










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.22 no.1 Santa Marta Jan./Dec. 1993
DISTRIBUCIÓN ESPACIAL DE LA TAXOCENOSIS ANNELIDA-MOLLUSCA EN LA PLATAFORMA CONTINENTAL DEL GOLFO DE SALAMANCA, CARIBE COLOMBIANO
Ángela Guzmán-Alvis1 y Juan Manuel Díaz2
1Apartado Aéreo 5273, El Rodadero-Santa Marta, Colombia (A.G.).
2Instituto de Investigaciones Marinas de Punta de Betín (INVEMAR), Apartado Aéreo 1016, Santa Marta, Colombia (J.M.D).
RESUMEN
Se hace una caracterización espacial preliminar de las comunidades macrozoobénticas de la plataforma continental al suroccidente de Santa Marta, Caribe colombiano. Para tal fin, se tomaron muestras de fondo en 17 estaciones, entre los 15 y 100 m de profundidad. El análisis de la taxocenosis se basa en aquellos grupos más abundantes, poliquetos y moluscos, analizados a nivel de familias. Se utiliza un análisis de agolpamiento entre estaciones y entre familias con el fin de caracterizar espacial-mente la taxocenosis. La distribución espacial de la taxocenosis se relaciona con la profundidad, la composición granulomótrica del sedimento y algunas características biológicas de sus componentes. Les resultados preliminares muestran básicamente tres taxocenosis: la primera se caracteriza por encontrarse entre 15 y 60 m de profundidad, en fondos heterogéneos de cienos y arenas (medias, finas y muy finas), presentando la mayor abundancia y diversidad; la segunda se presenta sobre los 60 m de profundidad, en fondos de cieno; la tercera asociación se presenta sobre los 100 m, en cieno y mues&tra las menores abundancias y diversidades.
ABSTRACT
A preliminary characterization of macrozoobenthic assemblages on the continental shelf off southwestern Santa Marta, Colombian Caribbean, is made on the basis of bottom samples taken at 17 stations, in depths between 15 and 100 m. Only families of the most abundant groups, Mollusca and Polychaeta, were taken into account for the analysis. Cluster analysis of similarity between stations and between families revealed the ocurrence of three assemblages, whose spatial distribution is related with sediment type, depth, and some biological features: one assemblage is found on bottoms with fine to very fine sand and muds at dephts between 15 and 60 m, showing the greater abundances and families diversity; the second assemblage is found at about 60 m depth in muddy bottom; a last assemblage inhabits also muddy bottom below 100 m depth and exhibits the lowest numbers of abundance and diversity.
INTRODUCCIÓN
Las comunidades macrozoobentónicas de fondos blandos en regiones tropicales han sido menos estudiadas en comparación con las de regiones subtropicales y templadas. Un mayor cúmulo de información sobre el bentos de regiones tropicales es necesario para confirmar, refutar o modificar los conceptos generales de la ecología del bentos, formulados a partir del conocimiento que se tiene sobre las comunidades de las zonas templadas (Alongi, 1989).
En el Caribe colombiano son pocos los estudios realizados sobre macrofauna bentónica de fondos blandos de plataforma. Algunos incluyen un sólo grupo taxonómico, como los ostrácodos de la plataforma de la Guajira (Ramírez, 1983), los crustáceos en la región de Santa Marta (Puentes et al, 1990) o los moluscos en la bahía de Nenguange (Díaz y Gotting, 1986); otros analizan varios grupos, como los realizados en la Bahía de Cartagena (Mora y Prieto, 1978), en el Golfo de Urabá (López, 1981) y en la zona somera de la plataforma frente a la Ciénaga Grande de Santa Marta (García y Sandoval, 1983; García y Salzwedel, 1991). La mayoría de estos trabajos ha sido realizada en fondos someros, por encima de la isóbata de 40 m. El único trabajo que abarca profundidades superiores se refiere a los micromoluscos recientes y subfósiles de la plataforma continental entre Bocas de Ceniza y el Golfo de Morrosquillo (Rodríguez, 1983).
El presente trabajo caracteriza espacialmente las taxocenosis bentónicas Annelida-Mollusca en la plataforma del Golfo de Salamanca con base en su composición por familias y estructura (dominancia, abundancia y diversidad), abarcando profundidades entre los 15 y 100 m. Además, pretende determinar en forma general las relaciones entre la distribución de las taxocenosis con la calidad textural del sedimento, la profundidad y algunos rasgos biológicos de los organismos, especialmente sus hábitos alimenticios y sus patrones de movilidad.
ÁREA DE ESTUDIO
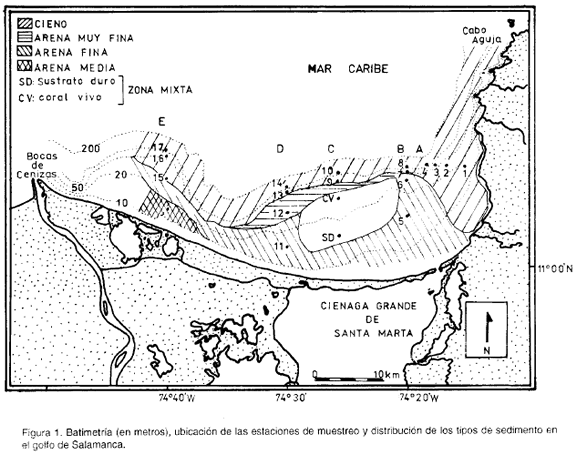
La región forma parte del Golfo de Salamanca y se encuentra ubicada entre los 11°00' y 11°10' N, y 74°12 y 74°40' W, en el Caribe Colombiano (figura 1). Blanco et al. (1992) definen el Golfo de Salamanca como el cuerpo de agua comprendido entre la línea de costa que va desde el Cabo de la Aguja hasta Bocas de Cenizas.
Morfológicamente, la plataforma continental en el área de estudio se caracteriza por ser relativamente amplia en comparación con la de las áreas adyacentes (Molina, 1990; CORPES, 1992), presentando su máxima extensión frente a la Ciénaga Grande de Santa Marta (16.5 km). La sedimentación es esencialmente de origen continental (Lorin et al., 1973; Erffa, 1973; García y Sandoval, 1983), constituida predominantemente por arenas grises de grano fino, distribuidas en la parte interna de la plataforma, y por lodos depositados cerca a la desembocadura del río Magdalena y en la parte externa de la plataforma. Presenta también algunos parches de material biodetrítico y restos dispersos de corales fósiles (Blanco, 1988; Molina, 1990).
La línea costera del área de estudio ha sufrido cambios morfológicos significativos en tiempos recientes, estando sometida a corrientes que no son uniformes a lo largo de la costa (Erffa, 1973). Estas corrientes de deriva costera, originadas por la influencia de los vientos alisios del NE, arrastran los sedimentos erosionados de la Isla de Salamanca y los depositan hacia la desembocadura del río Magdalena; otra parte de los sedimentos se desliza por cañones submarinos hacia el fondo del mar (Erffa, 1973).
La zona presenta dos tendencias estacionales: una época seca (diciembre-abril), regida por los vientos alisios del NE, y otra época lluviosa (mayo-noviembre), durante 3a cual la zona se encuentra influenciada por la descarga de aguas continentales y viento en calma, usualmente del S-SW, que se presenta en ausencia de los alisios (Blanco, 1988; Díaz, 1990). La precipitación total anual en la zona costera oscila entre 350 y 900 mm, en un gradiente en dirección SW-NE. La temperatura del aire promedio anual es de unos 28°C (Díaz, 1990).
Las mayores temperaturas superficiales del agua (alrededor de 29°C) se presentan en el período lluvioso de septiembre a noviembre. Durante los meses de diciembre a abril se presentan temperaturas menores (inferiores a 24°C), asociadas a la influencia de aguas de surgencia de salinidad elevada (Ramírez, 1990; Blanco et al, 1992).
MATERIALES Y MÉTODOS
En los meses de octubre y noviembre de 1991 se realizaron dos cruceros oceanográficos a bordo del B/I Ancón (INVEMAR, Santa Marta), durante los cuales se trabajaron cinco transectos lineales sistemáticos (A ,B, C, D y E), cada uno con estaciones ubicadas sobre las isóbatas de 15, 30, 60 y 100 m (figura 1). En cada estación se tomaron tres muestras del fondo con una draga Van Veen (cada muestra abarcando un área de 0.05 m2). El material extraído fue preservado con formalina al 5% en agua de mar previamente teñida con rosa de bengala, para permitir posteriormente una más ágil separación de los organismos del sedimento (Masón y Yevich, 1967).
Se estudió únicamente la macrofauna, definida como organismos retenidos en un tamiz con malla de 0.5 mm (Holme y Mclntyre, 1971; Stirn, 1981). El material fue extendido sobre una bandeja plástica para separar los organismos por taxa mayores, identificando hasta nivel de familia los anélidos y moluscos. Los resultados presentados se expresan con base a un área de 0.15 m2, que corresponde a la suma de los dragados tomados en cada estación.
Se escogieron los poliquetos, bivalvos y gastrópodos por conformar en conjunto más de la mitad del total de los organismos bentónicos (Longhurst y Pauly, 1987) y por tratarse de grupos poco vágiles, ligados en mayor o menor grado a determinada calidad textural del sedimento. Muchas de las comunidades del ben-tos de fondos blandos se nombran con base en estos grupos (Petersen, 1924; Thorsoh, 1957, citados por Gray, 1981). Se determinó el número total de familias por taxa,'así como el número de individuos por familia y por estación.
Para los análisis granulométricos se tomó una submuestra de cada dragado, la cual fue inicialmente secada a temperatura ambiente y posteriormente en estufa a 70°C hasta obtener peso constante. Las muestras constituidas por sedimentos fangosos fueron tamizadas en húmedo, aquellas conformadas por arenas se tamizaron en seco siguiendo la metodología recomendada por Holme y Mclntyre (1971), Folk (1980) y Stirn (1981). Tanto para el tamizado en húmedo como en seco se utilizó la batería de tamices que conforma la escala de Wentworth.
Se efectuaron dos análisis de agrupamiento: a) análisis normal (entre estaciones), que describe las asociaciones en el espacio, en el cual se redujo el número total de familias de 63 a 36, incluyendo sólo aquellas cuyo porcentaje de dominancia fue superior a 4 %, al menos en una estación, eliminando así las familias raras (Field et al., 1982) y disminuyendo el grado de distorsión interna de la técnica (Crisci y López, 1983). Se utilizó el índice de Bray-Curtis (con transformación de log (x+1)); b) análisis inverso (entre familias) que identifica las comunidades, trabajado a partir de los grupos formados en el análisis normal, seleccionándose aquellas familias cuyos porcentajes de dominancia por grupo fue mayor a 4%, reduciéndose el número de familias de 63 a 16. Esta matriz de datos fue doblemente estandarizada, una respecto a la abundancia total por estación (por columnas) y la otra por familias (por filas). La estandarización por columna responde a las grandes diferencias en la abundancia promedio de individuos entre los grupos, las cuales enmascaran la formación de los grupos. El índice utilizado (Bray-Curtis) es el mismo del análisis normal (Field et al., 1982; Jongman et al., 1987; Zupan, 1982). Se empleó la técnica del ligamiento promedio no ponderado (UPGMA) para la construcción de los dendrogramas (Matteucci y Colma, 1982; Crisci y López, 1983).
Una vez conformadas las asociaciones, con el fin de hacer comparaciones entre ellas, se utilizó el número promedio de individuos por familia dentro de cada grupo. Para cada una se determinaron los coeficientes de dominancia por familia, frecuencias, diversidad y uniformidad (Pielou, 1977; Odum, 1983; Margalef, 1986).
RESULTADOS
La figura 1 es una aproximación de los tipos de sedimentos presentes en el área de estudio. Para su elaboración se tomaron en cuenta, además de los resultados del presente trabajo, los de García y Sandoval (1983) y Molina (1990).
Básicamente se presentan cuatro tipos de sedimentos: cienos, arenas muy finas, arenas finas y arenas medias. Las arenas ocupan las estaciones de 15 y 30 m de profundidad y los cienos las de 60 y 100 m. Dentro de las arenas, las medias y finas se presentaron más cerca a la costa y las muy finas se encuentran entre las finas y los cienos. La excepción la constituyen las estaciones 1 y 2, que presentan cieno y están más cerca de la costa, debido quizás al transporte de sedimentos finos provenientes de la Ciénaga Grande de Santa Marta y depositados allí, como lo sugiere el patrón de circulación de corrientes en el sector (Blanco, 1988).
Dentro de los grupos analizados se encontró un total de 63 familias, 26 de las cuales corresponden a moluscos (18 bivalvos y 8 gastrópodos) y las restantes a los poliquetos. Estos últimos comprenden el 79.42% de los organismos aquí estudiados, el 20.58% restante se distribuye en 19.91% para bivalvos y 0.67% para gastrópodos.
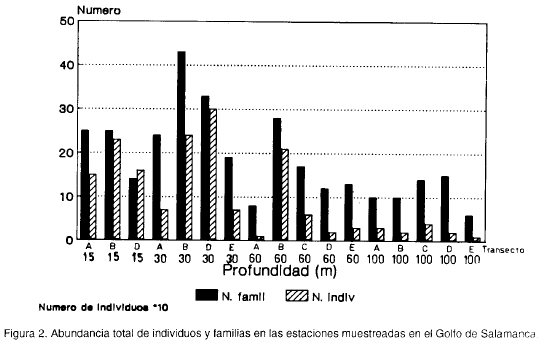
Las mayores abundancias por familias e individuos se presentaron en las estaciones de 15 y 30 m de profundidad (figura 2). El 55.74 % de la abundancia total de individuos se encontró en cuatro estaciones, dos de ellas ubicadas en el transecto B y las otras dos en el D, en las profundidades de 15 y 30 m. Las menores abundancias de individuos y familias se encontraron en general en las estaciones de 60 y 100 m de profundidad, exceptuando la estación de 60 m del transecto B.
La figura 3 muestra la asociación entre estaciones. Se formaron tres grupos, dos más similares entre sí, A y B, respecto al tercero, C; el grupo A está conformado por las estaciones 2, 5, 6, 7, 11, 12, 13 v 15, el grupo B por las estaciones 3, 9 y 16, y el C por las estaciones 1, 4, 8 y 14.
La figura 4 muestra la agrupación entre las familias; un grupo lo conforman Glyceridae, Syllidae, Terebellidae, Magelonidae, Lumbrineridae, Onuphiidae, Capitellidae, Orbiniidae y Cirratulidae; el segundo Ampharetidae, Lucinidao, Paraonidae y Sabellidae; el tercero, Maldanidae, Corbulidae y Nuculanidae.
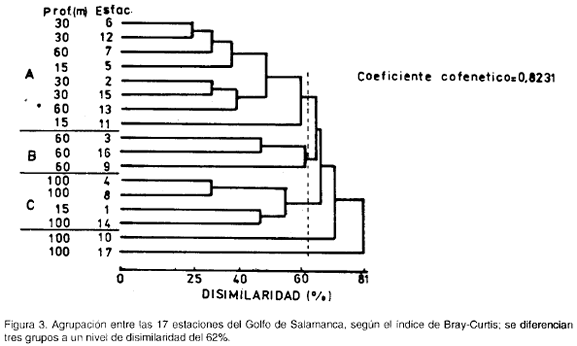
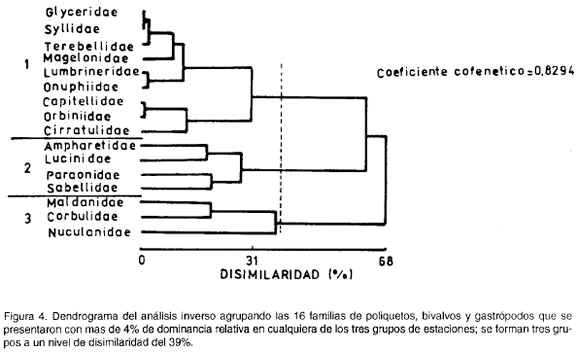
Para poder relacionar los dos dendrogramas, se analizaron las dominancias relativas al total por grupo de cada una de las familias seleccionadas en el análisis inverso, obteniéndose el siguiente esquema: el segundo grupo de familias es el más dominante en el grupo A de estaciones, el primero de familias en el B de estaciones y el tercero de familias en el C de estaciones.
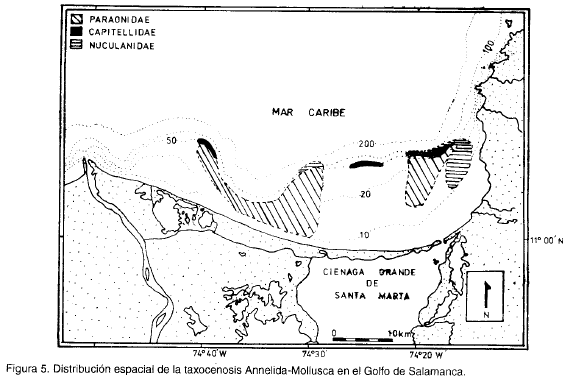
Teniendo en cuenta el tipo de sedimento, la profundidad y los grupos anteriormente mencionados, se obtuvo la figura 5, la cual muestra la distribución espacial de la taxocenosis. Básicamente se trata de tres asociaciones. La primera, denominada Paraonidae, es la más cercana a la costa, asentada en fondos heterogéneos conformados por cieno y arenas (medias, finas y muy finas), caracterizada por las familias Ampharetidae, Lucinidae, Paraonidae y Sabellidae. La segunda, denominada Capitellidae, está conformada por las familias Glyceridae, Syllidae, Terebellidae, Magelonidae, Lumbrineridae, Onuphiidae, Capitellidae, Orbiniidae y Cirratulidae, alrededor de la isóbata de 60 m, en sedimentos finos tipo cieno. La tercera, llamada Nuculanidae, representada por Maldanidae, Corbulidae y Nuculanidae, está asentada en sedimentos muy finos alrededor de los 100 m de profundidad.
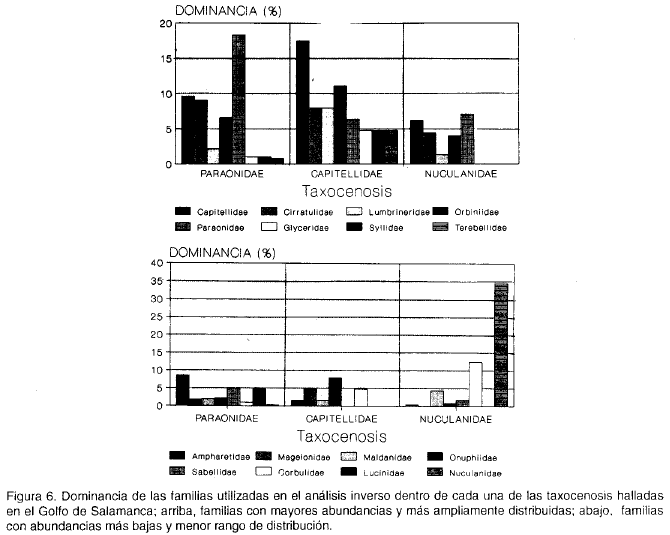
El comportamiento de la dominancia por familias es diferente en cada una de las asociaciones; es así como 9 de las 16 familias más dominantes se encuentran mejor representadas en la taxocenosis Capitellidae, siendo la familia más dominante Capitellidae (figura 6); la taxocenosis Nuculanidae se caracteriza por presentar en general las menores dominancias y el menor número de familias; aunque en la taxocenosis Paraonidae se encuentran todas las familias representadas en la figura 7, éstas muestran en general valores intermedios de dominancia respecto a las otras dos biocenosis, destacándose Paraonidae como la más dominante en esta asociación.

La diversidad es mayor en la taxocenosis Paraonidae y va disminuyendo hacia las comunidades más profundas, presentándose su menor valor en la biocenosis Nuculanidae. La uniformidad presenta un comportamiento inverso a la diversidad (tabla 1).
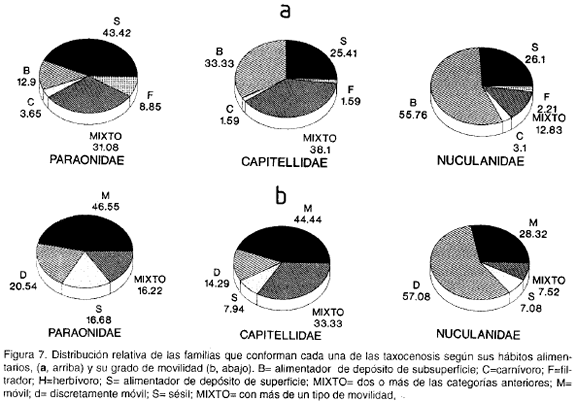
De acuerdo con las categorías alimentarias y de movilidad para los organismos bentónicos propuestas por Jumars y Fauchald (1977), Fauchald y Jumars (1979) y Morton (1983), las taxocenosis muestran algunas particularidades. Los elementos filtradores (F) son más abundantes en la comunidad Paraonidae que en las otras dos; por otro lado, los alimentadores de depósito de superficie (S) son reemplazados por los alimentadores de depósito de subsuperficie (B) en las taxocenosis más profundas (Capitellidae y Nuculanidae) (figura 7a). De acuerdo con las categorías de movilidad, dominan los móviles (M) en las taxocenosis Paraonidae y Capitellidae, los cuales son reemplazados por los discretamente móviles (D) en la comunidad Nuculanidae; los sésiles (S) son más abundantes en la biocenosis Paraonidae que en las otras dos, mientras que los de hábitos mixtos (M-D-S) son poco dominantes en la taxocenosis Nuculanidae en comparación con las otras dos asociaciones (figura 7b).
DISCUSIÓN Y CONCLUSIONES
La identificación de tres tipos de taxocenosis o asociaciones Mollusca-Annelida en el bentos de fondos blandos en el área estudiada a partir de los análisis de agrupamiento normal e inverso aplicados al nivel sistemático de familia, guarda relación con los gradientes de calidad textural del sedimento y de profundidad. Las investigaciones realizadas por Warwick (1988a,b) y Ferraro y Colé (1990) demuestran que un análisis de la información faunística a nivel de familia presenta el mismo grado de resolución, y los resultados son tan confiables, como cuando se hace a nivel de género o de especie para la determinación de los esquemas de distribución de comunidades bentónicas con respecto a gradientes ambientales. Esto representa un ahorro en tiempo y costos, ya que la identificación a nivel de géneros o especies requiere de especialistas en los diferentes grupos zoológicos y más tiempo. En regiones tropicales, la semejanza entre los análisis de familias y los de géneros y especies no ha sido corroborada; sin embargo, la comparación preliminar de los resultados aquí presentados con los hechos a nivel de géneros (información que será publicada en un futuro próximo) llevan a esquemas de distribución muy parecidos.
La relación observada entre la mayor abundancia y diversidad de la comunidad Paraonidae y las arenas, puede obedecer a la capacidad que tienen éstas para retener una mayor cantidad de agua rica en materia orgánica, la cual puede ser fácilmente aprovechada por los organismos. De otra parte, la arena ofrece un sustrato más estable en comparación con los cienos para el establecimiento de comunidades del endobentos (Lastra et al., 1991; Rhoads y Young, 1970).
El tipo de sedimento determina la porosidad y capilaridad del medio (Theede, 1981; Méndez et al., 1986). En los cienos y lodos, la porosidad y capilaridad es menor que en las arenas, por lo que en ellos se retiene una menor cantidad de agua rica en materia orgánica, la cual constituye una importante fuente de alimentó (De la Lanza, 1986; Méndez et al., 1986). Este hecho contribuye a dar explicación a los bajos valores de abundancia y diversidad encontrados en las biocenosis más profundas, asentadas básicamente en cienos.
La disminución de la diversidad en las comunidades más profundas es el reflejo de condiciones más exigentes. Rhoads y Young (1970), Rhoads (1974), Brenchley (1981,1982), Wilson (1991) y Lastra et al. (1991) anotan que la escasez de organismos alimentadores de depósito de superficie y filtradores puede relacionarse con la hipótesis de "amensalismo de grupo trófico", según la cual la modificación del medio bentónico por los alimentadores de depósito de subsuperficie (bioturbación), provoca la exclusión de muchos alimentadores de depósito de superficie y epifauna sésil. La bioturbación produce inestabilidad en el medio, ocasionando el "enterramiento" de las larvas asentadas de organismos alimentadores de depósito de superficie y epifauna sésil; además, la acción de los bioturbadores crea condiciones poco propicias para el asentamiento de nuevas larvas de estas categorías alimentarias y obstruye las estructuras de los organismos alimentadores de superficie.
La profundidad, además de influir sobre la abundancia, juega un papel importante en las características estructurales de los organismos. Es así como las zonas profundas estables con poco alimento presentan organismos frágiles, alargados o con apéndices alimenticios extensibles, mientras que aquellos que viven en áreas más someras y con un buen flujo de alimento tienden a ser más robustos (Jumars y Fauchald, 1977). Además, si se tienen en cuenta las categorías de los hábitos alimenticios, los resultados indican que la forma de captar el alimento tiende a cambiar con la profundidad, dominando los alimentadores de depósito de superficie en las áreas más someras (Paraonidae) y los de depósito de subsuperficie en la comunidad más profunda (Nuculanidae).
Teniendo en cuenta los tipos de movilidad, las familias que conforman la comunidad más profunda y que viven en fondos de cieno son ligeramente móviles en su mayoría. Según Rhoads (1974) y Brenchley (1982), el grado de movilidad se relaciona en general con el tipo de sedimento, siendo reducido en sedimentos finos cementados (donde los espacios entre los granos son llenados por partículas finas), mientras que en las arenas (donde tales espacios son llenados con agua) el sustrato propicia un grado de movilidad mayor. En el caso de la biocenosis Paraonidae, asentada en fondos primordialmente arenosos, los organismos dominantes tienen un relativo alto grado de movilidad.
La diversidad más alta se presentó en la taxocenosis Paraonidae, dentro de la cual se encuentra la mayor abundancia de individuos sésiles (fig. 8b). Jumars y Fauchald (1977) determinaron que la heterogeneidad estructural se relaciona positivamente con la diversidad y que ésta es máxima donde dominan los organismos sésiles. Ello obedecería a que las áreas dominadas por organismos sésiles son físicamente poco perturbadas, biológicamente no son rápidamente homogenizadas y la bioturbación es poca.
La comunidad más rica en cuanto a número de individuos y familias (Paraonidae) se encuentra en cercanías del área conocida como "Banco de las Animas", a unos 11 km al norte de la costa de la Isla de Salamanca, entre los paralelos 74°24' y 74°29' W (Blanco et al, 1992); esta zona constituye posiblemente un área de alimentación para los peces, como lo sugiere la gran abundancia y riqueza íctica halladas allí.
En la mitad oriental de esta misma área de la plataforma continental, García y Sandoval (1983) describieron dos biocenosis bentónicas, denominadas por ellos Nucula y Solen (ambos Bivalvia), que se extendían entre los 5 y 36 m de profundidad; comparando los resultados del presente trabajo con el realizado casi una década antes, cuya área común se encuentra representada por los transectos A y B (profundidades de 15 y 30 m), es evidente que las comunidades han sido remplazadas parcialmente: la comunidad Paraonidae ha desplazado parcialmente a las dos biocenosis asentadas en 1982 y posee, aunque en abundancias muy bajas, elementos de la comunidad Nucula, que, sin embargo, siguen siendo exclusivos de ese sector; la taxocenosis Nuculanidae desplazó totalmente a la comunidad Nucula, a pesar de que aún es dominada por bivalvos con las mismas características biológicas (captura de alimento y movilidad) que en aquella. Puesto que se trata de estudios realizados en dos épocas climáticas diferentes (abril de 1982, octubre-noviembre de 1991), ignoramos si se trata quizás de 2 fases temporales de una ciclocenosis (en el sentido de Schwerdtfeger, 1975), es decir, de comunidades que a lo largo del año cambian en su composición retornando siempre al mismo esquema, o efectivamente de un remplazo de comunidades ocasionado por alguna perturbación ambiental. Lo que sí parece prevalecer con el tiempo es el concepto de comunidades continuas o casi continuas, como fue descrito por García y Sandoval (1983) para esta área y por Díaz y Gotting (1986) para las taxocenosis de moluscos en la Bahía de Neguange, que implica el que una comunidad se va "diluyendo" en otra y que las taxa más dominantes se presentan en número variable en las demás biocenosis (Gray, 1981).
AGRADECIMIENTOS
El Instituto Colombiano de Investigaciones Científicas y Proyectos Especiales "Francisco José de Caldas" COLCIENCIAS aportó los medios para la financiación del proyecto "Reconocimiento piloto de fondos, ambiente, fauna asociada y recursos pesqueros en aguas costeras del Departamento del Magdalena", en marco del cual se realizó la presente investigación. El Instituto de Investigaciones Marinas de Punta de Betín - INVEMAR - facilitó la infraestructura requerida. Los miembros de la tripulación del B/I "Ancón" y demás participantes en los cruceros prestaron su valiosa colaboración en los muestreos. Oscar D. Solano y Jacobo A. Blanco prestaron invaluable apoyo y contribuyeron con acertadas sugerencias a la realización del presente trabajo. Los doctores Sven Zea y Camilo García pusieron a nuestra disposición valiosa bibliografía. Este trabajo forma parte de la tesis del primer autor para optar al grado de Magister Scientiae, especialidad en Biología Marina, de la Universidad Nacional de Colombia-INVEMAR.
BIBLIOGRAFÍA
1 Alongi, D. 1989. Ecology of tropical soft-bottom benthos: a review with emphasis on emerging concepts. Rev. Biol. Trop., 37(1): 85-100. [ Links ]
2 Blanco, J.A. 1988. Las variaciones ambientales estacionales en las aguas costeras y su importancia para la pesca en la región de Santa Marta, Caribe Colombiano. Tesis M.Sc. Biol. Mar. Universidad Nacional de Colombia-INVEMAR. Santa Marta, 50 p. [ Links ]
3 Blanco, J.A., J.M. Díaz, G. Ramírez y M.L Cortés. 1992. El Banco de las Animas: una amplia área arrecifal localizada sobre un antiguo delta del Río Magdalena. Resúmenes del VII Seminario de las Ciencias y las Tecnologías del Mar y Congreso Centroamericano y del Caribe en Ciencias del Mar, Comisión Colombiana de Oceanografía. Santa Marta. 40 p. [ Links ]
4 Brenchley, G. 1981. Disturbance and community structure: an experimental study of bioturbation in marine soft-bottom environments. J. Mar. Res., 39(4): 767-789. [ Links ]
5 Brenchley, G. 1982. Mechanisms of spatial competition in marine soft-bottom communities. J. Exp. Mar Biol. Ecol., 60: 17-33. [ Links ]
6 CORPES, 1992. El Caribe Colombiano: realidad ambiental y desarrollo. Consejo Regional de Planificación de la Costa Atlántica, Bogotá, 350 p. [ Links ]
7 Crisci, J. y M. López. 1983. Introducción a la teoría y práctica de la taxonomía numérica. OEA., monografía No 26. Washington D.C., 132 p. [ Links ]
8 De la Lanza, G. 1986. Materia orgánica en los sedimentos del sistema lagunar de Huizache y Caimanero: importancia, comportamiento y significado de los modelos de predicción. Ann. Inst. Cienc, del Mar y Limnol. Univ. Nal. Auton. Mexico, 13(1): 251-286. [ Links ]
9 Díaz, J. (ed). 1990. Estudio ecológico integrado de la zona costera de Santa Marta y Parque Nacional Natural Tayrona. INVEMAR. Informe final. Santa Marta, 440 p. [ Links ]
10 Díaz, J. y K. Gotting. 1986. Mollusken-Gemeinschaften der Bahía de Nenguange (Karibik, Kolumbien). Helgoländer Meeresunters, 40: 279-308. [ Links ]
11 Erffa, A. von. 1973. Sedimentation, Transport und Erosion an der Nordküste Kolumbiens zwischen Barranquilla und der Sierra Nevada de Santa Marta. Mitt. Inst. Colombo-Alemán Invest. Cient., 7: 155-209. [ Links ]
12 Ferraro, S. y F. Cole. 1990. Taxonomic level and sample size sufficient for assessing pollution impacts on the Southern California Bight macrobenthos. Mar. Ecol. Prog. Ser., 67:251-262. [ Links ]
13 Field, J., K. Clark y R. Warwick. 1982. A practical strategy for analysing multispecies distribution patterns. Mar. Ecol. Prog. Ser., 8: 37-52. [ Links ]
14 Folk, R. L. 1980. Petrology of sedimentary rocks. Hemphill Publishing Company, Austin, Texas, 182p. [ Links ]
15 Fauchald, K. and P. Jumars. 1979. The diet of worms: a study of polychaete feeding guilds. Oceanogr. Mar. Biol. Ann. Rev., 17, 193-284. [ Links ]
16 García, C. y H. Sandoval. 1983. Comunidades macrozoobénticas de fondos blandos en la Plataforma Continental de Ciénaga, Caribe Colombiano. Tesis de grado, Facultad Biol. Mar., Univ. Jorge Tadeo Lozano. Bogotá, 84 p. [ Links ]
17 García, C. y H. Salzwedel. 1991. Structure of soft-bottom macrobenthos in shallow areas off the Caribbean coast of Colombia: introducing a new strategy, 239-249, In: Elliot, M. y J. Ducrotoy (eds). Estuaries and coasts: spatial and temporal intercomparisons. ECSA 19 Symposium. Olsen y Olsen. Bremerhaven, Alemania. [ Links ]
18 Gray, J. 1981. The ecology of marine sediments (Cambridge Studies in modern Biology, 2). Cambridge Univ. Press. Syndicate, Cambridge, U.S.A., 193 p. [ Links ]
19 Holme, N. y A. Mclntyre. 1971. Methods for the study of marine benthos. Blackwell Scientific Publications. IBP Handbook No. 16, Oxford, 334 p. [ Links ]
20 Jogman, R., C. ter Braak y O. Van Tongeren. 1987. Data analysis in community and landscape ecology. Pudoc Wageningen, Wageningen, Holanda, 299 p. [ Links ]
21 Jumars P. and K. Fauchald. 1977. Feeding and metabolism, pp. 1-20, in: Could, B. C. (ed.). Ecology of marine benthos. Univ. South Carolina Press, Columbia, South Carolina. [ Links ]
22 Lastra, M., J. Palacio, A. Sánchez y J. Mora. 1991. Estructura trófica infralitoral de la Bahía de Santander. Cah. Biol. Mar., 32: 33-351. [ Links ]
23 Longhurst, A. y D. Pauly. 1987. Ecology of tropical oceans. Academic Press Inc., San Diego, 407p. [ Links ]
24 López, A. 1981. Estudio ecológico comparativo de la fauna macrobentónica de las Bahías de Colombia y Grande del Golfo de Urabá. Tesis de grado, Facultad Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá, 45 p. [ Links ]
25 Lorin, J., C. Hernández, A. Rovault y J. Bottagisio. 1973. Estudio sedimentológico de la plataforma continental entre Bocas de Ceniza y Santa Marta. Laboratoire Central D'Hydraulique de France, Barranquea, 42 p. + 50 planos. [ Links ]
26 Margalef, R. 1986. Ecología. Ediciones Omega S. A., Barcelona, 951 p. [ Links ]
27 Mason, W. and P. Yevich. 1967. The use of Phloxine B and rose bengal stains to facilite sorting benthic samples. Trans. Am. Microsc. Soc, 86(2):221-223. [ Links ]
28 Matteucci, S. y A. Coima. 1982. Metodología para el estudio de la vegetación. OEA. Monografía No 22, Washington D.C., 168 p. [ Links ]
29 Méndez, M., V. Solis-Weiss y A. Carranza-Edwards. 1986. La importancia de la granulometría en la distribución de organismos bentónicos. Estudio de playas del estado de Veracruz, México. Ann. Inst. Cienc. del Mar y Limnol. Univ. Nal Auton. México, 13(3): 45-56. [ Links ]
30 Molina, A. 1990. Estudio geológico de la plataforma continental caribe (Santa Marta-Punta Morro Hermoso). Mem. VII Semin. Nac. Cienc. Tecnof. Mar, Comisión Colombiana de Oceanografía, Cali, 120-131 p. [ Links ]
31 Mora, J. y G. Prieto. 1978. Estudio preliminar del macrobentos y sus condiciones medio ambientales en la bahía de Cartagena. Tesis de grado, Facultad Biol. Mar., Univ. Jorge Tadeo Lozano, Cartagena, 112 p. [ Links ]
32 Morton, B. 1983. Feeding and digestion in Bivalvia, pp. 65-147 in: Soleudding, A. y K. Wilbur (eds), The Mollusca, Vol. 5 (Physiology). Academic Press, New York. [ Links ]
33 Odum, E. 1983. Ecología. Tercera edición. Nueva Editorial Interamericana, México, 639 p. [ Links ]
34 Pielou, E. 1977. Mathematical ecology. John Wiley & Sons, New York, 385 p. [ Links ]
35 Puentes, L., N. Campos y R. Reyes. 1990. Decápodos de fondos blandos hallados en el área comprendida entre Pozos Colorados y la Bahía de Taganga, Caribe Colombiano. Bol. Ecotrópica, 23: 31-41. [ Links ]
36 Ramírez, M. 1983. Contribución al conocimiento de los ostrácodos (Arthropoda-Crustácea) actuales de la Plataforma Continental frente a Riohacha y su relación con factores del medio ambiente. Tesis de grado, Facultad Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá, 135 p. [ Links ]
37 Ramírez, G. 1990. Evaluación de parámetros hidrográficos y su relación con la surgencia en las aguas costeras, 19-54, In: Díaz, J. (ed). Estudio ecológico integrado de la zona costera de Santa Marta y Parque Nacional Natural Tayrona. INVEMAR. Informe final, Santa Marta, 440 p. [ Links ]
38 Rhoads, D. 1974. Organism-sediment relations on the muddy sea floor. Oceanogr. Mar. Biol. Ann. Rev., 12, 263-300. [ Links ]
39 Rhoads, D. y D. Young. 1970. The influence of deposit-feeding organisms on sediment stability and community trophic structure. J. Mar. Res., 28(2): 150-178. [ Links ]
40 Rodríguez, N. 1983. Micromoluscos recientes de la Plataforma Caribe Colombiana entre Bocas de Ceniza e Isla Fuerte. Tesis de grado, Facultad Biol. Mar., Universidad Jorge Tadeo Lozano, Bogotá, 247 p. [ Links ]
41 Schwerdtfeger, F. 1975. Synoekologie. Paul Parey, Hamburg, 451 p. [ Links ]
42 Stirn, J. 1981. Manual of methods in aquatic environment research. Part 8, Ecological assesment of pollution effects. FAO Fish. Tech. Pap., 209: 70 p. [ Links ]
43 Theede, J. 1981. Studies on the role of benthic animals of the western Baltic in the flow of energy and organic material. K'teler Meeresforsch. Sonderh., 5: 434-444. [ Links ]
44 Warwick, R. 1988a. Analysis of community attributes of the macrobenthos of Frierfjord/Langesundfjord attaxonomic levels higher than species. Mar. Ecol. Prog. Ser., 46:167-170. [ Links ]
45 Warwick, R. 1988b. The level of taxonomic discrimination required to detect pollution effects on marine benthic communities. Mar, Poll. Bull., 19(6):259-268. [ Links ]
46 Wilson, W. 1991. Competition and predation in marine soft-sediment communities. Ann. Rev. Ecol. Syst., 21: 221-241. [ Links ]
47 Zupan, J. 1982. Clustering of large data sets. Research Studies Press, Chinchester, 122 p. [ Links ]
