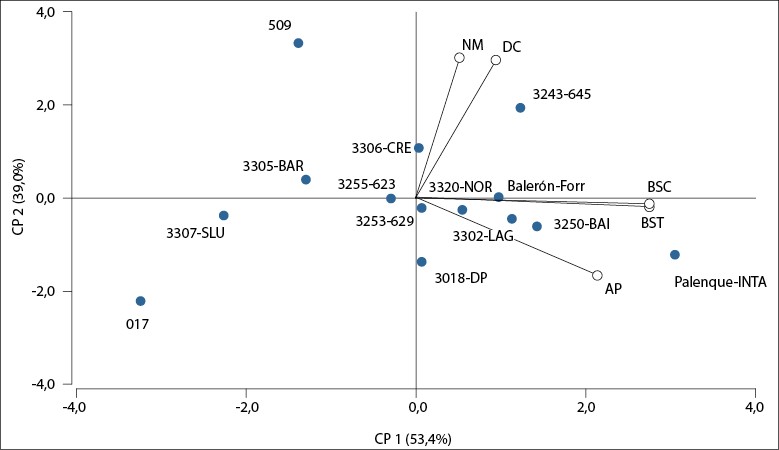
Fuente: Elaboración propia
Figura 3. Biplot del análisis de componentes principales para los cinco caracteres medidos en las diez poblaciones naturalizadas, dos cultivares comerciales y dos colecciones de Festucaarundinacea en Río Cuarto, Córdoba, Argentina. 2014-2016. AP: altura de planta (cm); DC: diámetro de corona (cm); NM: número de macollos (macollo/planta); BSC: biomasa seca producida por corte (g/planta); BST: biomasa seca total producida por año (g/planta).
Estudio de la interacción población × año
En la figura 4 se pueden observar los gráficos resultantes del modelo GGE aplicado en los diferentes caracteres medidos. El modelo GGE para los caracteres AP, DC y NM explicó, respectivamente, el 93%, 98% y 95% de la variación debida a los efectos combinados de poblaciones e interacción población × año. En el biplot de AP se observa que quedaron conformados cinco megambientes (delimitados por líneas de puntos), y que los tres años quedaron incluidos dentro del mismo megambiente. El cultivar Palenque-INTA se ubicó en el vértice del polígono, es decir, presentó el mejor comportamiento en AP en los tres años de evaluación. Otras poblaciones destacadas en este carácter fueron 3302-LAG, 3018-DP, 3320-NOR y 3250-BAI, junto al cultivar Balerón-Forr.
La población 3243-645 y la colección 509 se destacaron en los biplot de DC y NM en 2015 y 2016. La población 3306-CRE presentó un comportamiento superior en NM en 2014, indicado por la ubicación en el extremo del polígono del megambiente que incluye dicho año. El biplot GGE del carácter BSC explicó el 94% de la variabilidad, en el cual Palenque-INTA y la población 3302-LAG (ubicados en los vértices del polígono), junto a cuatro poblaciones (3250-BAI, 3243-645, 3306-CRE, 3253-629, 3320- NOR), se destacaron en 2014 y 2015.
Los biplot GGE de los caracteres fenotípicos determinaron que el efecto de las diferentes poblaciones (G) es mayor que el efecto de la interacción población × año (GA), ya que en todos los gráficos quedaron definidos de cinco a seis megambientes, y los diferentes años de evaluación quedaron incluidos en solo uno o dos megambientes (figura 4).
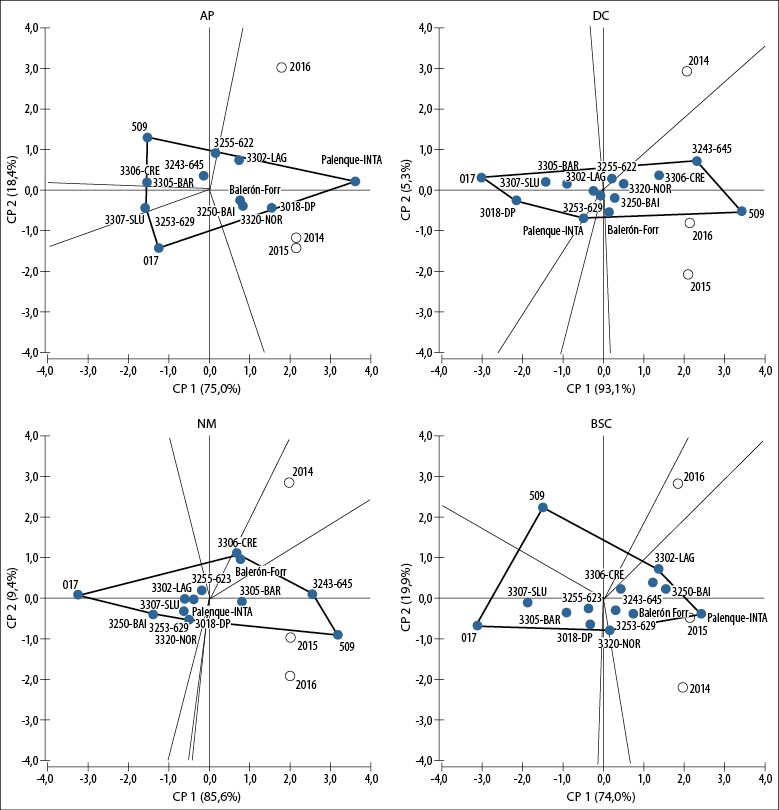
Fuente: Elaboración propia
Figura 4. Biplots revelados por el modelo GGE para los caracteres estimados en las poblaciones, colecciones y cultivares de Festuca arundinacea en Río Cuarto, Córdoba, Argentina. 2014-2016. AP: altura de planta (cm); DC: diámetro de corona (cm); NM: número de macollos (macollo/planta); BSC: biomasa seca producida bajo corte (g/planta).
Dehghani et al. (2015) analizaron el comportamiento de 24 genotipos de festuca alta en 14 ambientes diferentes en ensayos en campo conducidos en Irán. A través de gráficos biplot surgidos del modelo GGE, los investigadores pudieron definir megambientes y genotipos con mejor comportamiento en cada megambiente. En ese caso el biplot GGE explicó un 75% de la variación entre los genotipos evaluados para producción de biomasa, mientras que el GGE de BSC del presente estudio explicó una mayor variabilidad entre las poblaciones evaluadas, logrando identificar poblaciones superiores en determinados megambientes con un mayor grado de confianza.
Estabilidad y adaptabilidad de las poblaciones de festuca alta
El modelo AMMI1 es de utilidad para interpretar la estabilidad de las poblaciones en los diferentes años y su adaptabilidad (Gauch et al., 2008). En la figura 5 se muestra el biplot AMMI1, donde la CP1, que indica la estabilidad de las poblaciones (eje Y), explica el 58% de la interacción GA. Por su parte, el eje X indica su adaptabilidad a través del BSC promedio de cortes en tres años de evaluación. El cultivar Palenque-INTA presentó la mayor BSC relativa al resto de cultivares, colecciones y poblaciones, aunque con la menor estabilidad a través de los años de evaluación, mientras que las colecciones 017, 509 y la población 3307-SLU fueron las de menor BSC promedio. Estas también presentaron alta inestabilidad.
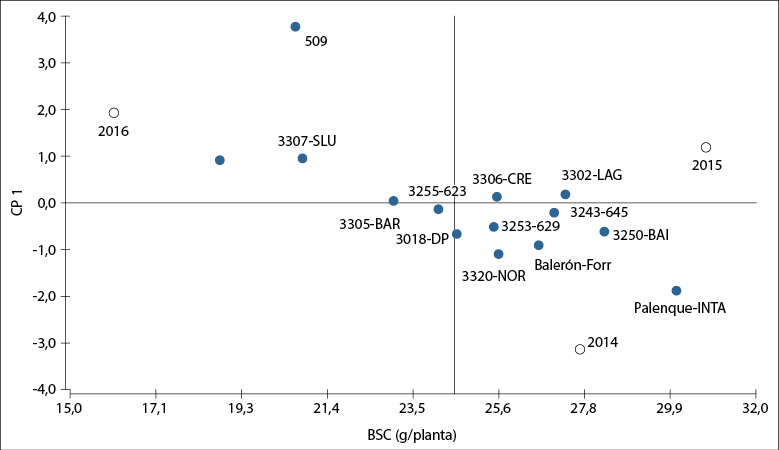
Fuente: Elaboración propia
Figura 5. Biplot AMMI1 que muestra poblaciones naturalizadas, colecciones, cultivares comerciales de Festucaarundinacea (círculos rellenos) y los años de evaluación 2014, 2015 y 2016 (círculos vacíos). Valor promedio general de producción de biomasa seca promedio por corte (BSC): 24,54 g/planta. Río Cuarto, Córdoba, Argentina.
Las poblaciones 3306-CRE, 3253-629, 3243-645, 3302-LAG y 3250-BAI presentaron valores promedio de BSC superiores al promedio general (24,5 g/planta) y resultaron estables a través de los años (presentaron valores cercanos a cero en la CP1 de la figura 5). Dehghani et al. (2016), en su estudio sobre 72 genotipos de festuca alta evaluados en 16 ambientes, informan que el modelo AMMI1 explica 48% de la interacción GA, mientras que en el presente trabajo fue posible explicar un mayor porcentaje de la interacción de GA (58%), sugiriendo que la metodología empleada logró captar una mayor cantidad de variabilidad entre las poblaciones evaluadas durante los tres años de ensayo y, por lo tanto, mejora la confianza en la identificación de poblaciones superiores.
Conclusiones
Los resultados del presente estudio sugieren que existe una gran variabilidad fenotípica entre las diez poblaciones de festuca alta naturalizadas en la región central subhúmeda semiárida de Argentina. Los análisis de correlaciones demostraron que el carácter altura de planta (AP) puede resultar útil para seleccionar indirectamente plantas superiores en producción de biomasa (BSC y BST). Con los análisis GGE, AMMI1 y ACP, se encontró que las poblaciones 3243-645 y 3250-BAI se destacaron en los caracteres evaluados, presentando además una adecuada adaptabilidad y estabilidad en producción de biomasa a través de los años. A pesar de que no se hallaron poblaciones superiores a los cultivares incluidos en el ensayo, se lograron identificar poblaciones que los superaron en estabilidad de producción de biomasa. Por otro lado, la amplia variabilidad fenotípica encontrada puede ser utilizada para continuar el programa de mejora y hallar genotipos superiores a los cultivares.
Agradecimientos
Los autores agradecen a la Universidad Nacional de Río Cuarto por facilitar los recursos e instalaciones para realizar esta investigación.
REFERENCIAS
Balzarini, M. G., & Di Rienzo, J. A. (2017). InfoGen versión 2017 [software]. FCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
Brendel, A. S., Bohn, V. Y., & Piccolo, M. C. (2017). Variabilidad de la precipitación y su relación con los rendimientos agrícolas en una región semiárida de la llanura pampeana (Argentina). Estudios Geográficos, 78(282), 7-29. http://doi.org/10.3989/estgeogr.201701 [ Links ]
Chapman, D. F., & Lemaire, G. (1993). Morphogenetic and structural determinants of plant regrowth after defoliation. In Proceedings of the XVII International Grassland Congress, Palmerston North and Rockhampton, 1993 (pp. 95-104). Sir Publishing. [ Links ]
Chtourou-Ghorbel, N., Chakroun, M., Elazreg, H., & Trifi-Farah, N. (2011). Agronomic evaluation and genetic variation of Tunisian tall fescue (Festuca arundinacea Schreb.). International Journal of Agronomy, 2011, Article ID 349240. http://doi.org/10.1155/2011/349240 [ Links ]
Cougnon, M., Shahidi, R., Schoelynck, J., Van Der Beeten, I., Van Waes, C., De Frenne, P., & Reheul, D. (2018). Factors affecting grazing preference by sheep in a breeding population of tall fescue (Festuca arundinacea Schreb.). Grass and Forage Science, 73(2), 330-339. http://doi.org/10.1111/gfs.12340 [ Links ]
Crossa, J. (1990). Statistical analyses of multilocation trials. Advances in agronomy, 44, 55-85. https://doi.org/10.1016/S0065-2113(08)60818-4 [ Links ]
Dehghani, M. R., Majidi, M. M., Saeidi, G., Mirlohi, A., Amiri, R., & Sorkhilalehloo, B. (2015). Application of GGE biplot to analyse stability of Iranian tall fescue (Lolium arundinaceum) genotypes. Crop and Pasture Science, 66(9), 963-972. http://doi.org/10.1071/CP15043 [ Links ]
Dehghani, M. R., Majidi, M. M., Mirlohi, A., & Saeidi, G. (2016). Study of genotype by environment interaction in tall fescue genotypes and their polycross progenies in Iran based on AMMI model analysis. Crop and Pasture Science, 67(7), 792-799. http://doi.org/10.1071/CP15386 [ Links ]
Di Rienzo, J. A., Casanoves, F., Balzarini, M. G., Gonzalez, L., Tablada, M., & Robledo, C. W. (2018). InfoStat versión 2018 [software]. Centro de Transferencia InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
Ebrahimiyan, M., Majidi, M. M., & Mirlohi, A. (2012). Genotypic variation and selection of traits related to forage yield in tall fescue under irrigated and drought stress environments. Grass and Forage Science, 68(1), 59-71. https://doi.org/10.1111/j.1365-2494.2012.00869.x [ Links ]
Gauch, H. G., Piepho, H. P., & Annicchiarico, P. (2008). Statistical analysis of yield trials by AMMI and GGE: Further considerations. Crop Science, 48(3), 866-889. https://doi.org/10.2135/cropsci2007.09.0513 [ Links ]
Hand, M. L., Cogan, N. O., Stewart, A. V., & Forster, J. W. (2010). Evolutionary history of tall fescue morphotypes inferred from molecular phylogenetics of the Lolium-Festuca species complex. BMC Evolutionary Biology, 10(1), 303. https://doi.org/10.1186/1471-2148-10-303 [ Links ]
Jauregui, C. G., Ruiz, M. D. L. A., & Ernst, R. D. (2017). Tolerancia a la salinidad en plántulas de agropiro criollo (Elymus scabrifolius) y agropiro alargado (Thinopyron ponticum). Pastos y Forrajes, 40(1), 29-36. http://scielo.sld.cu/pdf/pyf/v40n1/pyf04117.pdf [ Links ]
Ospina, S., Rusch, G. M., Pezo, D., Casanoves, F., & Sinclair, F. L. (2012). More stable productivity of semi natural grasslands than sown pastures in a seasonally dry climate. PLoS One, 7(5). http://doi.org/10.1371/journal.pone.0035555 [ Links ]
Pearson, K. (1920). Notes on the history of correlation. 1920. Biometrika, 13(1), 25-45. http://doi.org/10.1093/biomet/13.1.25 [ Links ]
Petigrosso, L. R., Gundel, P. E., Colabelli, M. N., Fernández, O. N., & Assuero, S. G. (2019). Hongos endófitos en festuca alta: del problema a las soluciones. Revista de Investigaciones Agropecuarias, 45(2), 292-303. http://ppct.caicyt.gov.ar/index.php/ria/article/view/14413/45454575768596 [ Links ]
Pirnajmedin, F., Majidi, M. M., Mirlohi, A., & Noroozi, A. (2016). Application of EST-derived microsatellite markers for analysis of genetic variation in tall fescue and its comparison with morphological markers. Biochemical Systematics and Ecology, 65, 225-233. http://doi.org/10.1016/j.bse.2016.02.026 [ Links ]
R Core Team. (2017). R: A language and environment for statistical computing [software]. R foundation for Statistical Computing. [ Links ]
Ramos, J. P. F., Santos, E. M., Santos, A. P. M., de Souza, W. H., & Oliveira, J. S. (2016). Ensiling of Forage Crops in Semiarid Regions. In T. Da Silva, & M. E. Santos (Eds.), Advances in Silage Production and Utilization (pp. 65-84). IntechOpen. http://doi.org/10.5772/65446 [ Links ]
Rosso, B. S., Rimieri, P., Carrete, J., Cattoni, M. I., Biagoli, C., Cuyeu, A. R., Pagano, E. M., Ríos, R. D. (2007). Caracterización agronómica, molecular y de la calidad nutricional de una colección de festuca alta (Festuca arundinacea Schreb.) del Banco de Germoplasma de Pergamino, Argentina. En T. Da Silva, & M.E. Santos (Eds.), Avances de investigación en recursos genéticos en el Cono Sur II (pp. 173- 179). Procisur. https://www.researchgate.net/publication/288817218 [ Links ]
Scheneiter, J. O., Kaufmann, I. I., Ferreyra, A. R., & Llorente, R. T. (2016). The herbage productivity of tall fescue in the Pampas region of Argentina is correlated to its ecological niche. Grass and Forage Science, 71(3), 403-412. http://doi.org/10.1111/gfs.12184 [ Links ]
Vega, D. J., di Santo, H., Grassi, E., Castillo, E., Ferreira, V., Bonamico, N., & Ferreira, A. (2018). Diferenciación fenotípica de poblaciones de festuca alta naturalizadas en la región central de Argentina. Ab Intus, 1(1), 1-8. http://www.ayv.unrc.edu.ar/ojs/index.php/Ab_Intus/article/view/47/8 [ Links ]
Yan, W., Hunt, L. A., Sheng, Q., & Szlavnics, Z. (2000). Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Science, 40(3), 597-605. https://doi.org/10.2135/cropsci2000.403597x [ Links ]
Recibido: 11 de Febrero de 2020; Aprobado: 03 de Diciembre de 2020
 Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.
Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.
