











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
Permalink
Introducción
El agave es una planta emblemática y de gran importancia socioeconómica para México y el estado de Michoacán. Se utiliza como forraje, medicina y fibra, es fuente de alimento para el ganado y humano, y como material de construcción, entre otros usos; pero su principal utilidad es para la elaboración de bebidas alcohólicas como el pulque, el mezcal (Enríquez-del Valle et al., 2018), la bacanora y el tequila, producido con Agave tequilana Weber var. Azul (García- Mendoza, 2004).
Comúnmente, esta planta tarda entre 7 y 8 años para cosecharse, aunque puede llegar hasta los 12 años, por lo que se ha tratado de reducir el tiempo de cosecha por medio de diferentes prácticas agrícolas: principalmente, mediante el manejo de la fertilización química, los riegos controlados (Martínez-Ramírez et al., 2012; Enríquez-del Valle et al., 2018) y el uso de recursos biológicos, como lo son los hongos micorrízicos arbusculares (HMA) (Montoya-Martínez et al., 2014; Quiñones-Aguilar et al., 2016; Trinidad-Cruz et al., 2017b) y combinación de fertilizaciones químicas y recursos microbianos (Zacarías-Toledo et al., 2016; García-Martínez et al., 2020). Generalmente esta especie se propaga mediante la propagación vegetativa de hijuelos rizomáticos o bulbilos aéreos, siendo estos métodos un potencial movilizador de enfermedades como Fusarium (Mendoza-Ramos et al., 2021), el cual es uno de los mayores problemas fitosanitarios que enfrenta el cultivo, esta enfermedad es comúnmente conocida como “marchitez” y es provocada por Fusarium oxysporum (Ramírez-Ramírez et al., 2017; Sierra-Gómez et al., 2019; López-Bautista et al., 2020; Mendoza-Ramos et al., 2021).
El síntoma característico es la marchitez de las hojas, ocasionado por la destrucción del sistema radical o bien por el taponamiento de los haces vasculares. En campo, las plantas presentan clorosis, enrollamiento de los bordes de las hojas, secado de las hojas más viejas del ápice a la base, pudrición extensiva de color marrón en la corona y pudrición de las raíces (Ramírez- Ramírez et al., 2017; Campos-Rivero et al., 2018). En las plantas de vivero no se cuenta con una caracterización de la sintomatología de esta enfermedad y su control es principalmente mediante la aplicación de productos fúngicos foliares a base de cobre, en toda la planta y directamente en el cogollo y el suelo; sin embargo, es poco efectivo su control, debido a que el problema se encuentra en la raíz y el fungicida difícilmente llega hasta ellas (Soltero-Quintana, 2002).
Por lo anterior se han buscado otras alternativas de control que sean más efectivas, ecológicas y sustentables, por lo que el uso de microorganismos como biocontrol o bioprotección es una opción prometedora. Se sabe que la utilización de HMA puede prevenir el ataque de bacterias (Zhu & Yao, 2004) y hongos fitopatógenos asociados a las enfermedades que afectan el agave (Trinidad-Cruz et al., 2014; 2017b) y la prevención de enfermedades mediante el uso de HMA tiene sus bases en que esta simbiosis da como resultado plantas mejor nutridas y vigorosas, por la adquisición de nutrientes poco disponibles para las plantas, pero accesibles a los hongos micorrízicos (Riveros-Angarita, 2010; Ujvári et al., 2021).
Existen diversos estudios donde se muestra la efectividad de los HMA como promotores del crecimiento vegetal y bioprotector contra enfermedades causadas por Fusarium oxysporum en diversos cultivos. En gladiolas (Gladiolus grandiflorus), Khalil-Gardezi et al. (2001) mencionan que las que son inoculadas con Glomus spp. obtienen un mayor crecimiento vegetativo y tienen un menor daño radical ocasionado por Fusarium. Al usar Glomus caledonium y un consorcio de Glomus spp. y Acaulospora spp. en pepino inoculado con F. oxysporum f. sp. cucumerimun, Jun-Li et al. (2010) reportan un mayor crecimiento y un menor daño por Fusarium en plantas con HMA. En palma datilera (Phoenix dactylifera), Jaiti et al. (2007) mostraron la efectividad de Glomus monosporus, G. deserticola, G. clarum y un consorcio nativo, como promotores de crecimiento y protección contra Fusarium oxysporum f. sp. albedinis. Particularmente en agaváceas, Trinidad-Cruz et al. (2017b) reportaron que plantas de Agave cupreata micorrizadas con un consorcio nativo y uno comercial de HMA e infestados con Fusarium Oxysporum, mostraron un incremento en el crecimiento de los agaves micorrizados, además de una disminución de más del 50 % de severidad de la enfermedad respecto a agaves sin micorrización. Estos autores concluyen que la micorrización de agaves con HMA pueden ser potenciales biofertilizantes y agentes de control biológico contra la enfermedad.
Es importante evidenciar los efectos positivos de la micorrización es bulbilos de agave, ya que los resultados que se presentan generalmente se realizaron en plántulas provenientes de rizoma y es necesario establecer si existen diferencias en los efectos dependiendo del origen del material vegetal y de la utilización de consorcios nativos provenientes de plantaciones de agave, por lo que en este trabajo se evaluó el efecto de los hongos micorrízicos arbusculares en el crecimiento y la bioprotección contra la marchitez causada por Fusarium oxysporum en bulbilos de agave tequilero bajo condiciones de invernadero. Se espera que la inoculación con consorcios de HMA nativos promueva el crecimiento de los bulbilos y disminuya o reduzca el daño ocasionado por Fusarium oxysporum.
Materiales y métodos
Material vegetal
Se utilizaron bulbilos florales de Agave tequilana Weber var. azul, recolectados de plantas adultas sanas de una parcela de agave de la zona de denominación de origen del tequila del municipio Guadalajara, Jalisco, México, de aproximadamente de tres meses de edad. Las plantas seleccionadas tuvieron condiciones morfológicas homogéneas y, en promedio, los bulbilos tuvieron una altura de 97 mm, un diámetro de piña de 22 mm, 4 hojas y peso promedio de 13 g; además, los bulbilos no tenían sistema radical, este lo comenzaron a desarrollar al momento del trasplante.
Hongos micorrízicos arbusculares
Se utilizaron dos fuentes de HMA, un consorcio micorrízico nativo de la zona mezcalera de Michoacán, BN, donde las esporas se obtuvieron de cultivos trampa en propagación con sorgo (Sorghum spp.). Este inóculo contenía esporas de Acaulospora denticulata, Acaulospora scrobiculata, Acaulospora spinosa, Claroideoglomus etunicatum, Diversispora aurantia y Glomus desertícola (Trinidad- Cruz, et al., 2017a), y el otro HMA fue un inoculo comercial (presentación en polvo) que contenía solo esporas de Rhizophagus intraradices (Ri), en ambos casos se inocularon 100 esporas por bulbilo y para inocular la cantidad de esporas requeridas se realizó el conteo de esporas del consorcio y del inóculo comercial, tomando 10 g de suelo seco para extraer las esporas mediante la técnica de tamizado húmedo, decantación (Gerdemann y Nicolson, 1963) y centrifugación con sacarosa al 50 % (Brundrett et al., 1996) por triplicado. Las esporas fueron contadas en un estereomicroscopio (VE-S6, Velab®) y se tomaron 15 g para inocular con 100 esporas de HMA por bulbilo de agave para el consorcio, 2,5 g para el inóculo comercial y para el testigo sin HMA se inoculó con 10 g de arena esterilizada, para que todos los bulbilos tuvieran el mismo manejo.
Obtención del inóculo de Fusarium oxysporum
La cepa de F. oxysporum fue proporcionada por el Laboratorio de Biotecnología Vegetal del Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco. Esta cepa fue aislada de plantas de A. tequilana con síntomas de marchitez (Qui-Zapata et al., 2011). Para obtener el inóculo de F. oxysporum, se partió de un cultivo de conservación de la cepa de la cual se realizó una transferencia de micelio y para su reactivación se llevó a cajas de Petri con medio de cultivo PDA estéril (121 °C, 1,05 kg cm-2, 20 min) (Trinidad-Cruz et al., 2017b). Estas se incubaron en oscuridad durante dos días a 27 °C ± 1 °C, después a 12/12 h de luz/oscuridad durante cinco días. Posteriormente, se recolectaron las esporas agregando 15 mL de agua desionizada estéril (20 min a 121 °C), a las cajas Petri y con un pincel esterilizado se hizo un barrido sobre la superficie; finalmente, se recuperó el agua destilada con las esporas con una micropipeta. La concentración de esporas se ajustó a 1 × 10. esporas mL-1 con un hematocitómetro y la suspensión de esporas se almacenó en tubos de 50 mL a 4 °C hasta su uso.
Obtención del sustrato infestado con Fusarium oxysporum
Para el sustrato con el inoculo de F. oxysporum se utilizó una mezcla de sustrato (arena, suelo, materia orgánica y vermiculita 30:50:5:15 v/v), con hojuelas molidas de avena y agua en proporciones 20:2:4 v/v, todo esterilizado (1 hora a 121 °C). Por cada 200 g de sustrato, se inocularon 500 µL de la suspensión de esporas diluidos en 9,5 mL de agua desionizada estéril (20 min a 121 °C), se incubó el sustrato inoculado con F. oxysporum en oscuridad a temperatura ambiente durante 20 días, mezclando el sustrato cada tercer día, se realizó un conteo de las unidades formadoras de colonias (UFC) por gramo de sustrato y se diluyó con sustrato estéril para tener una concentración de 1,5 × 10. propágulos g-1 de sustrato.
Micorrización del agave
Los bulbilos de agave previamente desinfectados (cloro al 3 % por 10 min y luego enjuagados tres veces con agua destilada por 5 minutos) se plantaron en macetas de plástico con capacidad de 250 g que contenían 200 g de una mezcla estéril (1 hora a 121 °C) de arena, suelo (suelo agrícola proveniente de una parcela experimental del Instituto de Investigaciones Agropecuarias y Forestales, Tarimbaro, Michoacán, México), materia orgánica (vermicomposta comercial) y vermiculita en proporciones 30:50:5:15, respectivamente. Al momento de plantar los bulbilos de A. tequilana, se colocaron 100 esporas viables de los inóculos utilizados en el agujero de siembra; para el caso del consorcio, se pesaron 15 gramos del medio donde se contenían los diferentes HMA que conformaban este (arena de río) y para el caso del inóculo monoespecífico, se pesaron 2,5 g y el tratamiento sin HMA fue inoculado con 5 gramos de arena estéril, de modo que todos los bulbilos tuvieran el mismo manejo. Una vez inoculados, los agaves se mantuvieron en condiciones de invernadero (temperatura y humedad relativa promedio de 26,4 °C ± 4 °C y 47 % ± 3 %, respectivamente) durante cuatro meses, con riegos a capacidad de campo con agua desionizada, cada vez que fuera necesario.
Determinación de crecimiento de agaves antes de la infección con F. oxysporum
Para evaluar la promoción del crecimiento de los bulbilos de A. tequilana, previo a la infección con Fusarium oxysporum, se utilizó un diseño completamente al azar donde se evaluaron tres tratamientos de HMA, el consorcio nativo BN, el inóculo comercial a base de Rhizofagus intrarádices (Ri) y un control sin micorriza. Se tomaron cinco y tres repeticiones para los tratamientos inoculados con HMA y sin HMA, respectivamente, donde cada planta representó una unidad experimental. A los cuatro meses después de la micorrización, se evaluó el número de hojas mediante el conteo de estas, el diámetro de la piña con un vernier digital, la altura de la planta con un flexómetro, la biomasa fresca total con una balanza analítica y la colonización de las raíces de agave con los HMA. Para esto se realizó primero el clareo y la tinción de raíces mediante la técnica de Phillips y Hayman (1970), se montaron segmentos de raíces en portaobjetos y se observaron en microscopio óptico las estructuras características de micorrización como esporas, vesículas, arbúsculos e hifas. Posteriormente, se cuantificó la colonización mediante el método de McGonigle et al. (1990).
Determinación del crecimiento de agaves infectados con Fusarium oxysporum y biocontrol por la micorrización
Los bulbilos de A. tequilana micorrizados durante cuatro meses fueron trasplantados con su cepellón a bolsas de polietileno negras de 2 kg de capacidad rellenas con el sustrato infestado con F. oxysporum; mientras el grupo de las plantas control (sin HMA) fue trasplantado al mismo sustrato pero sin el patógeno y todas las plantas se mantuvieron en condiciones de invernadero y se regaron dos veces por semana con agua desionizada. Después de 90 días (330 días después del establecimiento inicial), se realizó un muestreo destructivo donde se evaluó el número de hojas, el volumen de la raíz y su parte aérea (mediante la técnica de desplazamiento en agua), la biomasa fresca de raíz y su parte aérea (balanza analítica) y el grado de severidad ocasionado por F. oxysporum.
Para esto utilizó un diseño experimental factorial de 3 × 2 completamente al azar con diez repeticiones por tratamiento, donde se generaron seis tratamientos de la combinación de los tres niveles del factor micorriza (BN, Ri y sin HMA) y dos niveles del patógeno (con F. oxysporum y sin F. oxysporum); la unidad experimental fue una planta de A. tequilana por bolsa. Para la variable severidad, se generó una escala ordinal de severidad patogénica (figura 1) a partir de un experimento previo, donde se caracterizó el nivel y el avance del daño ocasionado por F. oxysporum en plantas jóvenes de Agave tequilana.
La escala contó con cinco niveles de enfermedad donde: el nivel 1 correspondió a la planta sana (asintomática); el nivel 2 muestra el inicio de la enfermedad (hojas con enrollamiento); en el nivel 3 se observa el avance medio de la enfermedad (hojas con enrollamiento, puntas necrosadas, clorosis); en el nivel 4 se observa un avance severo (la mayoría de las hojas con síntomas de marchitez, casi a la mitad de la hoja) y el nivel 5 correspondió a la planta muerta (hojas totalmente necrosadas y marchitas) (figura 1).

Análisis estadísticos
En ambos ensayos, el análisis de los datos cuantitativos fue mediante un análisis de varianza de una vía para el primer caso (antes de la inoculación con Fusarium oxysporum) y de dos para el segundo (después de la infección con Fusarium oxysporum), cuando el análisis mostró un efecto significativo (P < 0,05), se procedió a la prueba de comparación múltiple de medias con la prueba de Tukey (P < 0,05). Para los datos cualitativos, se realizó la prueba de Kruskal-Wallis e intervalos de confianza de la mediana, ambos con una P ≤ 0,05, donde todos los análisis se realizaron con el paquete estadístico SPSS.
Resultados y discusión
Efecto de los HMA como promotores de crecimiento previo a la infestación con Fusarium oxysporum
El análisis estadístico de las variables evaluadas a los 120 días de micorrizados los bulbilos mostró un efecto significativo (P < 0,05) de los diferentes HMA sobre las variables de crecimiento medidas, a excepción del diámetro de la piña (tabla 1).
Tabla 1. Efecto de los HMA sobre variables de crecimiento vegetal y la colonización micorrízica en plantas de agave bajo condiciones de invernadero a los 120 días después de la inoculación
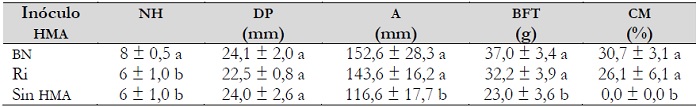
Notas aclaratorias:HMA: hongo micorrízico arbuscular; BN: Barranca de las Nueces; Ri: Rizophagus intraradices; A: altura; DP: diámetro de la piña; NH: número de hojas; BFT: biomasa fresca total y CM: colonización micorrízica; valores medios de N = 10 ± desviación estándar y diferentes letras en cada columna indican diferencias significativas (Tukey, P ≤ 0,05).
Fuente: Elaboración propia
Las plantas que se trataron con alguno de los inóculos micorrízicos aumentaron su tamaño significativamente respecto a las plantas sin inocular, siendo los bulbilos inoculados con el consorcio nativo BN los que obtuvieron el mayor crecimiento. En promedio, tuvieron dos hojas más y un incremento del 31 % y el 61 % en su altura y biomasa fresca total respecto a los bulbilos no micorrizados, respectivamente. Asimismo, al determinar la ganancia por el efecto micorrízico para las variables de crecimiento, se encontró que las plantas que fueron inoculadas con algún inóculo de HMA, en promedio incrementaron su altura en 30 mm (figura 2A). Para el número de hojas, los agaves micorrizados con el consorcio BN desarrollaron cuatro hojas más que los agaves con Ri y sin HMA que solo desarrollaron dos (figura 2B) y la ganancia en biomasa fresca de los agaves inoculados aumentó en promedio un 40 % respecto a los no micorrizados (figura 2C).
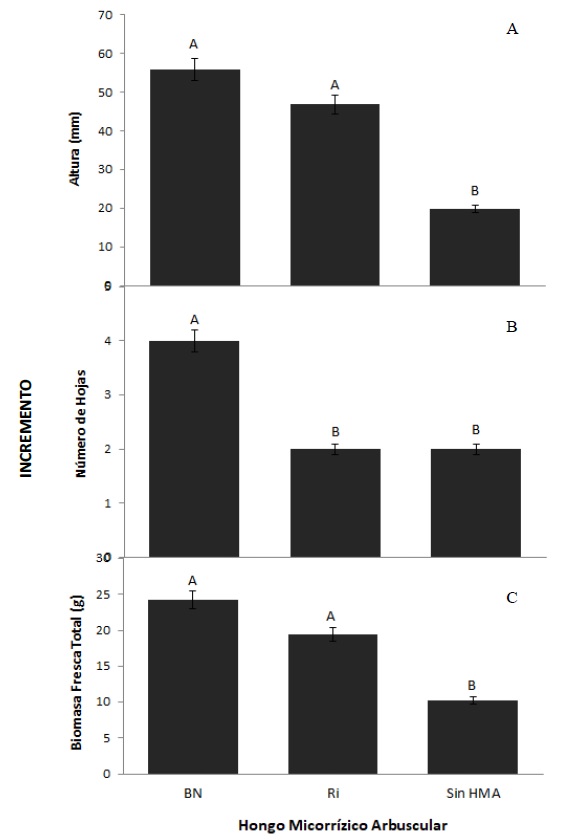
Fuente: Elaboración propia
Figura 2. Incremento en altura (A), número de hojas (B) y biomasa fresca total (C) por efecto de la inoculación con HMA en bulbilos de Agave tequilana a los 120 días de micorrizaciónNotas aclaratorias: HMA: hongo micorrízico arbuscular; BN: Barranca de las Nueces y Ri: Rizophagus intraradices; barras con letras distintas indican diferencia significativa (P < 0,05).
La respuesta en el crecimiento de plantas a la micorrización es muy diversa, algunos autores mencionan efectos positivos en variables morfológicas (Cozzolino et al., 2021), otros más reportan efectos negativos o nulos en este sentido (Smith & Smith, 2012). De manera particular en agaváceas, Pimienta-Barrios et al. (2009), evaluando Glomus fasciculatum y Glomus intraradices en plantas jóvenes de Agave tequilana, no encontraron diferencia en la longitud y el ancho de las hojas respecto a plantas sin micorrizar. Por su parte, Ruiz et al. (2011), en plantas in vitro de A. tequilana inoculadas con R. intraradices, mencionan que tuvieron los mismos valores en el número de hojas, la altura de planta y el peso fresco y seco, respecto a plantas sin micorrizar, evaluados a los 263 días después de la inoculación. Al respecto, Johnson et al. (1997) y Smith y Smith (2012) mencionan que, en algunos casos, el hongo puede estar actuando como un parásito, tomando el carbono de las plantas y aportando poco beneficio a las mismas, lo que se refleja en un crecimiento igual o menor a las plantas sin hongos micorrízicos. Asimismo, la selectividad hongo-planta puede estar influyendo en este caso de no promoción (Robles-Martínez et al., 2013; Quiñones-Aguilar et al., 2016).
En contraste, Robles-Martínez et al. (2013), trabajando con A. angustifolia micorrizados con diferentes hongos micorrízicos nativos, encontraron diferencias significativas en el peso seco del follaje respecto a plantas sin micorrización; Ruiz et al. (2011) reportan incrementos significativos en la biomasa fresca y seca de A. tequilanamicorrizados con Glomus intraradices; Santiz-Gómez et al. (2020) reportaron un incremento en la acumulación de azucares y un mejor desarrollo de A grivalvensis inoculado con G. fasciculatum; Montoya-Martínez et al. (2019) reportaron un incremento significativo en biomasa fresca, área foliar, número de hojas, entre otras variables en plantas de A tequilana inoculadas con consorcios nativos aislados de rizosfera de A cupreata y los mismos resultados fueron reportados por Quiñones-Aguilar et al. (2016) en A. inaequidens,Trinidad-Cruz et al. (2017b) en A. cupreata y García-Martínez et al. (2020) en agave tobalá (Agave potatorum Zucc) y agave coyote (Agave spp.), utilizando inóculos comerciales a base de esporas del género Glomus.
La promoción en el crecimiento se ha atribuido a varios procesos, como el incremento en la tasa fotosintética y una mayor absorción de agua y nutrientes debido a la actividad que realizan las hifas del hongo micorrízico al actuar como una extensión del sistema radicular de las plantas (Fernández-Lizarazo & Moreno-Fonseca, 2016; Vallejos-Torres et al., 2021), considerado este como el principal beneficio de los hongos micorrízicos (Quiñones-Aguilar et al., 2014). En nuestro caso, el incremento en las variables de crecimiento, evaluadas a los 120 días de inoculadas las plantas, pudo deberse al aumento en la fotosíntesis de las plantas y al beneficio de la mejora en la absorción de agua por parte de las hifas, ya que las raíces de las plantas presentaron una colonización del 28 %. Cui y Nobel (1992) reportaron una mayor tasa de absorción de CO2 en plantas de A deserti micorrizadas, lo cual se reflejó en plantas más grandes.
Con respecto a la colonización micorrízica, no se encontró diferencia significativa entre los agaves micorrizados con los diferentes HMA utilizados en este trabajo y en promedio se obtuvo una colonización del 28 %. Estos valores son relativamente bajos comparados con otras especies de agave, donde Robles-Martínez et al. (2013) reportan porcentajes de colonización del 35 % en plantas de Agave angustifolia, Zacarías-Toledo et al. (2016) del 73 % en A americana inoculados con G. fasciculatum y Santiz-Gómez et al. (2020) del 93 % en A. grivalvensis, inoculadas con Glomus Fasciculatum. En A. tequilana, Pimienta-Barrios et al. (2009) reportan porcentajes de colonización en hijuelos superiores al 70 % cuando fueron micorrizados con Glomus fasciculatum y mayores al 59 % cuando se micorrizaron con Glomus intraradices. En Agave inaequidens, Quiñones-Aguilar et al. (2016) reportan porcentajes de colonización del 20 al 45 % cuando fueron inoculados con consorcios nativos de HMA. Particularmente en Agave tequilana, Montoya-Martínez et al. (2019) reportaron porcentajes de colonización en promedio del 45 % cuando fueron inoculados con consorcios nativos de HMA aislados de rizosfera de Agave cupreata.
Los valores relativamente bajos de colonización en nuestro trabajo pueden ser debido, primeramente, al origen del material vegetal, pues se tienen reportes que en agaváceas dependiendo si es de hijuelos de rizoma o bulbilos (aéreos), dado que la colonización es distinta. En A. angustifolia se reporta que la colonización para el caso de hijuelos fue del 27,7 % y para bulbilos del 32,8 % (Robles-Martínez et al., 2013). Estos autores mencionan que la diferencia de colonización entre bulbilos e hijuelos no es del todo clara y, dependiendo de su origen, será la respuesta en el crecimiento cuando se encuentre micorrizada. En A. angustifolia, los hijuelos tienden a incrementar su biomasa y formación de nuevas hojas, mientras que los bulbilos aéreos tienden a incrementar su altura, como fue en nuestro caso.
Es necesario realizar trabajos donde se pueda corroborar lo anterior para el caso de A. tequilana y poder aclarar si la colonización de bulbilos aéreos es menor que en hijuelos rizomáticos. Otra razón en los bajos porcentajes de colonización podría ser por la selectividad planta-hongo o el tiempo en que se determinó la colonización, ya que al ser plantas de lento crecimiento, se necesitan tiempos más prolongados para alcanzar el máximo de colonización, sin embargo, los agaves micorrizados con el consorcio BN obtuvieron una mayor producción de hojas, esto en estas especies de lento crecimiento podría resultar una ventaja durante su desarrollo ya que al contar con una mayor superficie foliar, tendrán una mayor cantidad de producción de fotoasimilados, los cuales podrán ser utilizados posteriormente durante su crecimiento y que podría comprobarse parcialmente, de alguna manera, con la biomasa fresca total, que en el caso de estas plantas obtuvieron un 61 % más biomasa que los agaves sin micorrización.
Una posible causa de por qué las plantas inoculadas con BN incrementaron su biomasa es porque este inóculo está conformado por varias especies de hongos micorrízicos y se sabe que la promoción en el crecimiento de especies micorrizadas es mejor cuando se utilizan consorcios en lugar de inóculos no específicos, además que son mejores los consorcios cuando son de especies nativas o de rizosfera de especies de la misma familia o género de la especie a micorrizar (Quiñones-Aguilar et al., 2016; Chimal-Sánchez et al., 2018; Vallejos-Torres et al., 2021). La razón de esto puede deberse a la complementariedad funcional o el sinergismo de las diferentes especies de hongos micorrízicos presentes en los consorcios (García-Martínez et al., 2020).
Efecto de los HMA como promotores de crecimiento y biocontrol contra Fusarium oxysporum
A los 210 días de establecidas las plantas de agave, se realizó un análisis de varianza factorial, considerando como factores la micorrización y la inoculación al sustrato con Fusarium oxysporum. Los resultados del análisis de las variables evaluadas a esta fecha de muestreo mostraron solo efecto significativo para el factor de micorrización (P < 0,05), por lo que se procedió a la comparación de medias solo considerando este factor para el crecimiento de los agaves (tabla 2).
Tabla 2. Efecto de los hongos micorrízicos arbusculares sobre las variables de crecimiento vegetal en bulbilos de Agave tequilana, bajo condiciones de invernadero, a los 210 días después de la inoculación
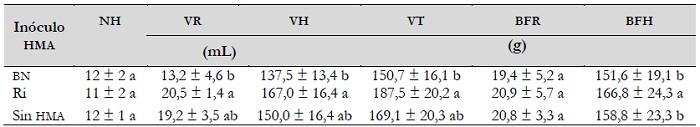
Notas aclaratorias: HMA: hongo micorrízico arbuscular; BN: Barranca de las Nueces; Ri: Rizophagus intraradices; NH: número de hojas; VR: volumen de raíz; VH: volumen de hoja; VT: volumen total; BFR: biomasa fresca de raíz; BFF: biomasa fresca foliar; Total de valores medios de N = 10 ± desviación estándar y diferentes letras en cada columna indican diferencias significativas según la prueba de Tukey (P ≤ 0,05).
Fuente: Elaboración propia
El análisis estadístico mostró efecto significativo para el volumen de raíz, hoja y total y la biomasa fresca de hoja. El mayor crecimiento se alcanzó en los agaves micorrizados con Ri, aunque para algunas variables no se obtuvieron diferencias significativas respecto a plantas sin micorrización o micorrizadas con el inóculo BN. Para las variables de número de hojas y biomasa fresca de raíz, en promedio los agaves tuvieron 12 hojas y su raíz pesó 20 gramos (tabla 2). Respecto a la biomasa fresca total, los agaves que fueron micorrizados con Ri, resultaron ser estadísticamente superiores a los agaves inoculados con el consorcio BN y sin micorrizar, que resultaron ser estadísticamente iguales entre sí (figura 3). En promedio, los agaves micorrizados con Ri incrementaron su biomasa un 12 % respecto a los demás tratamientos.
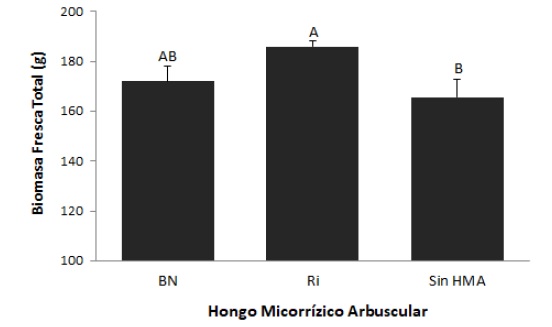
Fuente: Elaboración propia
Figura 3. Efecto de la inoculación con HMA en bulbilos de Agave tequilana a los 210 días después de la inoculaciónNotas aclaratorias: HMA: hongo micorrízico arbuscular; BN: Barranca de las Nueces y Ri: Rizophagus intraradices; las barras con letras distintas indican diferencia significativa (P < 0,05).
Se pudo observar que el efecto promotor inicial (a los 120 días de micorrización) que presentaron las plantas micorrizadas respecto a las no micorrizadas se fue perdiendo, pues en la mayoría de las variables evaluadas los valores de estas resultaron estadísticamente similares. Varias pueden ser las causas de este efecto, en primer lugar, el patógeno utilizado para este trabajo (F. oxysporum) ataca principalmente la raíz (Ávila-Miranda et al., 2010; Trinidad-Cruz et al., 2014; Ramírez- Ramírez et al., 2017), lo que podría ser que el ataque haya dañado el sistema radicular de los agaves y se disminuyera el beneficio de la micorrización en el crecimiento de la planta. Por otro lado, se ha planteado en algunos trabajos donde se inoculan plantas con patógenos y que esto implique un cambio de recipiente (maceta de mayor tamaño) que se haga una reinoculación con HMA para seguir favoreciendo la competencia entre microorganismos, además de que al momento de pasar la planta a un recipiente mayor se reduce el estrés mecánico, permitiendo una mayor penetración de agua (Ruiz et al., 2011), además de que haya una producción de raíces que requieran ser colonizadas y puedan explorar el nuevo volumen.
A esta fecha de muestreo, las plantas micorrizadas con el consorcio nativo BN resultaron tener un crecimiento estadísticamente similar a las plantas sin micorrizar. Esto podría sugerir que la infestación con Fusarium oxysporum pudo haber dañado el sistema radicular de estas plantas y verse reflejado en una disminución de nutrientes y agua, lo cual disminuiría el crecimiento y que parcialmente se puede explicar con los valores de volumen de raíz, el cual en estas plantas fue el más bajo, con un 36 %, respecto a las que en promedio obtuvieron las plantas micorrizadas con Ri y sin micorrizar.
La mayor ganancia de crecimiento en las plantas micorrizadas con Ri pudo deberse a la selectividad entre la especie de hongo específica de este inóculo y la planta. En trabajos con agave inoculado con HMA, es reportado un mayor crecimiento cuando fue inoculado con especies de Rhizophagus respeto a otras. En el inóculo Ri, la especie dominante son los HMA del género Rhizophagus y además se sabe que la colonización es distinta entre los diferentes géneros y especies de HMA.
Respecto al efecto bioprotector de la micorrización contra Fusarium oxysporum, el análisis estadístico realizado no mostró efecto en ninguno de los tratamientos evaluados. Todas las plantas inoculadas con el patógeno mostraron algún nivel de daño y las plantas micorrizadas, en general, presentaron hojas con enrollamiento, puntas necrosadas y pérdida del color verde en el 60 % de las hojas que, de acuerdo a la escala generada, presentaron un nivel tres de severidad; mientras que las plantas sin micorrizar presentaron la mayoría de las hojas con síntomas severos de marchitez, casi a la mitad de la hoja (80 %) que, de acuerdo a la escala generada, presentaron un nivel cuatro. El hecho de que no se observara un efecto bioprotector de los HMA ante el patógeno pudo deberse a varias circunstancias, por ejemplo, este patógeno afecta la raíz y no en el tejido foliar. Trinidad-Cruz et al. (2014) realizaron pruebas de patogenicidad con plantas de Agave cupreata micorrizadas e infestadas a 90 días con Fusarium oxysporum y reportaron que las plantas con y sin micorrización presentaron el mismo nivel de severidad foliar y que los agaves micorrizados mostraron un menor daño en sus raíces.
En este trabajo, solo se consideró el daño a las hojas, lo cual ante lo mencionado anteriormente podría suponerse que el control se manifestó en la raíz. En nuestro caso, el volumen radical de plantas micorrizadas con Ri fue similar a plantas sin inocular, sin embargo, el hecho de que estas plantas estuviera micorrizadas pudo ocasionar que tuvieran una mejor resistencia a la enfermedad, hecho que se pudiera reflejar en que estas plantas resultaron con la mayor ganancia en biomasa fresca. La efectividad del control de patógenos del género Fusarium en plantas usando HMA se ha reportado previamente, Jaiti et al. (2007) evaluaron la efectividad de los HMA contra la marchitez causada por F. oxysporumy la inducción de algunas reacciones bioquímicas relacionadas con la defensa de las plantas en palma datilera.
Estos autores reportaron que los HMA indujeron cambios en la actividad de enzimas relacionadas con la defensa de la planta cuando estas eran infectadas por el fitopatógeno. Kapoor (2008) realizó un experimento de bioprotección por HMA en tomate utilizando 10 mL de una suspensión de esporas de F. oxysporum f. sp. lycopersici, a una concentración de 2,8 × 10. esporas mL−1 y reportó una reducción de la enfermedad del 75 % en plantas inoculadas, respecto a plantas sin inoculación. Estos trabajos muestran que los HMA sí manifiestan un efecto de bioprotección contra patógenos como Fusarium oxysporum, aunque esto no se pudo encontrar totalmente en este trabajo.
El bajo porcentaje de colonización (30 %) en nuestro caso pudo haber favorecido mayores sitios de infección para el patógeno, además que los agaves pudieron tener una nutrición no adecuada, lo cual facilitaría el establecimiento del hongo y el desarrollo de la enfermedad. Se sabe que plantas poco nutridas limitan su crecimiento y son más susceptibles al ataque de plagas y enfermedades (Enríquez del Valle et al., 2018).
Conclusiones
Se observó un incremento en el crecimiento en los bulbilos de agave cuando se inocularon con HMA, previo a la infección con el patógeno Fusarium oxysporum. Además, no se encontraron diferencias significativas en las variables evaluadas en los agaves después de 100 días de estar infestados con Fusarium oxysporum. Respecto al daño ocasionado por Fusarium, el análisis estadístico no mostró diferencia significativa entre bulbilos de agave micorrizados y sin micorrizar, todos los bulbilos mostraron un nivel de daño, indicando que no se presentó un efecto de bioprotección contra el patógeno a nivel foliar; sin embargo, pudo identificarse un cierto control en agaves micorrizados, ya que las raíces de agaves micorrizadas con el inoculo Ri, presentaron un mayor crecimiento respecto al control, lo que podría indicar que el posible control puede ocurrir a nivel radicular. Es necesario continuar realizando investigaciones sobre el biocontrol de enfermedades utilizando recursos microbianos, pues, aunque en este trabajo estadísticamente no se encontró control contra la enfermedad en la sintomatología foliar, el control pudo estar a nivel radicular.
Descargos de responsabilidad
Todos los autores realizaron aportes significativos al documento, están de acuerdo con su publicación y manifiestan que no existen conflictos de interés en este estudio.
Contribución de los autores
Amelia Cristina Montoya-Martínez: registro de información en invernadero, elaboración de bases de datos, análisis de información y elaboración de manuscrito; Evangelina Esmeralda Quiñones-Aguilar: elaboración de diseño experimental, protocolos metodológicos, análisis de información, elaboración de proyecto para acceso a recursos económicos y elaboración de manuscrito; Gabriel Rincón-Enríquez: elaboración de diseño experimental, análisis de información, elaboración de proyecto para acceso a recursos económicos, protocolos metodológicos, y elaboración de manuscrito. Luis López-Pérez: elaboración de proyecto para acceso a recursos económicos, supervisión de actividades, análisis de información y elaboración de manuscrito.
Implicaciones éticas
Este artículo se ajustó a los códigos de ética de las instituciones educativas y de investigación en las cuáles están adscritos los colaboradores del mismo. Así mismo tuvo el consentimiento de los colaboradores para usar la información suministrada en la documentación del proceso presentado en el mismo.

