










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.21 no.1 Santa Marta Jan./Dec. 1992
MACROINVERTEBRADOS COLONIZADORES DE RAÍCES DE RHIZOPHORA MANGLE EN LA BAHÍA DE CHENGUE, CARIBE COLOMBIANO
Rosalba Reyes1 y Néstor Hernando Campos2
1Carrera 17 N° 11 -44, Santa Marta, Colombia.
2Prof. ICN, Univ. Nal. Colombia, c/o INVEMAR, A. A. 1016, Santa Marta, Colombia.
RESUMEN
Con el fin de estudiar la colonización de macroinvertebrados en las raíces de Rhizophora mangle, se realizó entre marzo y diciembre de 1989 un experimento a corto término, in situ en la Bahía de Chengue, usando como sustrato estacas de mangle y se comparó la composición de macroinvertebrados de éstas con la fauna asociada a las raíces de mangle. La mayoría de las especies colonizadoras formaron parte de la fauna asociada a las raíces; en total colonizaron 80 entidades conformadas por 35 crustáceos, 15 moluscos, 11 anélidos, 10 cnidarios, 4 briozoos, 3 esponjas y 2 cordados; se cuantificaron 2002 Individuos, la mayoría crustáceos (1757). La colonización de las estacas se vio favorecida en los meses lluviosos, época en la que se midieron los valores más altos de diversidad en las dos estaciones. La vecindad de otros ecosistemas es un factor determinante en el arribo de organismos, hecho que se evidenció en la similaridad de la fauna asociada a las raíces de mangle, con la fauna colonizadora de las estacas en la estación ubicada en la laguna y la menor afinidad con la fauna adherida a las estacas de la estación expuesta directamente al mar.
ABSTRACT
In order to study the colonization of macroinvertebrates on the Rhizophora mangle roots, a short term in situ experimet was conducted between March and December 1989, at Bahia de Chengue, using mangrove stick as substrate and comparing their macroinvertebrate composition to the fauna associated to the mangrove roots. Most of the colonizing species were also associated to the roots; 80 species as a total appeared as colonizers: 35 crustaceans, 15 molluscs, 11 annelids, 10 cnidarians, 4 briozoan, 3 sponges and 2 chordates; 2002 specimens were counted, most of them (1757) crustaceans. The rainy season seems to favor colonization, since the highest values of diversity were measured then. Closeness to other habitats is a determinant factor for the arrive of organisms; this is shown by the similarity between the fauna associated to the mangrove roots and that colonizing the sticks in the lagoon, and the lower affinity between the roots's fauna and that colonizing the sticks directly exposed to the sea.
INTRODUCCIÓN
El manglar es una unidad ecológica de características muy peculiares y exclusiva de las costas y estuarios tropicales; su distribución geográfica está estrechamente ceñida a la faja climática comprendida entre los trópicos de Cáncer y de Capricornio (Pannier y Pannier, 1976). En la región del Mar Caribe se encuentran Rhizophora mangle Linnaeus, 1753, Avicennia germinans (Linnaeus, 1753), Laguncularia racemosa (Gaetner, 1805), Conocarpus erectus Linnaeus, 1753 y Pelliciera rhizophorae Triana y Planchón, 1862 (Cintron y Schaeffer-Novelli, 1983; Calderón-Sáenz, 1983).
La mayoría de los trabajos realizados sobre las comunidades de las raíces del mangle se caracterizan por resaltar la necesidad de la disponibilidad de las raíces como sustrato duro para una gran cantidad de biota. Esta disponibilidad es restringida, ocasionando una competencia espacial entre los organismos sésiles; la escasez de sustrato puede ser superada por ciertas especies a través de adaptaciones morfológicas y químicas especializadas. Esta situación se puede apreciar en las raíces de mangle, donde pequeñas esponjas, hidroides y briozoos crecen sobre colonizadores primarios (Rutzler, 1969).
Entre los estudios experimentales sobre colonización se destaca el de Sutherland (1980), quien utilizó láminas de asbesto como sustrato para estudiar la dinámica de la comunidad epibéntica de las raíces del mangle rojo en la Bahía de Buche en Venezuela. En Colombia tan sólo se destaca un ensayo realizado por Palacio (1983) sobre colonización en la Ciénaga Grande de Santa Marta.
Con el fin de estudiar los cambios que se presentan a lo largo del año en la composición de los macroinvertebrados colonizantes en las raíces de R. mangle, se realizó un experimento a corto término in situ, usando como sustrato estacas de mangle.
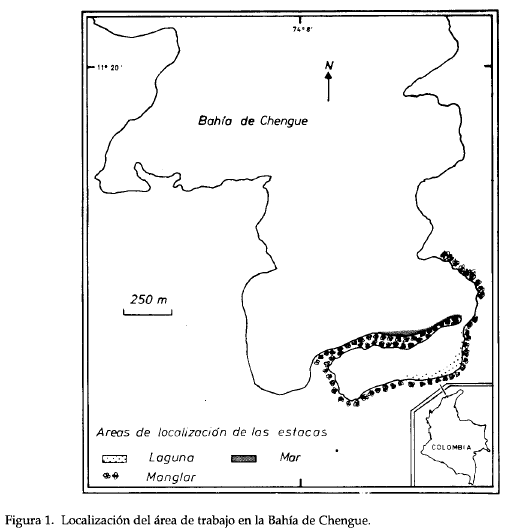
Este estudio se realizó en la Bahía de Chengue, Parque Nacional Natural Tayrona (PNNT), la cuál presenta orientación sur-norte y cubre una superficie aproximada de 2.5 Km2. En las partes sur y sureste de la bahía se encuentran formaciones coralinas que crecen delante de fondos someros poblados por Thalassia testudinum. La acumulación de sedimento en el sur propició la formación de una laguna costera bordeada por R. mangle y esporádicamente A. germinans (ob. pers.).
MATERIALES Y MÉTODOS
Se seleccionaron dos estaciones: una en la parte interna de la laguna (LAG) y la otra en la franja externa del manglar, en la parte expuesta directamente al mar (MAR) (Fig. 1). Se emplearon como sustrato estacas de mangle de aproximadamente 120 cm de longitud y 7 cm de diámetro; se colocaron diez estacas a lo largo del borde externo del mangar en cada estación. Se realizaron muestreos en marzo, junio, septiembre y diciembre de 1989, en cada uno se recolectaron cinco estacas por estación, y se reemplazaron por otras diez, para cubrir posibles pérdidas. Las raíces se colectaron cubriéndola con una red de ojo de malla de 5 mm y se cortó con un serrucho por encima de la marca del nivel de agua. Las estacas y las raíces se empacaron en bolsas de polietileno debidamente rotuladas y se transportaron al laboratorio, donde se congelaron.
De las raíces de mangle y de las estacas se retiraron manualmente los organismos, se separaron por taxa y se preservaron en alcohol al 70% en frascos codificados. La determinación espeífica se realizó con base en claves taxonómicas y con la ayuda de especialistas en los diferentes grupos de organismos. Una vez identificados se realizó el conteo de individuos de las especies sésiles-solitarias y vágiles. No fue posible cuantificar la fauna colonial debido a la superposición de las colonias, y algunas especies de moluscos y crustáceos por la dificultad para tomar todos los individuos capturados; de estos organismos sólo se registró presencia-ausencia.
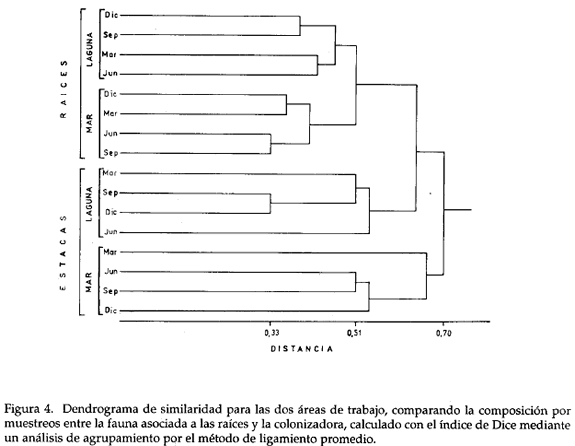
La caracterización de las comunidades de macroinvertebrados colonizadores se hizo mediante el índice de diversidad de Shannon y Weaver (1963) a partir del número de individuos por especie por muestreo en las estacas. Para la comparación de las comunidades se utilizó el índice de similaridad de Dice (Boesch, 1977; Digby y Kempton, 1987) a partir de datos binarios (presencia-ausencia). El índice de Dice se empleó también para comparar la composición faunística de las raíces de mangle con la de las estacas. Se realizó un análisis de agrupamiento por el método de ligamiento por promedio de grupo a partir de las matrices de asociación y se construyeron los respectivos dendrogramas.
RESULTADOS
El componente faunístico para este estudio de colonización provino de la recolección de 40 estacas de mangle. En la tabla 1 se da la lista y el número de individuos de las especies cuantificables y en la tabla 2 se da la lista completa de las especies colonizadoras.
El número total de especies colonizadoras de las dos estaciones difirió tan sólo en una, siendo 52 en MAR y 53 en LAG. En el muestreo de septiembre se reclutó el mayor número de especies en las dos estaciones (38 en LAG y 32 en MAR) y en junio el menor número (20 en LAG y 15 MAR). Durante todos los muestreos la estación LAG presentó mayor número de especies que la estación MAR (Fig. 2a).
De los 2002 individuos colonizadores de las estacas durante el año de estudio, se cuantificaron 421 en LAG y 1581 en MAR. La mayor fluctuación en el número de individuos se presentó en MAR, con 31 individuos en junio y 1306 en marzo (Fig. 2b). Las 80 especies de macroinvertebrados se distribuyeron por taxones así: tres esponjas, diez cnidarios, cuatro briozoos, 15 moluscos, once anélidos, 35 crustáceos y dos cordados (Fig. 2c).
En cuanto al número de individuos, el taxon mejor representado fue el de los crustáceos (1757), encontrados en su mayoría en las estacas de la estación MAR (1467). De los 132 individuos de anéli dos, el mayor número (105) se halló en LAG y el menor (27) en MAR. El mayor número de individuos de moluscos se halló MAR (87) y el menor en LAG (26) (Fig. 2d).
El elevado número de formas colonizadoras de estacas estuvo representado en general, por los mismos taxa que se hallaron en las raíces, a excepción de sipuncúlidos y entoproctos (tabla 1). Entre los organismos coloniales sésiles, que se pueden considerar pioneros, por la abundancia en que se hallaron en las estacas, se destacaron los hidroides, los briozoos y las ascidias.
En la tabla 2 se comparan las especies capturadas en las raíces y en las estacas en las dos estaciones de la Bahía de Chengue (presencia-ausencia). Del total de especies colectadas en las raíces y en las estacas (189), 80 se encontraron en las estacas y 179 en las raíces. La ausencia de 10 especies en las raíces parece indicar que no son típicas del manglar, como la esponja Callyspongia pallida, los moluscos Batillaria minima, Gibberula lavalleeana, Pilsbryspira hardfordiana y Crucibulum striatum. Estas especies no han sido registradas en otros trabajos como fauna asociada a raíces de mangle, además de tres especies de anfípodos no identificados, poslarvas de camarones peneidos y estado Glaucothoe de cangrejos no identificados.
Otras especies colonizaron las estacas en una sola época del año y se caracterizaron por presentar poblaciones muy abundantes. En las estacas colocadas entre marzo y diciembre colonizaron los bivalvos Crassostrea rhizophorae y Lopha frons; los cirripedios Balanns eburneus, B. amphitrite y B. improvisus y en las estacas colocadas entre junio y septiembre el isópodo Limnoria quadripunctata (tabla 1).
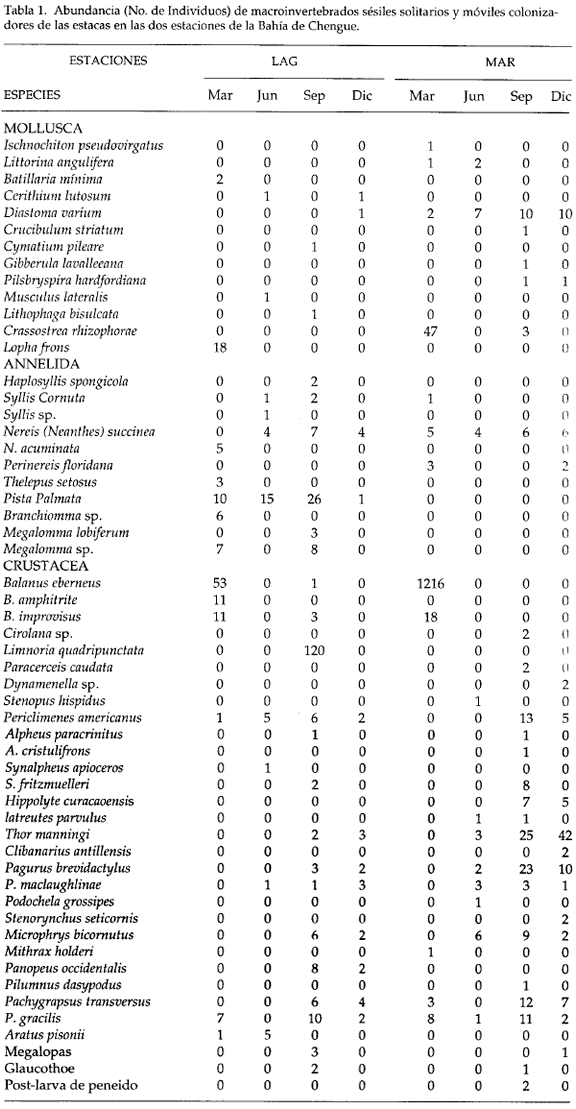
Se observó que de 15 especies de esponjas presentes en las raíces de la Bahía de Chengue, tres colonizaron las estacas; de éstas, Dysidea etheria y Haliclona hogarthi se hallaron tanto en las raíces como en las estacas.
De las 15 especies de cnidarios, diez hidroides colonizaron las estacas; entre éstas se destacaron por colonizar en las dos estaciones un atecado sin identificar, Halecium halecinum, Clytia gravieri, Dynamena crisioides y Plumularia halecioides.
De las seis especies de briozoos presentes en las raíces cuatro colonizaron las estacas y únicamente Amathia distans se encontró en las dos estaciones.
Los moluscos estuvieron representados por 43 especies. Entre las 15 que colonizaron las estacas, cuatro no se hallaron en las raíces y solo Diastoma varium se encontró en las dos estaciones.
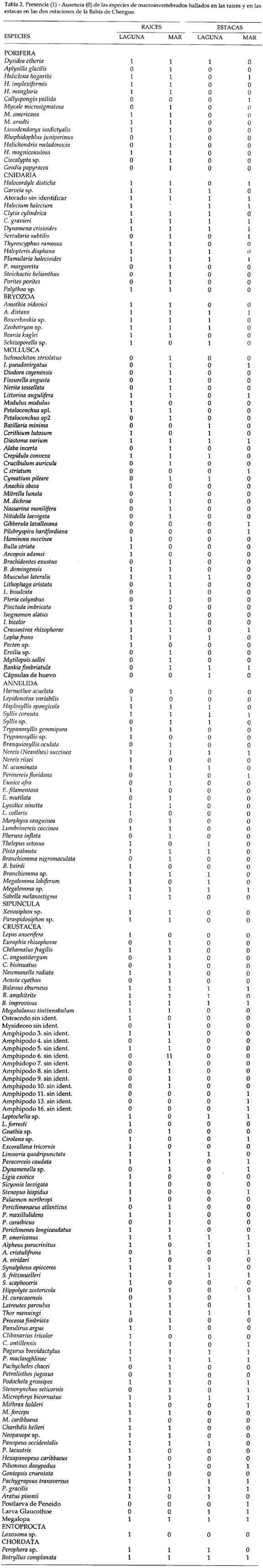
Los crustáceos, con 78 especies, fue el grupo más representativo en la Bahía de Chengue. De las 35 formas que colonizaron las estacas, dos no estuvieron presentes en las raíces (las postlarvas de peneido y glaucothoe). Los cirripedios B. eburneus y B. improvisus, los camarones Periclimenes americanus, Synalpheus fritzmuelleri y Thor manningi, los cangrejos Pagurus brevidactylus, P. maciauqhlinae, Microphrys bicornutus, Pachygrapsus transversus, P. gracilis y megalopas.
El cordado Botryllus complanata se encontró en las raíces y en las estacas en las dos estaciones.
En la mayoría de los muestreos se presentaron valores más bajos de diversidad en las estacas que en las raíces, a excepción de la estación LAG en diciembre (Tab. 3). En la estación LAG, la mayor diversidad en las raíces se presentó en septiembre (3.95) y en las estacas en diciembre (3.44). En la estación MAR, el valor más alto para las raíces se obtuvo en junio (4,71) y para las estacas en septiembre (3.80).
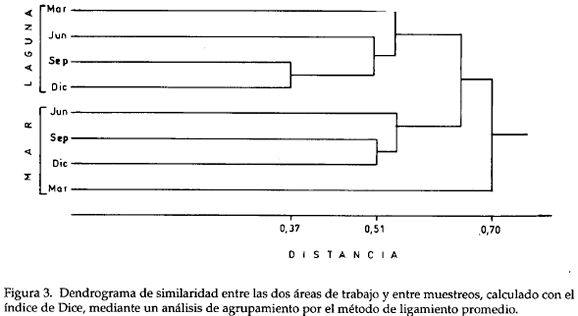
Del dendrograma (Fig. 3), se infiere la formación de dos grupos entre los muestreos de cada estación. Los resultados de septiembre y diciembre mostraron mayor grado de similaridad en cada una de las estaciones, que los efectuados en marzo.
El análisis de agrupamiento entre los muestreos de la fauna de las raíces de mangle y la de las estacas (presencia-ausencia) en las dos estaciones de Chengue, reveló la formación de dos subgrupos entre los muestreos de las raíces de la misma estación. Igualmente se formaron dos subgrupos entre los muestreos de las estacas (Fig. 4). Mostraron mayor similitud en los subgrupos, los muestreos de raíces y estacas realizados en septiembre y diciembre en la estación LAG y se agruparon los muestreos de junio y septiembre en MAR. La fauna colonizadora de las estacas en LAG, presentó mayor afinidad con la fauna asociada a las raíces en las dos estaciones de Chengue, que con la de las estacas en MAR.
DISCUSIÓN
La presencia en las estacas de numerosas especies observadas en las raíces de mangle, corrobora lo afirmado por Thorson (1964), con respecto a que muchas larvas de vida corta como las de briozoos, poliquetos y ascidias permanecen cerca de donde nacieron y colonizan muchas veces entre sus parientes, tal como se observó con los crustáceos, cnidarios, briozoos y anélidos.
El patrón de colonización inicial nunca es uniforme. Un determinado grupo taxonómico no tiene la hegemonía en la colonización de un sustrato vacío, sino el o los que gocen de ciertas características temporales de comportamiento reproductivo (Gómez y Belgrano, 1986). Sin embargo, existencia de especies colonizadoras permanentes indica desoves continuos. Este comportamiento es característico de aguas tropicales (Tarazona et al. 1983 en Gómez y Belgrano, 1986).
Thorson (1964) afirma que muchas larvas son fotopositivas, pero en el momento de colonizar invierten súbitamente su comportamiento y buscan Jugares sombreados u oscuros. Es probable que el arribo de las larvas a las estacas haya sido favorecido por la ubicación de éstas en el área sombreada entre el borde del manglar. Algunas formas vágiles como camarones y cangrejos pueden provenir de ecosistemas vecinos y llegar en estado juvenil o adulto, para utilizar el área del manglar como sitio de refugio, de alimentación y reproducción.
La colonización masiva de un sustrato por una especie en una sola época del año, podría obedecer a que algunas larvas de animales sésiles intermareales como cirripedios, serpúlidos, bivalvos y ascidias son gregarias. Estas larvas se ponen en contacto con especímenes ya fijados de la misma especie (Thorson, 1964).
El mayor número de especies arribó hacia septiembre y diciembre (época lluviosa) cuando se registraron temperaturas más altas y salinidades relativamente bajas (Reyes, 1991). Se detectó la influencia de la época de muestreo sobre la composición de la fauna colonizadora al presentarse mayor similitud en las estacas recolectadas en las dos estaciones en septiembre y diciembre (época lluviosa) (Reyes, 1991). Similarmente, Zea (1990) registró el mayor reclutamiento de esponjas en hábitats rocosos y coralinos de la región durante la estación lluviosa.
La menor variabilidad y los valores relativamente altos la diversidad en la estación LAG, muestra cierta continuidad en el aporte de larvas por parte de los donadores asociados al manglar, lo que indica la existencia de suficientes adultos durante el año. La mayor variabilidad del índice de diversidad en la estación MAR, puede ser el resultado de la combinación de varios factores, tales como ciclos reproductivos, cercanía de otros ecosistemas y patrones de conducta de las diferentes especies. Se esperaban valores bajos de diversidad para las estacas, debido a que se trata de comunidades pioneras (Margalef, 1974). Sin embargo, en general, los valores fueron altos, debido probablemente a que por el tiempo de exposición de las estacas (tres meses) fue suficiente para la adhesión de varias especies. Pudieron influir también el sustrato similar al natural y la cercanía de los donadores.
El valor más alto de diversidad en las estacas colocadas en el trimestre siguiente al mes de máxima diversidad en las raíces, indica que la disponibilidad de donadores en el medio afecta en gran medida la colonización de los sustratos.
La similaridad de la fauna colonizadora de las estacas por estación muestra diferencias producidas por la ubicación de las dos estaciones y en aspectos biológicos de las especies. El mayor grado de similaridad entre la fauna colonizadora de las estacas en la estación LAG con la fauna de las raíces de las dos estaciones (LAG y MAR), demuestra la influencia más directa de los organismos adheridos a las raíces de estos dos sitios. De otro lado, la menor similaridad entre la fauna colonizadora de las estacas en MAR con la fauna de las raíces muestra la influencia de ecosistemas vecinos.
AGRADECIMIENTOS
Este trabajo es parte de la tesis de Magister Scientiae presentado por la estudiante Rosalba Reyes a la Universidad Nacional de Colombia, dentro del convenio Universidad Nacional de Colombia - INVEMAR-COLCIENCIAS. Se contó con la colaboración de los especialistas de cada taxa: Zea, S. en Porifera; Flórez, L. en Hidrozoa; Manjarrés, G. en Anthozoa y Sipuncula; Pérez, L. en Bryozoa; Dueñas, R. en Annelida; Díaz, J.M. en Mollusca; Puentes, L. en Natantia y Sieg, J. en Tanaidacea. El proyecto fue financiado conjuntamente por COLCIENCIAS - INVEMARy el CINDEC de la Universidad Nacional de Colombia. Expresamos nuestros agradecimientos al Prof. S. Zea por su colaboración y aporte al desarrollo del proyecto y a R. Madera jefe del centro de cómputo.
BIBLIOGRAFÍA
1 Boesch, D.F. 1977. Application of numerical classification in ecological investigations of water pollution. Sepecial Scientific Report 77, Virginia Instit. Mar. Sci., 114 p. [ Links ]
2 Calderon-Sáenz, E. 1983. Hallazgo de Pelliciera rhizophorae Triana y Planchón (Theaceae) en la costa del Atlántico, con observaciones taxonómicas y biogeográficas preliminares. Bol. Mus. del Mar, 11:110-111. [ Links ]
3 Cintrón, G. y Y. Schaeffer-Novelli. 1983. Introducción a la ecología del manglar. UNESCO, Montevideo-Uruguay, 109 p. [ Links ]
4 Digby, P.G.N. y R.A. Kempton. 1987 Multivariate analysis of ecological communities. Champman and Hall. Londres, 106 p. [ Links ]
5 Gómez, E. y C. Belgrano. 1986. Estudio de colonización y sucesión de macrofauna bentónica en fondo somero a través de experimento in situ en la Bahía de Ancón. Tesis Biólogo Univ. Nal. Agraria la Molina, Lima, Perú, 68 p. [ Links ]
6 Margalef, R. 1974. Ecología. Editorial Omega, Barcelona, 367 p. [ Links ]
7 Palacio, J.A. 1983. Die benthische Makroinvertebratenfauna der tropischen Astuarregion Ciénaqa Grande de Santa Marta (Kolumbien) und ihre Aktivität im Wechsel zwischen Trocken und Regenzeit. Disertación, Univ. Bochum, RFA, 150 p. [ Links ]
8 Pannier, F. y R. Pannier. 1976. Interpretación fisiológica de la distribución de manglare; en la costa del continente suramericana. En: Memorias Seminario sobre el Océano Pacífico Sudamericano, Septiembre 1 - 5, 1976, Cali, Colombia,: 537-560. [ Links ]
9 Reyes, R. 1991. Macroinvertebrados asociados a las raíces de Rhizophora mangle Linnaeus, 1753 (mangle rojo), en la Ciénaga Grande de Santa Marta y en la Bahía de Chengue y Nenguange, Caribe colombiano. Tesis M. Sc., Univ. Nal. Colombia, 94 p. [ Links ]
10 Rützler, K. 1969. The mangrove community aspects of its structure faunistics and ecology. En: UNAM-UNESCO (E.). Mem. Simp Intern. Lagunas Costeras (Noviembre 28 - 30,1967), México D. F. 515-536. [ Links ]
11 Shannon, CE. y W. Weaver. 1963. The mathematical theory of communication Univ. Illinois Press, Urbana, 117p. [ Links ]
12 Sutherland, D. 1980. Dynamics of the epibenthic community on roots of mangrove Rhizophora mangle, at Bahía de Buche, Venezuela. Mar. Biol., 58:75-84. [ Links ]
13 Thorson, G. 1964. Ligth as an ecological factor in the dispersal and settlement of larvae of marine bottom invertebrates. Ophelia, 1(1):167-208. [ Links ]
14 Zea, S. 1987. Distribution, cover and recruitment of Demospoges (Porifera, Demospongie) in rocky and reefal habitats of Santa Marta, Colombian Caribbean. Tesis Ph. D., Univ. Texas, Austin, 154 p. [ Links ]
