










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.22 no.1 Santa Marta Jan./Dec. 1993
RECRUITMENT PATTERNS OF SESSILE INVERTEBRATES ONTO FOULING PLATES IN THE BAY OF SANTA MARTA, COLOMBIAN CARIBBEAN
Camilo B. García1 and Horst Salzwedel2
1Universidad Jorge Jadeo Lozano, Facultad de Biología Marina, Santa Marta, A, A. 5273, El Rodadero, Santa Marta, Colombia (C.B.G.).
2Corporación Autónoma Regional del Magdalena y de la Sierra Nevada, CORPAMAG-PROCIENAGA, calle 23 # 4-27, Santa Marta, Colombia. (H.S.).
ABSTRACT
Recruitment of sessile invertebrates onto fouling plates immersed for consecutive two-week periods during one year was found to correlate with the local climatic regime. For selected species it was found that they recruited at significantly higher levels in certain local climatic periods, but that these periods are not necessarily the same for the different species. A conceptual model is presented in which the recruitment process is divided in a pre-settlement and a post-settlement phase. The model attempts to predict under which combination of global circumstances periods of increased recruitment are to be expected. Application of the model to the data at hand highlights where the research needs for the future are.
RESUMEN
Se encontró que el reclutamiento de invertebrados sésiles en placas colectoras inmersas por periodos consecutivos de dos semanas durante un año, correlaciona con el régimen climático local. Para especies seleccionadas se encontró que reclutaron a niveles significativamente más altos en ciertos periodos del clima local, pero que estos periodos no son necesariamente los mismos para las diferentes especies. Se presenta un modelo conceptual en el cual el proceso de reclutamiento se divide en una fase pre-asentamiento y una post-asentamiento. El modelo pretende predecir bajo que combinación de circunstancias globales serían de esperarse periodos de mayor reclutamiento. Aplicación del modelo a los datos disponibles pone de relieve cuales son las necesidades futuras de investigación.
INTRODUCTION
The recruitment issue is becoming central for our understanding of the formation and development of invertebrate communities (Connell, 1985; Bhaud, 1988; Fairweather, 1991). One aspect to consider is that of the periodicity of recruitment. In temperate waters it is a well established fact that most spawning and recruitment occurs in the warmest months of the year, concomitantly with the seasonal increase in primary production (Cecere and Matarrese, 1983; Grahame and Branch, 1985; Starr et al., 1990). In tropical waters the recruitment of invertebrates is generally not confined to a definite period in the year, but tends to occur all year around. However, it has been repeatedly observed that during the recruitment seasons the intensity of recruitment fluctuates, although not synchronously among species (Sutherland, 1980; Alongi, 1989; Hatcher et al., 1989).
The combination of bioecological factors that contribute to such periods of increased recruitment in the tropics has been less studied. A fundamental question is why should there be periods of increased recruitment pressure in the tropics with their low amplitude fluctuations in climatic and environmental conditions: dry season/rainy season oscillation (Longhurst and Pauly, 1987; Hatcher et al., 1989). A first partial answer may view this as a consequence of increased synchronous reproductive activity of parental populations. Such increased reproduction obeys the constraint for sessile invertebrates to optimize the fertilization rate (Grahame and Branch, 1985; Cameron, 1986). However, mortality of larvae in the plankton is substantial (Thorson, 1950; Rumrill, 1990), so that recruitment fluctuations do not necessarily reflect fluctuations in initial number of larvae.
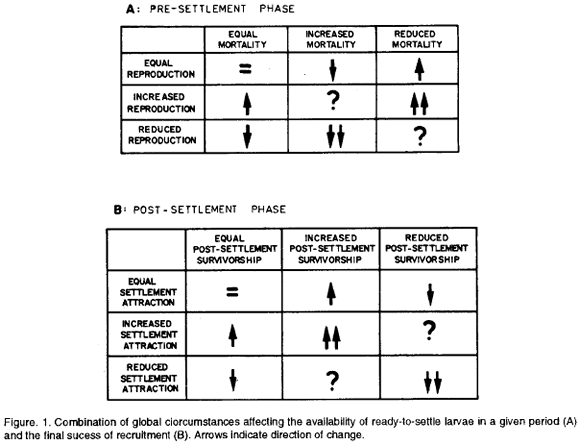
For the sake of study, the processes leading to the recruitment of sessile invertebrates can be divided into a pre-settlement phase and a post-settlement phase. For each of these phases it is possible to construct a matrix of global circumstances showing the combinations which would affect the number of ready-to-settle larvae and the final success of recruitment. Such matrices are shown in Fig. 1. One global circumstance apparently not considered in the pre-settlement matrix is the effect of transport by currents away from potential suitable substrata for settlement. Operationally, however, transport away from a given substratum can be considered equivalent to increased larval mortality (Fig. 1, matrix A) in terms of the availability of larvae capable of settling on that substratum. The contrary would also be truth: transport to the substratum can be considered equivalent to increased reproduction.
Justification for the post-settlement matrix is the observation that settlement is also influenced by larval selectivity (Hadfield, 1986; Chia,1990; Young, 1991) and that post-settlement mortality can be considerable (Connell, 1985; Zajac et al., 1989; Hurlbut, 1991). Thus, for successful efective recruitment it is not sufficient that settlement takes place. The settlers should put up with the particular environmental characteristics of their settling places. A possible artifact in evaluating post-settlement mortality, i.e. early mortality, is the time between censuses. Clearly, the longer the time between censuses, the stronger the impact of mortality on the observations.
In this paper recruitment patterns of sessile invertebrates on fouling plates in a tropical system is examined in the context of the conceptual model presented above as far as possible with the available data. More than to reach some conclusion, the purpose of the exercise is to signal where research efforts should be put in the future. On the basis of the results provided by both non-metric multidimensional scaling (NMDS) and classification analysis to group plates inmersed in consecutive two-week periods (see below) we specifically test two null hypotheses which concern the relation between recruitment and the local climatic regime. For all the periods as discriminated by NMDS and classification, which correspond fairly well with local climatic fluctuations (see below), we hypothetize, first, that the intensity of recruitment either total or for selected species is the same, and second, that the growth rate of selected species expressed as mean size reached after two weeks of immersion, is also the same.
The second hypothesis is an a-posteriori attempt to explain the existence of increased invertebrate recruitment periods in the system studied in terms of availability of food expressed as higher growth rates.
STUDY SITE
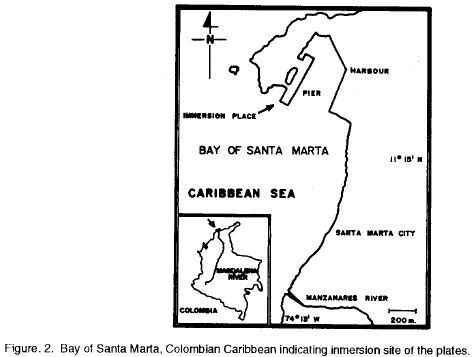
The study was carried out under a Pier in the Bay of Santa Marta (11°15'08"N; 74°13'13"W), Colombian Caribbean. See map with study site in Fig. 2. The Bay of Santa Marta lies at the margin of the Sierra Nevada de Santa Marta, a huge mountainous area, which is the main determinant of the local climate affecting direction and velocity of winds, and distribution of rain (Müller, 1979). Two main seasons have been distinguished during the year: the dry season from December to April, and the rainy season from May to November, with a short dry period from July to August called "Veranillo de San Juan" (Müller, 1979; Ramírez 1983; Salzwedel and Müller 1983). Figure 3 shows the generalized annual patterns of precipitation, water surface temperature and salinity for the Bay of Santa Marta.
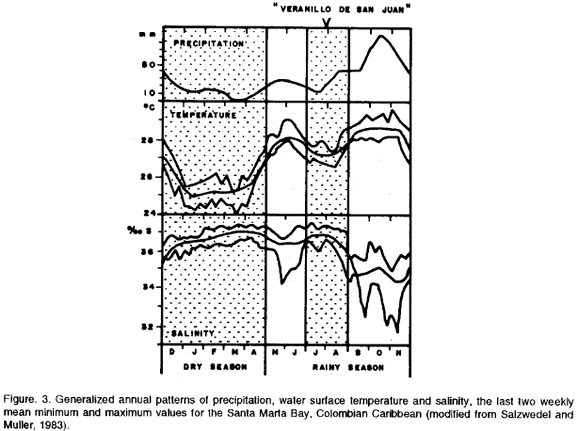
During the dry season the trade winds blowing northeast reach their highest intensity and velocity, and displace surface water off the bay. This causes an upwelling phenomenon which brings cooler and more saline waters into the bay. During the rainy season with calmer winds, temperature rises and salinity decreses due to the increased discharge from the Manzanares River (and marginally from the Magdalena River). In July-August ("Veranillo de San Juan") a small but marked decrease in temperature and increase in salinity has been observed (Müller, 1979; Ramírez, 1983; Salzwedel and Müller, 1983).
MATERIAL AND METHODS
Asbestos plates (10 cm X 10 cm) were used to simulate discrete, small isolated patches of habitat. The plates were fixed to a PVC frame tied to the bottom under a pier and maintained up right by means of buoys so that the plates were placed at approximately 9 m depth. Only the side of the plates facing outside of the pier was studied.
The sampling was done biweekly starting April 30, 1981 and ending April 29, 1982. On each sampling date two plates were taken out, one of which had been immersed at the start and the other two weeks before sampling, and replaced in the frame by new ones. Plates were also regularly photographed in situ (35 mm slides). Thus, three series of plates and a photographic record of different stages of development were obtained. We will deal here with the second series, i.e. the series that reflects the bi-weekly potential of recruitment (species which may recruit in a given two week period). Observations on the other two series of plates will be reported elsewhere.
Unfortunately in storage the plates dried out, precluding the identification of soft-bodied organisms (Porifera and Tunicata). Porifera did recruit on the two-week plates as indicated by the slide analysis. They did so, however, at too low rates, and growth within the two-week periods was too slow to permit safe quantification from the slides of number of colonies.
Solitary sessile organisms, i.e. serpulid polychaetes and barnacles, were counted and measured, when feasible (see below). In the case of colonial organisms (encrusting bryozoans) number of colonies were counted. For the analyses bryozoans were pooled as one group since they were all encrusting forms and species were not identified. A number of serpulid species were also pooled in groups on account of their very similar tubes, which from the two-week plates could not safely be told apart. These are Hydroides cf brachyacanthus, Hydroides parvus and Protula sp. henceforth referred to as Hb/Hp/P, Salmacina sp., Filograna sp. and Josephella marenzelli, henceforth referred to as S/F/J, and Pseudovermilia multiespinosa and Pseudovermilia occidentalis, henceforth referred to as Pseudovermilia spp. Other species recruiting to the two-week plates were the serpulid polychaetes Pomatoceros minutus, Spirorbinae 1,2 and 3 and the cirriped Balanus trigonus.
For summarizing and identifying patterns in the data we made use of clasification and ordination techniques. Dendrograms were made using the Bray-Curtis measure of distance for constructing the dissimilarity matrix, and the unweighted pair groups method using arithmetic averages (UPGMA) as grouping strategy. The same dissimilarity matrix was used to perform nonmetric multidimensional scaling (NMDS) on the data (Gauch, 1982; Ludwig and Reynolds, 1988). Analyses of variance, most of the time parametric but in some instances nonparametric, were applied when performing univariate comparisons, complemented with a-posteriori multiple comparisons (Tukey HSD test, Siegel, 1956; Underwood, 1981). The plates within one period were considered as replicates for the analysis on recruitment intensity. For the analysis on growth rate (mean size) data within one period are pooled. Homogeneity of variance was tested and the data transformed when necessary (see text). Correlations between abundances and sizes of recruits were examined by means of the Spearman rank correlation coefficient (Siegel, 1956).
RESULTS
For most species, recruitment was found to occur rather continually during the period of immersion (Fig. 4A-D). The exception was Spirobinae 1, which did not recruit during weeks 41-52 (Fig. 4B). Recruiment intensity, though, was found to vary during the year of immersion. The most conspicuos variation was shown by B. trigonus, which during weeks 0-34 exhibited low recruitment rates. After this point recruitment increased dramatically, reaching two strong pulses in weeks 41-42 and 45-46 (Fig. 4A). This pattern causes total recmitment apparently to peak at these dates, but when the contribution of B. trigonus is subtracted, a decresing tendency in total recruitment during weeks 35-52 (Fig. 4A) appears. The more important contributors to total recruitment were Spirorbinae 3 with 2407 individuals, B. trigonus with 2342 individuals, S/F/J with 1775 individuals and bryozoans with 999 colonies. The other species recruited at rates one order of magnitude lower (Fig. 4A-D).
Nonmetric multidimensional scaling (NMDS) and clasiffication performed on abundance data log (X+l) transformed (Underwood, 1981), show a grouping into four periods that closely fits the climatic fluctuations in the area. Thus, plates immersed during May-June (first part of the rainy season), July-August (short dry period "Veranillo de San Juan"), September-December (Second part of the rainy season and beginning of the dry season) and January-April (dry season) cluster apart (Fig. 5A-B). On this basis the two null hypotheses mentioned in the introduction refering to recruitment intensity and growth rates in relation to local climatic regime, were tested.
Intensity of total recruitment (all data together) did not differ between periods, nor were differences found for any pair of means (Table 1). However, as above when the contribution of B. trigonus is subtracted, recruitment does show different intensities in the different periods (Table 1). The a-posteriori multiple comparisons test shows that total recruitment to the two-week plates (without B. trigonus) occurred in a pattern which is somewhat shifted with respect to the generalized climatic annual changes (Fig. 3), recruitment concentrating in the rainy season (Table 1).
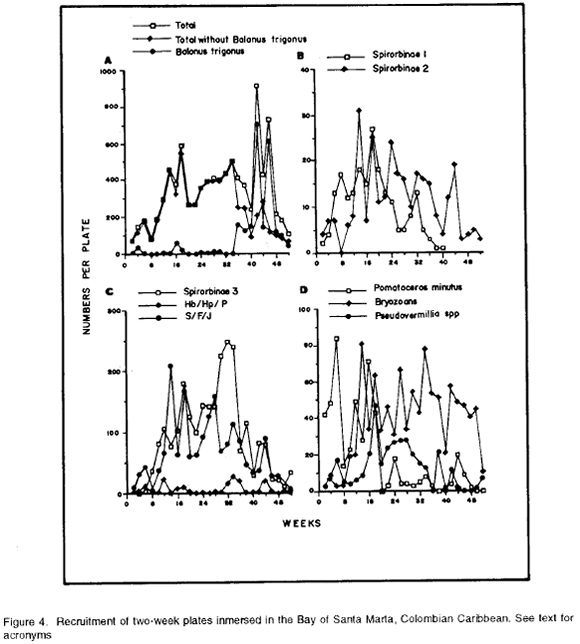
As expected from the results above, ANOVA suggests differences in recruitment intensity of B. trigonus in the four periods (Table 1). The multiple comparisons test indicates that mean recruitment intensity in period 4 (January-April, corresponding to the dry season, Figs. 3 and 5A-B) was significantly higher compared to the other periods, for which it did not differ (Table 1). Clearly B. trigonus exhibits a strong seasonality in recruitment in the year of immersion of the plates, this being concentrated in the dry season.
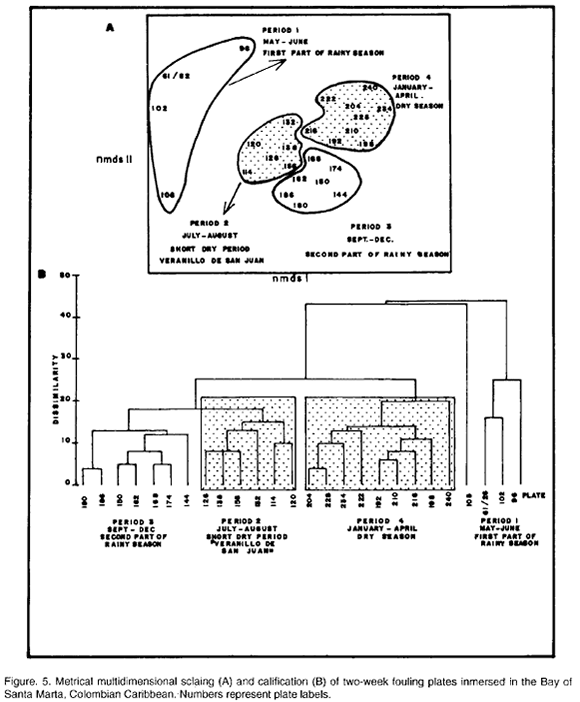

Intensity of recruitment of P. minutus also differs for the four periods (Table 1). The a-posteriori multiple comparisons test shows a main recruitment season extending over periods 1 and 2 (May-August, Fig. 5A-B) which correspond with the first part of the rainy season and the "Veranillo de San Juan" (Fig. 3), while the rest of the year was of consistently lower recruitment intensity (Table 1).
Spirorbinae 3 shows a more complex behavior. Recruitment intensity differs between periods (Table 1). The four periods form a hierarchy of mean recruitment intensities. Thus, as the first part of the rainy season progressed and through the "Veranillo de San Juan" recruitment intensity increased from the lowest level to peak in the second part of the rainy season and from there to drop then in the dry season to a level similar to that in the "Veranillo de San Juan" (Table 1, Fig. 4C).
Although overall differences in recruitment intensity of Spirorbinae 2 between the periods were found (ANOVA, Table 1) no single pairwise difference in mean recruitment intensity turned out to be significant (Table 1). Looking at Figure 4B marked fluctuations spanning the year can be seen. This suggests only weak differences between periods. No particular pattern in recruitment intensity is apparent for this species.
In contrast, bryozoans show a defined pattern of mean recruitment intensities. First, the periods are different from one another (Table 1). The multiple comparisons test (Table 1) shows that period 1 (May-June, first part of rainy season Fig. 5A-B) exhibits a significantly lower mean recruitment intensity in relation to the other 3 periods, which are very similar.
In order to test the second null hypothesis, namely, that the growth rate of recruits expressed as mean size reached after two weeks of immersion of the plates does not vary between the periods identified by NMDS and clasification (Fig. 5A-B), we selected four species: Balanus trigonus, Pomatocerus minutus, Spirorbinae 3 and Spirorbinae 2.
B. trigonus grew at different rates in the four periods (ANOVA, Table 1). The Tukey HSD test (Table 1) clearly identifies period 2 (July to August, short dry period "Veranillo de San Juan", Fig. 5A-B) as the period in which B. trigonus reaches a maximun mean size for the four periods (Table 1).
For P. minutus heterogeneity of variances could not be removed. However, since the ANOVA F probability is highly significant (p= 0.000) and nonparametric Kruskal-Wallis one way analyses of variance on the same data and periods is consistent (p=0.000) with its parametric equivalent, the parametric ANOVA will be considered valid here. As with B. trigonus, period 2 (July-August, "Veranillo de San Juan, Fig 5A-B) is the one with significantly higher growth rate, while for the rest of the year the growth rate did not vary (Table 1, Tukey HSD test).
For Spirorbinae 3 heterogeneity of variances could likewise not be removed. But based on the same arguments as above incluiding the Kruskal-Wallis one-way analyses of variance (p=0.000) the parametric ANOVA is also considered valid in this case. This species shows significant differences in growth rates that form a hierarchy (Table 1). Maximum growth rates occur during the first part of the rainy season and "Veranillo de San Juan" then gradual decline through the second part of the rainy season to a minimum growth rate in the dry season.
With Spirorbinae 2 the same situation was found as above with respect to heterogeneity of variances. However, this case does not need much argumentation as the ANOVA F turned out to be non-significant and this result is reliable (Underwood 1981). No differences were found between any pairwise comparison of mean size (Tukey HSD test, Table 1). Thus, during the year of immersion of the plates this species grew at similar rates the entire year.
Associations between recruitment rate and growth rate of recruits of the selected species were investigated by means of Spearman rank correlation coefficients. Comparisons were made considering the plates both individually and grouped in the same fashion as suggested by ordination and classification (Fig. 5A-B). Compared were abundance of recruits (or mean abundance when comparing by groups) versus mean size (per plate and per group, respectively) of each of the selected species. This comparison addressed the question whether assumed better feeding conditions (manifested as larger size) result in increased recruitment.
Only in one case was a significant positive association found: between abundance and mean size of Pomatoceros minutus, considering the plates individually (p < 0.05). Thus, at least for the year of immersion of the plates, environmental conditions promoting increased recruitment were apparently unrelated to environmental conditions leading to higher growth rates.
DISCUSSION
The expectation that recruitment in the Caribbean tends to occur all year round was substantiated by this study. The recruitment patterns, however, were complex, though apparently related to the local climatic regime. Four different recruitment modalities for non-colonial species could be identified: (1) Recruitment occurring most of the year but failing in one definite period. An example is Spirorbinae 1, which was not found on the two-week plates from week 42 to week 52, second part of the rainy season. (2) Recruitment occurring all year round but with definite periods of significantly higher recruitment. Balanus trigonus, for instance, recruited preferentially in the dry season, while Pomatoceros triqueter recruited preferentially in the first part of the rainy season. (3) Gradually increasing recruitment reaching a maximum within a definite period, after which recruitment significantly decreases, thus suggesting a broad semicyclical pattern during the year. The recruitment of Spirorbinae 3, for example, increased from the beginning of the rainy season through the "Veranillo de San Juan" to reach a maximum in the second part of the rainy season, falling then significantly in the dry season. And (4) Recruitment with a nearly constant intensity all year round or with intensity varying independently of climatic change. Spirorbinae 2, for instance, showed no preference as to the season of recruitment.
Similar habits of recruitment have already been noted for other tropical systems, such as sandy beaches and subtidal shallow soft-bottoms (Alongi, 1989 and references therein), rocky shores (Ortega, 1987; Sutherland, 1980) but also for fouling systems (e.g. Galán, 1976). Thus, there appears to be an increasing amount of evidence supporting the assertion that recruitment in the tropics operates in a variety of forms of which continuous recruitment at constant rate is only one among several and probably the less usual. The different forms imply fluctuations of the intensity of recruitment in definite periods during the year which often can be correlated with climatic fluctuations as in the present study. The recruitment fluctuations of different species do not need to occur simultaneously or to be of the same sign. In fact total recruitment intensity was found not to vary with climatic change in this study, which is explained by the different periods of increased (or decreased) recruitment that cancel total recruitment differences between climatic periods. However, as mentioned above, if the contribution of Balanus trigonus is ignored, a net tendency of increased recruitment in the rainy season emerges, which is interesting because recent studies on demosponges in the area (Zea, 1992A, B) have shown that their recruitment to fouling plates attached against hard bottoms was concentrated in the rainy season as well.
The causes of periods of significantly higher recruitment pressure, as show by the two-week plates in the study site, requires an explanation. Not less important is to try to explain why such periods occured in different times of the year for the different species and their apparent relation to the local climatic regime.
In the conceptual model presented in Fig. 1 the process of recruitment was divided in two phases, one concerning the availability of ready-to-settle larvae (Fig. 1, Matrix A) or pre-settiement phase and the other concerning actual settlement (Fig. 2, Matrix B) or post-settlement phase. The question here is: which combination of global circumstances can explain the observed recruitment patterns in the fouling system studied.
For the pre-settlement phase it can be seen in Fig. 1 (Matrix A) that there are three combinations of global circumstances which would lead to increased numbers of ready-to-settle larvae. Of these three, the combination equal reproduction/reduced mortality probably does not apply for the present case, for it has been shown that tropical invertebrates undergo fluctuations in reproduction which may be correlated with climatic fluctuations (e.g. Alongi 1989 and references therein). More realistic appear the combinations increased reproduction-equal mortality, and increased reproduction/reduced mortality, the last one resulting in the highest number of ready-to-settle larvae.
The next step is actual settlement. It would seem reasonable to assume that the plates used were neither particulary attractive nor repellent to the larvae. Thus, only two global interactions appear relevant in this phase: equal settlement attraction/equal post-settlement survivorship and equal settlement attraction/increased post-settlement survivorship (Fig. 1, Matrix B), the last one leading to the highest effective recruitment.
If we assume that the main factor affecting post-settlement survivorship was feeding conditions (e.g. Zajac et al, 1989), which does not seem unrealistic as space was not at a premium, there was no interference by adults, and predation on the recruits is unlikely because of the plates being isolated, we may expect a positive correlation between periods of increased recruitment and mean size reached in two weeks of sessile life. That was not the case. The consistent lack of correlation between abundance and size of recruits found in this study suggests that conditions promoting higher recruitment are uncoupled from the conditions promoting growth of recruits. Therefore, in the post-settlement phase the combination of global circumstances equal settlement attraction/increased post-settlement survivorship does not seem to apply. If we consider the combination equal settlement attraction/equal post-settlement survivorship as the more likely combination, it follows that periods of increased recruitment are a function only of the occurence in the pre-settlement phase (Fig. 1, Matrix A). Clearly, the problem is that we do not know when the assumed periods of increased reproduction (see above) took place and cannot make estimations because we do not know the lenght of planktonic life for the different larvae nor where they come from. The extent of mortality in the water column is another open question, perhaps the central one.
As mentioned in the section Study Site, the dry season (December to April) in the Santa Marta area is related to an upwelling phenomenon (Müller, 1979; Ramírez, 1983; Salzwedel and Müller, 1983). However, there has been, to our knowledge, no study on primary production in the Bay of Santa Marta, and there is reason to suspect that in the rainy season the entry of nutrients carried by continental water is not less important, as found by Ramírez (1987) for the Bay of Nenguange, which lies not much further to the north of the Bay of Santa Marta. Moreover, the same author in a later study concluded that upwelling in the Santa Marta area has mostly physical rather than biological consequences due to the oligotrophic nature of the water transported to the photic zone (Ramírez 1990). Thus, it seems unlikely that a significant change in primary production during the dry season occurs. In fact, there are indications that, if there is an increase in primary production at all, it occurs in the rainy season. For instance, Caycedo (1977) for the year 1974-1975 recorded in the Bay of Nenguange the largest phytoplankton bloom at the beginning of the rainy season. In the present study three of the four species considered grew at higher rates in the first part of the rainy season and in the "Veranillo de San Juan" (see Results).
If we assume that changes in primary production are unimportant, we should then postulate changes in temperature, salinity, turbidity and even moon phases (e.g. Lessios, 1991) or some combination of these factors as triggers for increased reproduction and as factors potentially affecting mortality in the water column. For instance, for demospongie in the area, Zea (1992A) found a close correlation between recruitment and high temperatures and low salinities as they occur in the rainy season (see Zea, 1992A for a discussion on these factors).
Questions as to why there should be periods of increased recruitment, why these periods were found apparently to correlate with the local climatic regime, and why these periods differ from one species to the other (although there seems to be a certain convergence in the rainy season) in the system studied cannot be answered at this stage. Most intriguing are situations in the model where the sign of fluctuations in number of ready-to-settle larvae and success of recruitment are unpredictable (Fig. 1). We hope, however, to have shown some of the needs for future research.
ACKNOWLEDGEMENTS
This paper is based on a Dr. rer. nat. dissertation by C.B. Garcia, Biology/Chemistry Faculty, University of Bremen, Germany. The first author has been financially supported by a scholarship of the Deutscher Akademischer Austausch Dienst, DAAD. Field work was financed by the Gesellschaft für Technische Zusammenarbeit, GTZ and supported by the Instituto de Investigaciones Marinas de Punta de Betín, INVEMAR. Laboratory and data analysis were carried out at the Alfred Wegener Institut für Polar- und Meeresforschung. Dr. Harry A. ten Hove (Institut of Taxpnomic Zoology, Amsterdam) kindly identified the serpulid poiychaetes. This paper was oraly presented in the VIII Seminario Nacional de Ciencias y Tecnologías del Mar, Santa Marta, 1992. Comments of the anonymous reviewers substantially improved the manuscript.
REFERENCES
1 Alongi, D. M. 1989. Ecology of tropical soft-bottom benthos: a review with emphasis on emerging concepts. Revista de Biología Tropical 37(1):85-100. [ Links ]
2 Bhaud, M. 1988. La biologie larvaire chez les invertébrés marins: une question d'actualité. Vie Milieu 38(1): 19-24. [ Links ]
3 Cameron, R. A. 1986. Introduction to the invertebrate larval ecology workshop: a brief background. Bulletin of Marine Science 39(2): 145-161. [ Links ]
4 Caycedo, I. E. 1977. Fitoplancton de la Bahía de Neguange (Parque Nacional Tayrona), mar Caribe, Colombia. Anales del Instituto de Investigaciones Marinas de Punta de Betín 9: 17-44. [ Links ]
5 Cecere, E. and Matarrese, A. 1983. Settlement periods of barnacles in the mar picolo of Taranto (Southern Italy) from 1972 to 1982. Obalia 9:105-117. [ Links ]
6 Chia, F. 1990, Differential larval settlement of benthic marine invertebrates, 3-12. In: Reproduction, genetics and distribution of marine organisms (Ed by J. Ryland and P.J. Tyler). Olsen and Olsen. [ Links ]
7 Connell, J.H. 1985, The consequences of variation in initial settlement vs post-settlement mortality in rocky intertidal communities. Journal of Experimental Marine Biology and Ecology 93:11-45. [ Links ]
8 Fairweather, P. G. 1991. Implications of "supply-side" ecology for environmental assessment and management. Trends in Ecology and Evolution 6(2):60-63. [ Links ]
9 Galán, A. 1976. Contribuciones al estudio de las incrustaciones biológicas en la laguna de la Restinga, Isla Margarita, Venezuela. Boletín Instituto Oceanográfico Universidad de Oriente 15(2): 153-168. [ Links ]
10 Gauch, H. G., 1982. Multivariate analysis in community ecology. Cambridge University Press, 298 pp. [ Links ]
11 Grahame, J. and Branch, G. M. 1985. Reproductive patterns of marine invertebrates. Oceanography and Marine Biology Annual Review 23:373-398. [ Links ]
12 Hadfield, M. G. 1986. Settlement and recruitment of marine invertebrates: a perspective and some proposals. Bulletin of Marine Science 39(2):418-425. [ Links ]
13 Hatcher, B. G.; Johannes, R. E. and Robertson, A.I. 1989. Review of research relevant to the conservation of shallow tropical marine ecosystems. Oceanography and Marine Biology Annual Review 27:337-414. [ Links ]
14 Hulrbut, J-C. 1991. Larval substratum selection and post-settlement mortality as determinants of the distribution of two bryozoans. Journal of Experimental Marine Biology and Ecology 147:103-109. [ Links ]
15 Lessios, H. A. 1991. Presence and absence of monthly reproductive rhythms among eight Caribbean echinoids off the coast of Panama. Journal of Experimental Marine Biology and Ecology 153: 27-47. [ Links ]
16 Longhurst, A. L. and Pauly, D. 1987. Ecology of tropical oceans. Academic Press, Inc. 407 pp. [ Links ]
17 Ludwig, J. A. and Reynolds, J. F. 1988. Statistical ecology. A prime on methods and computing. John Wiley and Sons, 337 pp. [ Links ]
18 Müller, K. 1979. Interrelaciones entre salinidad y temperatura en la Bahía de Santa Marta. Anales del Instituto de Investigaciones Marinas Punta de Betín 11:219-226. [ Links ]
19 Ortega, S. 1987. Habitat segregation and temporal variation in some tropical intertidal populations. Journal of Experimental Marine Biology and Ecology 113: 247-265. [ Links ]
20 Ramírez, G. 1983. Características físico-químicas de la Bahía de Santa Marta (agosto 1980-julio 1981). Anales del Instituto de Investigaciones Marinas Punta de Betín 13: 111-121. [ Links ]
21 Ramírez, G. 1987. Características hidroquímicas y composición química de los sedimentos de la Bahía de Neguange, Caribe Colombiano. Anales del Instituto de Investigaciones Marinas Punta de Betín 17:15-26. [ Links ]
22 Ramírez, G. 1990. Distribución de nutrientes inorgánicos en las aguas costeras de la región de Santa Marta, Caribe colombiano, 244-254. En: Memorias VII Seminario Nacional de las Ciencias y Tecnologías del Mar, Cali. Comisión Colombiana de Oceanografía. [ Links ]
23 Roughgarden, J.; Gaines, S. and Possingham, H. 1988. Recruitment dynamics in complex life cicles. Science 24:1460-1466. [ Links ]
24 Rumrill, S. S. 1990. Natural mortality of marine invertebrate larvae. Ophelia 32(1-2):163-198. [ Links ]
25 Salzwedel, H. and Müller, K. 1983. A summary of metereological and hidrological data from the Bay of Santa Marta, Colombian Caribbean. Anales del Instituto de Investigaciones Marinas Punta de Betín 13:67-83. [ Links ]
26 Siegel, S. 1956. Nonparametric statistics for the behavioral sciences. McGraw-Hill, 312 pp. [ Links ]
27 Starr, M.; Hirnmelman, J. H. and Therriault, J-C. 1990. Direct coupling of marine invertebrate spawning with phytoplankton blooms. Science 247:1071-1074. [ Links ]
28 Sutherland, J. P. 1980. Dynamics of the epibenthic community on roots of the mangrove Rhizophora mangle, at Bahía de Buche, Venezuela. Marine Biology 58:75-84. [ Links ]
29 Thorson, G. 1950. Reproductive and larval ecology of marine bottom invertebrates. Biological Review 25:1-45. [ Links ]
30 Underwood, A. J. 1981. Techniques of analysis of variance in experimental marine biology and ecology. Oceanography and Marine Biology Annual Review 19:513-605. [ Links ]
31 Young, B. L. 1991. Spartina axis zones: prefered settlements sites of barnacles. Journal of Experimental Marine Biology and Ecology 151:71-82. [ Links ]
32 Zajac, R-N.; Whitlatch, R. B. and Osman, R. J. 1989. Effects of inter-specific density and food supply on survivorship and growth of newly settled benthos. Marine Ecology Progress Series 56:127-132. [ Links ]
33 Zea, S. 1992a. Estimation of demosponge (Porifera, Demospongiae) larval settlement rates from short-term recruitment rates: preliminary experiments. Helgolánder Meeresuntersuchungen 46(3): 293-300. [ Links ]
34 Zea, S. 1992b. Recruitment of Demosponges (Porifera, Demospongiae) in rocky and coral habitats of Santa Marta, Colombian Caribbean. Marine Ecology. P. S. Z. N. I. 13(1): in press. [ Links ]
