










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista de Medicina Veterinaria
versión impresa ISSN 0122-9354
Rev. Med. Vet. no.28 Bogotá jul./dic. 2014
Bioquímica sanguínea y concentración plasmática de corticosterona en pollo de engorde bajo estrés calórico
Elvis Alexander Díaz López1 / Luís Fernando Uribe Velásquez2 / William Narváez Solarte3
1 Médico veterinario zootecnista, Esp. MSc. Grupo de Investigación en Nutrición, Metabolismo y Seguridad Alimentaria, Departamento de Producción Agropecuaria, Universidad de Caldas, Manizales, Colombia.
elvis.diaz@ucaldas.edu.co
2 Médico veterinario zootecnista, MSc. PhD. Grupo de Investigación Cienvet, Departamento de Salud Animal, Universidad de Caldas, Manizales, Colombia.
lfuribe@ucaldas.edu.co
3 Zootecnista, Msc. PhD. Grupo de Investigación en Nutrición, Metabolismo y Seguridad Alimentaria, Departamento de Salud Animal, Universidad de Caldas, Manizales, Colombia.
wnarvaez@ucaldas.edu.co
Recibido: 29 de abril del 2014. Aceptado: 24 de junio del 2014
Cómo citar este artículo: Díaz López EA, Uribe Velásquez LF, Narváez Solarte W. Bioquímica sanguínea y concentración plasmática de corticosterona en pollo de engorde bajo estrés calórico. Rev Med Vet. 2014;(28):31-42.
Resumen
Las altas temperaturas ambientales propician susceptibilidad al estrés calórico en los pollos de engorde, lo que genera cambios metabólicos. Se buscó determinar los cambios en la bioquímica sanguínea y la concentración plasmática de corticosterona del pollo de engorde sometido a estrés calórico crónico y a las condiciones de temperatura ambiental del piedemonte amazónico colombiano, sobre las concentraciones plasmáticas de corticosterona, glucosa, proteínas totales, albumina, globulina, sodio, cloro, potasio, magnesio, fósforo y calcio. Se utilizaron pollos machos de 2 estirpes de 21 días de edad, distribuidos en un diseño experimental irrestrictamente al azar, en un esquema bifactorial, con 4 tratamientos. Se hicieron 5 repeticiones por tratamiento y 25 animales por unidad experimental. Las aves fueron alimentadas con una dieta básica de maíz y torta de soya con 3,100 Kcal de EM y 19,5% de proteína hasta los 42 días de edad. El factor estirpe no tuvo efecto en las variables evaluadas (p ≥ 0,05). No obstante, existió diferencia estadística significativa (p ≤ 0,05) en todas las variables evaluadas al ser comparadas las concentraciones de los metabolitos de las aves en condiciones de estrés calórico crónico con las de las aves expuestas a las condiciones de temperatura del piedemonte amazónico colombiano. Se concluyó que la bioquímica sanguínea sufrió modificaciones significativas bajo las 2 temperaturas experimentales, con mayor detrimento fisiológico de las aves con estrés calórico crónico. La concentración de corticosterona se constituyó en el indicador más sensible y constante de la condición fisiológica de estrés crónico por calor.
Palabras clave: avicultura, albúmina, estirpe, estrés, globulinas.
Blood Biochemistry and Plasma Corticosterone Concentration in Broiler Chickens Under Heat Stress
Abstract
High ambient temperatures cause susceptibility to heat stress in broiler chickens, generating metabolic changes. This paper seeks to determine the changes in blood biochemistry and plasma corticosterone concentration, as well as in glucose, total protein, albumin, globulin, sodium, chlorine, potassium, magnesium, phosphorus, and calcium in broiler chickens under chronic heat stress and at ambient temperature conditions at the Colombian Amazonian piedmont. 21-days-old male chickens of two lines were studied, distributed in an unrestricted random design, in a two-factor scheme, with four treatments. Five repetitions per treatment were performed, and 25 animals per experimental unit examined. Broilers were fed a basic diet of corn and soybean meal with 3,100 kcal ME and 19.5% protein until they reached 42 days of age. The line factor had no effect on the evaluated variables (p ≥ 0.05). However, there was statistically significant difference (p ≤ 0.05) in all variables when concentrations of metabolites in broilers under chronic heat stress were compared to those of chickens exposed to ambient temperatures at the Colombian Amazon piedmont. In conclusion, blood biochemistry suffered significant changes under both experimental temperatures, with more physiological detriment in broilers under chronic heat stress. Concentration of corticosterone became the most sensitive and consistent indicator of the physiological condition of chronic heat stress.
Keywords: poultry farming, albumin, line, stress, globulins.
Bioquímica sanguínea e concentração plasmática de corticosterona em frango de engorde bajo estresse calórico
Resumo
As altas temperaturas ambientais propiciam susceptibilidade ao estresse calórico nos frangos de engorde, o que gera mudanças metabólicas. Buscou-se determinar as mudanças na bioquímica sanguínea e a concentração plasmática de corticosterona do frango de engorde sometido a estresse calórico crónico e às condições de temperatura ambiental do sopé amazônico colombiano, sobre as concentrações plasmáticas de corticosterona, glucose, proteínas totais, albumina, globulina, sódio, cloro, potássio, magnésio, fósforo e cálcio. Utilizaram-se frangos machos de 2 estirpes de 21 dias de idade, distribuídos em um desenho experimental irrestritamente aleatório, em um esquema bivariado, com 4 tratamentos. Realizaram-se 5 repetições por tratamento e 25 animais por unidade experimental. As aves foram alimentadas com uma dieta básica de milho e torta de soja com 3,100 Kcal de EM e 19,5% de proteína até os 42 dias de idade. O fator estirpe não teve efeito nas variáveis avaliadas (p ≥ 0,05). Não obstante, existiu diferença estatística significativa (p ≤ 0,05) em todas as variáveis avaliadas ao ser comparadas as concentrações dos metabolitos das aves em condições de estresse calórico crônico com as das aves expostas às condições de temperatura do sopé amazônico colombiano. Concluiu-se que a bioquímica sanguínea sofreu modificações significativas sob as 2 temperaturas experimentais, com maior detrimento fisiológico das aves com estresse calórico crônico. A concentração de corticosterona se constituiu no indicador mais sensível e constante da condição fisiológica de estresse crônico por calor.
Palavras chave: avicultura, albumina, estirpe, estresse, globulinas.
INTRODUCCIÓN
Los ambientes calurosos afectan negativamente a los pollos de engorde de rápido crecimiento y ello interfiere con la disipación del calor producido internamente. Este hecho en ocasiones conduce a un incremento letal de la temperatura corporal (1,2). La temperatura ambiental alta es uno de los factores de mayor impacto negativo en el rendimiento del pollo de engorde, ya que causa disminución del consumo de alimento, menor ganancia de peso y aumento de la mortalidad, como resultado de intensas modificaciones hematológicas, bioquímicas, metabólicas y hormonales (3-5), originadas por la condición climática.
Cahaner y colaboradores (2) e Islam y Nishibori (6) afirman que existen diferencias en la tolerancia al estrés calórico entre las distintas estirpes de pollo, con lo cual indican que los linajes de rápido crecimiento, por su alta selección, son más sensibles a temperaturas ambientales altas.
Los electrolitos monovalentes como el sodio (Na+), el potasio (K+) y el cloro (Cl) ejercen funciones muy específicas en el equilibrio ácido-base. Estos electrolitos acompañados de minerales en su forma iónica como el calcio, el magnesio, el fósforo y las proteínas plasmáticas desempeñan un papel importante en la síntesis de proteínas de los tejidos, en el mantenimiento de la homeostasis intra y extracelular, en el mantenimiento del potencial eléctrico de las membranas celulares, en la regulación de la presión osmótica, así como en el funcionamiento del sistema nervioso (7). Sin embargo, el grado de impacto de las distintas formas de estrés calórico sobre los diferentes compuestos plasmáticos y sus efectos deletéreos sobre el pollo de engorde son todavía controversiales.
Colombia es un país con diversidad bioclimatológica por su ubicación en la zona tórrida. La radiación solar y la sensación térmica son relativamente constantes a lo largo del año, y su temperatura promedio aumenta en las épocas de verano. Entre estas zonas se encuentra el piedemonte amazónico, caracterizada por tener clima cálido, alta precipitación anual y alta humedad relativa, factores estos que aunados se convierten en limitantes para que el pollo de engorde moderno exprese a plenitud su potencial genético.
Bajo las condiciones climáticas del piedemonte amazónico colombiano, los resultados zootécnicos de la producción de pollo de engorde no son muy eficientes como resultado del efecto de las altas temperaturas, las que frecuentemente sobrepasan los límites de termorregulación de las aves. Debido a ello, también se presentan efectos fisiológicos perjudiciales en la salud de las aves.
El objetivo del presente estudio fue evaluar en dos estirpes modernas de pollo de engorde el efecto del estrés calórico crónico inducido y el estrés calórico bajo las condiciones ambientales del piedemonte amazónico colombiano, sobre la concentración plasmática de corticosterona y la bioquímica sanguínea, medida a través del ionograma plasmático, como una expresión fisiológica de las aves durante la fase comprendida entre los 21 y 42 días de edad.
MATERIALES Y MÉTODOS
Localización
El experimento se realizó en la granja Pasarela, situada en la zona rural del municipio de Florencia, Caquetá, Colombia, ubicada a 1°34'35.58''N y 75°32'39.68'' de longitud occidental y a 243 msnm con un clima cálido húmedo tropical. La evaluación de las condiciones ambientales se establecieron a través de la medición de la presión atmosférica utilizando un barómetro de tipo higrotérmico (PCE-THB 38, exactitud: 0,01 mmHg, España 2010). Se realizaron 1500 mediciones durante diferentes épocas del año (lluvia, transición y periodo seco), en seis lugares de la granja. Con los datos obtenidos se realizó un análisis estadístico descriptivo para determinar las condiciones climatológicas de la granja y su variabilidad (tabla 1).
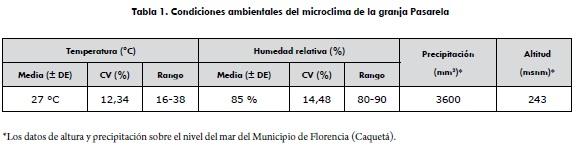
A los grupos de aves experimentales se les aplicaron prácticas de manejo convencionales, en cuanto a planes sanitarios, nutricionales y alimenticios requeridos para cada etapa productiva. Los pollos fueron criados durante las tres primeras semanas de vida, en un corral independiente de los módulos experimentales y sometidos a una temperatura inicial de 32°C ± 2°C y una humedad relativa de 66 ± 1,9%. La temperatura se redujo gradualmente hasta alcanzar una temperatura ambiente 27,07 ± 0,5°C y una humedad relativa de 78,2 ± 1,6% a los 21 días de edad, tiempo en el que se retiró la calefacción.
Animales
Fueron utilizados 250 pollos machos Cobb-500® y 250 pollos machos Ross 308®, de 21 días de edad. De cada línea genética se formaron grupos homogéneos con un peso promedio inicial de 773,56 ± 8,28 g, los cuales se alojaron en corrales de 3,8 m2, a una densidad de 6,58 aves/m2.
Etapa experimental
Se distribuyeron 125 pollos Cobb® y 125 pollos Ross® en un corral experimental, dentro de un galpón abierto en sus laterales sometidos a la temperatura ambiental en el piedemonte amazónico; y 125 pollos Cobb® y 125 pollos Ross® en un corral experimental dentro un galpón acondicionado con aislamiento plástico, madera comprimida y control ambiental (8) utilizando dos equipos acondicionadores de aire evaporativo (AS-T190206-Sumo GO®, USA, 2010), con rango de operación de temperatura (4-33 °C) y humedad relativa (0-95%). Las dos áreas experimentales fueron provistas de equipos comederos y bebederos.
Tratamientos
Los tratamientos aplicados desde los 21 hasta los 42 días de edad consistieron en evaluar al pollo de engorde macho de las dos estirpes comerciales en condiciones de estrés calórico crónico, manteniendo constante la temperatura a 30 ± 1 °C en un galpón con ambiente controlado y en un galpón abierto con la temperatura ambiental media del piedemonte amazónico (tabla 2). El registro de la temperatura ambiental y la humedad relativa de cada ambiente se realizó cada dos horas utilizando termohigrómetros digitales de máximas y mínimas (Kex Germany industries Inc. Modelo SH-121, con sensibilidad de 0,1°C y 0,1%, Rothenberg, Alemania, 2010), localizado a nivel de la altura de las aves durante las 24 horas del día del periodo experimental (tabla 3).
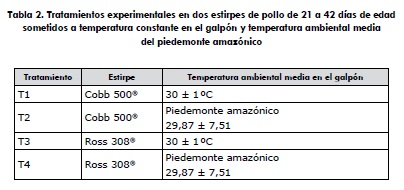
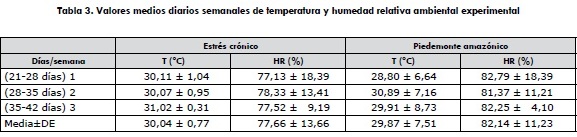
Los tratamientos se distribuyeron en un diseño irrestrictamente al azar (DIA), conformando un esquema factorial 2 x 2 (dos estirpes de pollo de engorde y dos condiciones de temperatura ambiental de cría) con cuatro tratamientos, cinco repeticiones por tratamiento y 25 animales por unidad experimental.
El programa de luz durante el periodo experimental para todos los tratamientos fue de 20 horas de luz día, complementando la luz natural con artificial incandescente, con una intensidad lumínica media de 10 ± 0,5 lux (9).
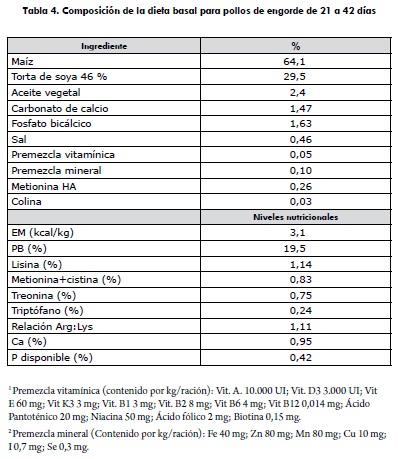
Las aves fueron alimentadas con una dieta básica de maíz, torta de soya y aceite vegetal con 3,1 Kcal de EM y 19,5% de proteína, según las exigencias nutricionales descritas por Rostagno y colaboradores (10) y NRC (11) para satisfacer los requerimientos nutricionales de las aves durante la fase de engorde (tabla 4). La dieta fue ofrecida en tipo migaja y junto con el agua fue consumida ad libitum por las aves durante todo el periodo experimental.
Muestreo
A los 42 días de edad de las aves en mismo periodo de tiempo y hora (6:00-7:00 a. m.) en todos los tratamientos, se obtuvieron muestras de sangre a través de punción de la vena braquial (12), previo ayuno de alimento durante 8 horas. El procedimiento de captura y sangrado no superó el minuto; por lo tanto, no debió influenciar la concentración de corticosterona plasmática (13,14).
Fueron extraídas cinco muestras, de 6 ml de sangre/ave, de cada unidad experimental para un total de 25 muestras por tratamiento para realizar la bioquímica sanguínea y establecer la concentración de corticosterona plasmática.
Para la obtención del plasma, a la sangre extraída se le adicionó ácido etilendiaminotetraacético (EDTA) o heparina (para la evaluación de iones plasmáticos) como anticoagulante, siendo centrifugada inmediatamente después de su extracción a 2800 rpm durante 15 min. En promedio se obtuvieron 2,4 mL de plasma por ave, los cuales fueron almacenados a -4°C hasta su evaluación. Las concentraciones plasmáticas de glucosa y corticosterona se estimaron en forma inmediata después de la obtención de las muestras. Los demás parámetros plasmáticos se evaluaron al siguiente día.
La corticosterona fue medida por la técnica de inmunoensayo directo, técnica fundamentada en el uso de anticuerpos marcados enzimáticamente para detectar la concentración de la hormona en las muestras de estudio, realizando la curva estándar para calcular la concentración de esta sobre la base de estándares. Para la prueba de Elisa se utilizó el kit marca Accubind®. La absorbancia se determinó con un espectrofotómetro lector de microplaca (Multiskan Plus-Fisher Scientific®) a 540 nm.
Las concentraciones plasmáticas de sodio, cloro, potasio, calcio, magnesio y fósforo se determinaron mediante métodos enzimáticos colorimétricos cuyas concentraciones se determinaron con un espectrofotómetro Spectronic® 20D, con una longitud de onda amplia de 340 a 950 nm, utilizando kits comerciales específicos para estos iones de la marca Spinreact® España.
Las concentraciones de albúmina, globulina y proteínas plasmáticas totales fueron medidas utilizando el método colorimétrico de Biuret. Se determinó la concentración de proteínas en las muestras de plasma mediante espectroscopia ultravioleta visible a una longitud de onda de 540 nm.
La determinación cuantitativa de la glucosa plasmática se determinó por el método enzimático colorimétrico glucosa oxidasa-peroxidasa, con lo cual se obtuvo la concentración de glucosa en plasma mediante espectroscopia a una longitud de onda de 505 nm.
Análisis estadístico
Los resultados de las variables evaluadas fueron sometidos a análisis de varianza, utilizando el procedimiento General Linear Models (GLM) de SAS-8.02 (2006). Las diferencias entre tratamientos fueron establecidas mediante la prueba de comparación múltiple de Tukey o la prueba de F a un nivel de confianza de p ≤ 0,05.
RESULTADOS Y DISCUSIÓN
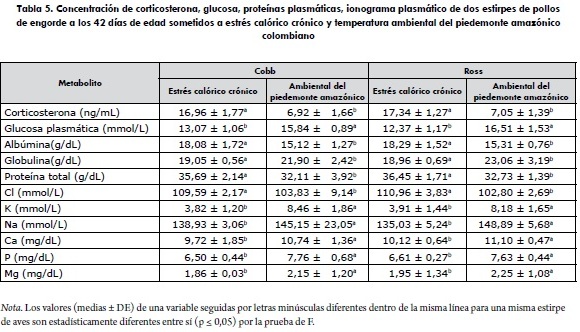
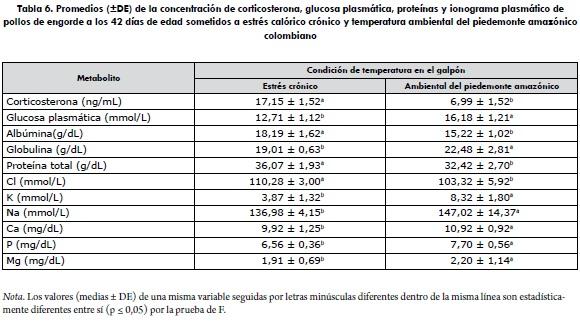
No existió efecto del factor estirpe (p ≥ 0,05) sobre ninguna de las variables de la bioquímica sanguínea y la concentración plasmática de corticosterona evaluadas en la investigación (tabla 5). Sin embargo, independientemente del linaje, las aves que estuvieron bajo el efecto del estrés calórico crónico presentaron estadísticamente (p ≤ 0,05) mayores concentraciones plasmáticas de corticosterona, albúmina, proteína total y cloro, e inferiores concentraciones de glucosa plasmática, globulinas, potasio, sodio, calcio, fósforo y magnesio que las aves criadas con la temperatura ambiental del piedemonte amazónico (tabla 6). Los resultados permiten inferir que, independientemente de la estirpe comercial de pollo de engorde, la respuesta fisiológica y hormonal de las aves sigue un comportamiento similar cuando se exponen a condiciones de estrés calórico crónico.
Corticosterona
Virden y colaboradores (15) después de someter pollos de engorde a estrés calórico crónico reportan concentraciones plasmáticas 18,5 ng/mL de corticosterona con una disminución en los niveles plasmáticos después del pico inicial. Al respecto, Post, Rebel y Terhuurne (16) y Shini y colaboradores (17) sostienen que las cantidades altas de corticosterona plasmática apuntan hacia una forma de estrés crónico, mientras que Mormede y colaboradores (18) consideran a esta hormona como el indicador fisiológico más importante para medir la respuesta del pollo de engorde al estrés.
Los máximos valores de corticosterona presentados por las aves con la condición estrés calórico crónico están por encima de las concentraciones normales fisiológicas de 2,49 a 6,40 ng/mL (16,17,19). Dichas concentraciones pueden ser ocasionadas por el efecto continuo de una alta temperatura, ya que según Puvadolpirod y Thaxton (19), este factor ambiental genera una respuesta hormonal de mayor frecuencia y duración, asociada con la adaptación y atenuación de la respuesta al estrés calórico; mientras que las aves expuestas a las condiciones de temperatura de estrés calórico en el piedemonte amazónico se encontraron cerca del límite normal superior, con un promedio, independientemente de la estirpe, de 6,99 ng/mL. Estos resultados se pueden relacionar con una exposición al estrés calórico agudo (20,21), el cual pudo provocar un aumento rápido y de corta duración en la concentración plasmática de este metabolito, con una posible tendencia de retorno a los valores normales, una vez que el factor estresor disminuye durante las horas diurnas y nocturnas más frescas del día.
Glucosa plasmática
Aunque estadísticamente las aves bajo las condiciones de estrés calórico crónico presentaron menor (p ≤ 0,05) concentración de glucosa que aquellas criadas bajo las condiciones de estrés del piedemonte amazónico, todos los valores circulantes de glucosa en el plasma sanguíneo encontrados en la investigación están dentro del intervalo de 11,1 a 22,2 mmol/L considerado fisiológicamente normal (22). La variabilidad en la concentración de glucosa plasmática, entre las aves expuestas a las diferentes condiciones de temperatura, se explica en función de los mecanismos homeostáticos eficientes del organismo, entre los que se involucra el control endocrino a través de la insulina y el glucagón sobre el glucógeno y los glucocorticoides en la gluconeogénesis (23). Revidatti y colaboradores (24) observaron valores de glucosa plasmática normales en pollos de engorde a los 42 días de edad después de ser sometidos a estrés. Puvadolpirod y Thaxton (19), después de inyectar pollos de engorde con hormona adrenocorticotrópica, concluyeron que el aumento de la concentración de corticosterona sanguínea conduce a una tendencia en la disminución de la glucosa plasmática, situación que se presenta bajo las condiciones de estrés calórico crónico, en el cual las concentraciones de glucocorticoides endógenos pueden aumentar, con subsecuente incremento de la concentración de glucosa plasmática durante el tiempo que permanezca el estímulo estresor (23,25).
Albúmina
La mayor concentración de globulina y menor concentración de albúmina y proteína total de las aves con estrés calórico crónico son aquellas bajo temperatura del piedemonte amazónico, probablemente se debe a la mayor producción de corticosterona plasmática de las aves con ese tipo de estrés, y a los efectos que posee esta hormona sobre el metabolismo nitrogenado (24). Es importante destacar que los valores de proteínas plasmática halladas en este trabajo se encuentran dentro de la media referencial normal de proteína total (29,7-35,9 g/dL), albúmina (15,1-16,1 g/dL) y globulina (15,6-21,6 g/dL) en pollos de engorde de 42 días de edad (26).
Los resultados obtenidos en esta investigación permiten identificar una manifestación de repuesta probablemente encaminada a un acondicionamiento de las dos estirpes de aves frente a las condiciones ambientales experimentales establecidas (27).
Ionograma plasmático
Los resultados de la concentración de electrolitos corroboran los reportes de Belay y Teeter (28), Borges (29) y Mushlag y colaboradores (30), y se pueden interpretar como parte de la respuesta funcional a través de los mecanismos homeostáticos, dirigidos a contrarrestar la alcalosis respiratoria generada durante el estrés calórico crónico.
Sodio
Después de comparar los resultados de la natremia con los parámetros normales fisiológicos de 140,50 a 149,50 mmol/L, para pollos de engorde a los 42 días de edad (30), se evidencia una disminución en la concentración de este ion para las dos estirpes de aves expuestas a estrés calórico crónico, en la que se observa una diferencia inferior de 4,2% para el Cobb y 6,8% para el Ross. Este hallazgo es muestra de la alteración del equilibrio ácido-básico con una probable variación en la presión osmótica, como resultado de una baja retención de sodio en los riñones, favorecido —ocasionalmente— por un aumento en el consumo de agua, en el que el gradiente osmótico pudo facilitar el movimiento del agua del fluido intracelular hacia la orina, transportando posiblemente más Na+, hecho que incidiría en la presentación de una hiponatremia (28). En este fenómeno fisiológico, conocido como hemodilución (31), los tejidos empiezan a liberar K+ hacia el torrente sanguíneo, debido a una alteración en la permeabilidad de las membranas, predisponiendo al animal a la presentación de signos neurológicos.
Cloro
El aumento de la cloremia observado en las aves expuestas al estrés calórico crónico se presume como consecuencia del funcionamiento de un sistema tampón, en el cual el Cl deprime la excreción de H+ y la reabsorción de HCO3 por los riñones, lo que contribuye a la acidificación de la sangre como posible respuesta a la alcalosis (29).
Potasio
La disminución de la calemia en las aves en condiciones de estrés calórico crónico se puede atribuir a un aumento en la excreción de este ion durante el estrés calórico, con la consecuente reducción en la retención (29,32,33). El incremento en la excreción de K+ a través de la orina es variable, y está ligada a la concentración plasmática de Na+ y al estado de hidratación del ave. Las pérdidas también pueden ser causadas probablemente por aumento en el consumo de agua, ya que el gradiente osmótico favorece el movimiento de agua del fluido celular para la formación de orina (28).
Calcio y magnesio
Todos los resultados de la calcemia y magnesemia determinados se encuentran dentro de los intervalos paramétricos fisiológicos normales de 8,9 a 12,0 mg/dL y 1,82 a 2,78 mg/dL, respectivamente, establecidos para pollos de engorde a los 42 días de edad (34). Los valores inferiores de la calcemia, fosfatemia y magnesemia de los pollos de engorde en las condiciones de estrés calórico crónico posiblemente sean consecuencia de la disminución en el consumo de alimento ocasionado por la condición ambiental (35) y como respuesta del ajuste homeostático realizado por el organismo animal en las condiciones climáticas estresoras.
Fósforo
Al contrastar los resultados experimentales con la concentración plasmática normal de fósforo que está entre 7,2 y 7,81 mg/dL en pollos de engorde a los 42 días de edad (34), se evidencia disminución en los índices presentados por las dos estirpes de aves sometidas a estrés calórico crónico; mientras que aquellas aves en las condiciones de temperatura del piedemonte amazónico se mantuvieron dentro de los parámetros normales. Este efecto puede estar relacionado con un incremento en la excreción del fósforo y, al mismo tiempo, disminución en la absorción intestinal y renal (35). Al respecto, Lumei (36) sostiene que en cuadros de deshidratación asociados con poliuria o pasaje rápido del alimento en estrés calórico crónico puede ocurrir un incremento en la excreción de algunos metabolitos, entre ellos el fósforo.
El estrés calórico cíclico generado en los animales por las condiciones climáticas ambientales del piedemonte amazónico de esta zona puede haber formado alteraciones en la excreción y absorción de fósforo en riñones e intestinos, variando la fosfatemia en los momentos posteriores a la presentación del estrés agudo, regresando luego a su valor normal en un tiempo variable. Este es un fenómeno biológico que según Puvadolpirod y Thaxton (19) puede ocurrir dependiendo de la duración y la magnitud del estímulo estresor inicial.
CONCLUSIONES
Las concentraciones de corticosterona, glucosa plasmática y el ionograma plasmático del pollo de engorde, independientemente de la estirpe, en las condiciones de estrés calórico, sufren modificaciones como respuesta del organismo animal a mantener su homeostasis. Sin embargo, el efecto es más pronunciado cuando los animales se exponen a condiciones de estrés calórico crónico que cuando se someten a esta condición climática en ambientes como el del piedemonte amazónico, donde el comportamiento cíclico diario de la temperatura tiene algunos periodos de temperaturas menos calurosos, permitiéndole al animal una recuperación homeostática.
Las dos líneas de pollo evaluadas a la edad de 42 días, expuestas a estrés calórico crónico a partir de los 21 días de edad, evidenciaron aumento en la concentración de corticosterona plasmática, hecho que permite ratificar esta hormona como un indicador fisiológico de respuesta del organismo animal a la condición de estrés calórico crónico.
REFERENCIAS
1. Cahaner A, Deeb N. Breeding broilers for adaptability to hot conditions. Procedente del XXII World Poultry Congress;2004 jun. 8-13; Estambul (Turquía). (CD-ROM) [ Links ]
2. Cahaner A, Ajuh JA, Siegmund Schultze M, Azoulay Y, Druyan S, Zárate AV. Effect of the genetically reduced feather coverage in naked neck and featherless broilers on their performance under hot conditions. Poult Sci. 2008;87(12):2517-27. [ Links ]
3. Gyimothy, I. Stress factors and stress responses in poultry breeding Literature review. Magy Allatorvosok Lapja. 2004;126(2):101-6. [ Links ]
4. Mushtaq T, Sarwar M, Nawaz H, Mirza MA, Ahmad T. Effect and interactions of sodium and chloride on broiler starter performance (one to twenty-eight days) under subtropical summer condition. Poult Sci. 2005;84(11):1716-22. [ Links ]
5. Ahmad T, Sarwar M. Dietary electrolyte balance: Implications in heat stressed broilers. Worlds Poult Sci J. 2006;62(4):638-53. [ Links ]
6. Islam MA, Nishibori M. Indigenous naked neck chicken: a valuable genetic resource for Bangladesh. World Poult Sci J. 2009;65(1):125-38. [ Links ]
7. Olanrewaju HA, Wongpichet S, Thaxton JP. Dozier WA 3rd, Branton SL. Stress and acid-base balance in chickens. Poult Sci. 2006;85(7) : 1266-74. [ Links ]
8. Estrada Pareja MM, Márquez Girón SM, Restrepo Betancur LF. Efecto de la temperatura y la humedad relativa en los parámetros productivos y la transferencia de calor en pollos de engorde. Rev Col Cienc Pec. 2007;20(3):288-303. [ Links ]
9. Schwean-Lardner K, Classen H. Iluminación para pollo de engorde. Aviagen TM [internet]; 2010 [citado 2014 mar. 31]. Disponible en: file:///C:/Users/Admin/Downloads/LightingforBroilers2010-ES.1.pdf [ Links ]
10. Rostagno HS, Albino LFT, Donzele JL, Gomes PC, Oliveira RF, Lopes DC, et al. Tabelas brasileiras para aves e suínos: composição de alimentos e exigências nutricionais. 2a. ed. Viçosa (Brasil): UFV Departamento de Zootecnia; 2005. [ Links ]
11. National Research Council (NRC). Nutrient requirements of poultry. 9 Rev Ed National Academy Press. Washington, DC; 1994: 155. [ Links ]
12. Samour J. Clinical and diagnostic procedures. 3a. ed. Livingston New York: Avian Medicine; 2000: 178. [ Links ]
13. Lagadic H, Faure JM, Mills AD, Williams JB. Effects of blood sampling on plasma concentrations of corticosterone and glucose in laying hens caged in groups. Br Poult Sci. 1990;31(4):823-9. [ Links ]
14. Romero LM, Reed JM. Collecting baseline corticosterone samples in the field: Is under 3 min good enough? Comp Biochem Physiol A Mol Integr Physiol. 2005;140(1):73-9. [ Links ]
15. Virden WS, Thaxton JP, Corzo A, Dozier WA 3rd, Kidd MT. Evaluation of models using corticosterone and adrenocorticotropin to induce conditions mimicking physiological stress in commercial broilers. Poult Sci. 2007;86(12):2485-91. [ Links ]
16. Post J, Rebel MJ, Terhuurne AA. Physiological effect of elevated plasma corticosterone concentration in broiler chickens. An alternative means by which to assess the physiological effect of stress. Poult Sci. 2003;82(8):1313-8. [ Links ]
17. Shini S, Kaiser P, Shini A, Bryden WL. Biological response of chickens (Gallus gallusdomesticus) induced by corticosterone and a bacterial endotoxin. Comp Biochem Physiol B. 2008;149(2):324-33. [ Links ]
18. Mormede P, Andanson S, Auperin B, Beerda B, Guemene D, Malmkvist J, et al. Exploration of the hypothalamic-pituitary-adrenal functions as a tool to evaluate animal welfare. Physiol Behav. 2007;92(3):317-39. [ Links ]
19. Puvadolpirod S, Thaxton JP. Model of physiological stress in chickens 1. Response parameters. Poult Sci. 2000;79(3):363-9. [ Links ]
20. Yuanianto VD, Hayashi K, Kaneda S, Ohtsuka A, Tomita Y. Effect of environmental temperature on muscle protein turnover and production in tube-fed broiler chickens. Br J Nutr.1997; 77(6): 897-909. [ Links ]
21. Cockrem JF. Stress, corticosterone response and avian personalities. J Ornithol. 2007;148(2):169-78. [ Links ]
22. Noriega MLVC. Importância da hematologia no diagnóstico das aves. Procedente de IV Encontro Técnico Sobre Avicultura de Corte da Região de Descalvado; 2000; Descalvado (Sao Paulo). p. 1-11. [ Links ]
23. Harr KE. Clinical chemistry of companion avian species: A review. Vet Clinic Pathol. 2002;31(3):140-51. [ Links ]
24. Revidatti FA, Fernandez RJ, Terraes JC, Sandoval GL, Luchi PE. Modificaciones del peso corporal e indicadores de estrés en pollos parrilleros sometidos a inmovilización y volteo. Rev Vet Arg. 2002;12(1):1-8. [ Links ]
25. Carsia, RV, Harvey S. Adrenals. En: Whittow GC, editor. Sturkie's Avian Physiology. 5a ed. San Diego, CA: Academic Press; 2000. [ Links ]
26. González FHD, Haida KS, Mahl D, Giannesi G, Kronbauers E. Incidência de doenças metabólicas em frangos de corte no sul do Brasil e uso do perfil bioquímico sanguíneo para o seu estudo. Ciênc Avic. 2001;3(2):141-7. [ Links ]
27. González FHD, Silva SC. Introdução a Bioquímica Clínica Veterinária. Porto Alegre, Brasil: Universidade Federal do Rio Grande do Sul; 2003. [ Links ]
28. Belay T, Teeter RG. Broiler water balance and thermobalance during thermo neutral and high ambient temperature exposure. Poult Sci. 1993;72(1):116-24. [ Links ]
29. Borges SA. Suplementação de cloreto de potássio e bicarbonato de sódio para frangos de corte durante o verão. Jaboticabal [Dissertação Mestrado]. São Paulo: Universidade Estadual Paulista; 1997. [ Links ]
30. Mushlag J, Mirza MA, Athor M, Hooge DM, Ahmad T, Ahmad G, et al. Dietary sodium and chloride for 29 to 42 day old broilers at constant electrolyte balance under subtropical conditions. J Appl Poult Res. 2007;16(2):161-70. [ Links ]
31. Borges AS, Maiorka A, Silva AVF. Fisiologia do estresse calórico e a utilização de eletrólitos em frangos de corte. Ciênc Rural. 2003;33(5):975-81. [ Links ]
32. Smith MO, Teeter RG. Potassium balance of the 5 to 8-week-old broiler exposed to constant heat of cycling high temperature stress and the effects of supplemental potassium chloride on body weight gain and feed efficiency. Poult Sci. 1987;66(3):487-92. [ Links ]
33. Salvador D, Ariki J, Borges AS, Pedroso AA, Barbosa Moraes VM. Suplementação de bicarbonato de sódio na ração e na água de bebida de frangos de corte submetidos ao estresse calórico. ARS Veterinária. 1999;15(2):144-8. [ Links ]
34. Kaneko JJ, Harvey JW, Bruss ML. Clinical biochemistry of domestic animals. Nueva York: Academic Press; 1989. [ Links ]
35. Klasing, K. Comparative avian nutrition. Londres: Cab International; 1998. [ Links ]
36. Lumei JT. Avian clinical biochemistry. En: Kaneko JJ, Harvey JW, Bruss ML, editores. Clinical biochemistry of domestic animals. 5a ed. San Diego, CA: Academic Press; 1997. p. 857-79. [ Links ]
