










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.23 no.1 Santa Marta Jan./Dec. 1994
DETERMINACIÓN COMPARATIVA DE LA PRODUCCIÓN PRIMARIA CON LAS TÉCNICAS DE 14C Y O2 EN AGUAS SALOBRES CON DIFERENTES GRADOS DE EUTROFICACIÓN
Klaus Gocke1 y Carlos A. Hernández J.2
1Institut für Meereskunde an der Universität Kiel, Düsternbrooker Weg 20, D-24105 Kiel, Alemania (K.G.).
2Universidad de Jorge Tadeo Lozano, Facultad de Biología Marina, A. Aéreo 842, Santa Marta, Colombia (C.H.J.).
RESUMEN
Se estudió la producción primaria del fitoplancton comparando las técnicas de la fijación de 14CO2 y la liberación de O2 en la Bahía de Kiel (BK, mesotrófico), el Fiordo de Kiel (FK, eutrófico) y el Fiordo de Schlei (hipertrófico). Estos lugares salobres de la parte más occidental del Mar Báltico se distinguen marcadamente en su producción primaria (PP). En promedio y considerando solamente la profundidad de máxima PP, la producción del Schlei por unidad de volumen excedió 9 veces la del Fiordo de Kiel y más de 40 veces la de la Bahía de Kiel. Por unidad de área las diferencias fueron mucho menos pronunciadas, dado que la profundidad de compensación se encontró a los 0.8 m (BK: 5.5 m, FK: 5.2 m). De acuerdo con consideraciones teóricas se encontró que la PP determinada con la técnica de 14C (14C-PP) fue mayor que la producción neta (PN) y menor que la producción bruta (PB), ambas determinadas con el método de O2 empleando un cociente fotosintético (CF) de 1.2. La PN promedio del Fiordo de Kiel fue 9 % menor y la PB 36 % mayor que la 14C-PP, en la Bahía de Kiel la PN fue 40 % menor y la PB 72 % mayor. Sin embargo, las relaciones PN<14C-PP<PB se observaron solamente en el Fiordo y la Bahía de Kiel. Al contrario en el poliproductivo Fiordo de Schlei se encontró que la 14C-PP fue mayor que la PN y PB (32 % mayor que la PN y 7 % mayor que la PB). Esta discordancia con la teoría se explica principalmente por el hecho que el fitoplancton se encontró en un estado senescente, lo que resultaría en un CF menor de 1.2.
ABSTRACT
The primary productivity of phytoplankton was determined in the mesotrophic Kiel Bight (KB), the eutrophic Kiel Fjord (KF) and the hypertrophic Schlei Fjord using the 14C-technique and the O2-method. Results of these measurements were compared. These brackish areas of the western-most part of the Baltic Sea differ markedly in primary productivity (PP). If the depth of maximum productivity alone is considered, the PP, on a volumetric basis, of the Schlei was 9 times higher than in the Kiel Fjord and more than 40 times that of the Kiel Bight. On an area basis the differences were much smaller since the compensation depth in the Schlei was found at no more than 0.8 m, whereas in the Kiel Bight it was 5.5 m and 5.2 m in the Kiel Fjord. In agreement with theoretical considerations, the PP determined with the 14C-technique (14C-PP) was larger than the net productivity (NP) and smaller than the gross productivity (GP), both determined with the O2-method using a photosynthetic quotient of 1.2. On average, in the Kiel Fjord the NP was 9 % smaller and the GP 36 % larger than the 14C-PP. in the Kiel Bight the NP was 40 % smaller and the GP 72 % larger. However, the relation NP<14CPP<GP was found only in the Kiel Bight and -Fjord, whereas in the polyproductive Schlei the 14C-PP was larger than both the NP and GP (32 % larger than the NP and 7 % larger than the GP). This deviation from theoretical values may be due, in part, to a senescent phytoplankton, which may have had a PQ smaller than 1.2.
INTRODUCCIÓN
La producción primaria (PP) es probablemente el parámetro de función más frecuentemente determinado tanto en los biótopos marinos como limnológicos. Existen varias posibilidades para su estudio, las técnicas más usadas consisten en la determinación del oxígeno liberado y la incorporación de carbono radioactivo.
El primer método fue introducido por Gaarder y Gran (1927) y es conocido usualmente como el "método de O2" o el "método de botellas claras y oscuras". La técnica tiene como ventaja, principalmente una ejecución simple, ya que fuera del equipo para la incubación de las botellas, solamente requiere los mismos aparatos y reactivos que se necesitan para la determinación de la concentración de oxígeno disuelto. Además permite la determinación de la producción primaria neta (PN), la producción primaria bruta (PB) y la respiración de la comunidad planctónica (R). Hay que aclarar que con respecto a la respiración se hace una simplificación ya que no se toma en cuenta una posible fotorespiración, más bien se considera la respiración de las algas en la oscuridad como si fuera idéntica con su respiración en la luz. Esta simplificación podría ser incorrecta en algunos casos. En la mayoría de ellos, por lo menos en el ambiente acuático, debería ser igual o el error que se comete debería ser muy pequeño (Parsons et al., 1984). Una de las desventajas más grandes del método de O2, la que limita bastante su aplicación, es su reducida sensibilidad. Tanto en mar abierto como en zonas costeras y aguas continentales oligotróficas las diferencias entre las concentraciones de oxígeno de las botellas claras, oscuras e iniciales (incluso después de una incubación prolongada durante todo el día) son tan pequeñas que el método usual de la determinación de O2 no permite determinarlas con suficiente exactitud (Vollenweider, 1974). Según este autor la producción primaria más baja detectable corresponde a alrededor de 10 mg Cm-3d-1 Bryan et al. (1976) mejoraron el método considerablemente aplicando una detección fotométrica del punto final de la titulación. Los autores opinan que su versión tiene suficiente sensibilidad para permitir la determinación de la producción primaria incluso en biótopos oligotróficos donde la PP no sobrepasa los 2-3 mg Cm-3d-1 (P.J.LeB. Williams, comunicación personal). Otra desventaja es el problema de convertir las cantidades de oxígeno liberado en cantidades de carbono fijado. Para hacer esto se aplica normalmente un cociente fotosintético (CF) de 1.2 o sea una incorporación de 1000 moléculas de CO2 lleva consigo una liberación de 1200 moléculas de O2 (Parsons et al., 1984; Margalef, 1986). El CF de 1.2 toma en consideración que la biomasa de las algas es más reducida que glucosa, lo que significa que el CF no puede ser 1 como en la ecuación de la fotosíntesis sino tiene que ser más alto. El cociente fotosintético entonces depende de la composición de la biomasa de los organismos fotosintéticos y de la fuente de nitrógeno. Una porción alta de grasas y la utilización de NO3 resulta en un valor alto del CF (Davies y Williams, 1984; Raine, 1983; Williams et al., 1979).
El método para la determinación de la producción primaria que hoy en día se aplica más que cualquier otro, es el que introdujo Steemann-Nielsen (1952) en la ecología acuática. Se conoce mejor bajo el nombre de "método de 14C" ya que se basa en la utilización de NaH14CO3, El método tiene como ventaja su alta sensibilidad, lo que permite su uso incluso en aguas ultra oligotróficas (PP<1 mg Cm-3d-1; Strickland, 1960). Además se puede llevar a cabo una diferenciación de los productores primarios por clase de tamaño y se puede determinar la exudación de materia orgánica disuelta por el fitoplancton. Otra ventaja es que no requiere un factor de conversión ya que se mide directamente la formación de carbono orgánico. Desventajas son los altos costos de cada experimento y del sofisticado equipo necesario. Una desventaja importante consiste en que no se obtiene ni la producción neta ni la bruta sino un valor entre ambas (Raymont, 1980; Margalef, 1986, Irwin, 1991), dependiendo de la duración de la incubación el resultado corresponde más a la PN o PB.
Ya que el conocimiento de la magnitud de la producción primaria es un prerrequisito para poder entender la dinámica del ciclo de material en cualquier ecosistema, hubo muchos esfuerzos desde la introducción del método de 14C para determinar si los resultados de este método son comparables a los de la técnica de O2 más antigua (Peterson, 1980; Williams et al., 1983; Grande et al., 1989). Esto también ha sido el objetivo del presente estudio, con el cual se pretendió en 3 áreas muy diferentes hacer tal comparación con respecto a su productividad. Se aplicó exactamente la misma versión de los 2 métodos para ver si el grado trófico del sistema influye en el resultado.
ÁREA DE ESTUDIO
Fiordo de Kíel. Es una extensión de la parte oeste de la Bahía de Kiel adyacente a la ciudad de Kiel (Fig. 1). Tiene una longitud de 17 km. Su parte interior está casi separada de la parte exterior por una angostura de 1 km. Las aguas son salobres con una variación de salinidad marcada durante el año. Recibe un río pequeño, el cual tiene poca importancia para la salinidad, ya que ésta depende totalmente de las variaciones de la Bahía de Kiel. Kándler (1959) y Lenz (1977) describieron detalladamente la topografía e hidrografía del fiordo. La parte interna es eutrófica Según Moigis (1983) la producción primaria (determinada con el método de O2) es de 370 g Cm-3d-1.
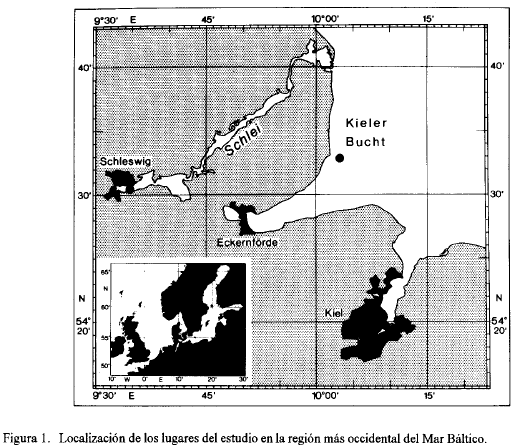
Bahía de Kiel. Representa el área más occidental del Mar Báltico. Es una región de transición entre el Mar del Norte y el Mar Báltico (Fig. 1). Por tal razón está sometida a cambios rápidos y fuertes de salinidad. Varía entre 10 y 20 %o (valores extremos, capa superficial) y 20 y 30 %o en la capa profunda debajo de 25 m (Lenz, 1977). Los valores altos se observan durante el invierno. La Bahía de Kiel es mesotrófica, se ha reportado una producción primaria anual (determinada con el método de 14C) de 158 g C (Bodungen,1975).
Fiordo de Schlei. Se encuentra en un canal glacial inundado conectado con la Bahía de Kiel (Fig. 1). Se parece mucho a un río y tiene una extensión de alrededor de 40 km. Solamente las partes más internas tienen un carácter lagunar. En esta parte del Schlei la salinidad varía durante el año entre 3 y 9 %o. Debido a su reducida profundidad (alrededor de 4 m en la estación de estudio) no existe estratificación de la columna de agua o es de corta duración (Nellen y Rheinheimer, 1970). El Schlei recibe aguas negras tratadas y además efluentes de los terrenos fertilizados. El aporte de nutrientes junto con sus condiciones hidrográficas causa una hipertrofia notable. Según Schiemann (1974) la producción primaria es de 850 g Cm-3d-1 (determinado con el método de O2) en la parte interna del sistema.
Una descripción comparativa del Fiordo de Kiel y del Fiordo de Schlei sobre el grado de trofia y su influencia sobre el microplancton autótrofo y heterótrofo es dada por Gocke y Rheinheimer (1991).
MATERIALES Y MÉTODOS
Muestreo e Incubación. Se realizaron tres series de incubaciones en el Fiordo de Kiel, dos en la Bahía de Kiel y dos en el Fiordo de Schlei entre el 11 de agosto y el 3 de septiembre de 1992. Se determinó la PP a 5 profundidades de luz para obtener su distribución vertical y para poder calcular la producción por unidad de área. En la tabla se presentan las profundidades a las cuales se obtuvieron las muestras para cada profundidad de incubación. Para el primer experimento en el Fiordo de Kiel (11.8.) y para los dos experimentos en la Bahía de Kiel, se tomó una sola muestra (a 2 m), con la cual se llenaron los frascos correspondientes a las tres profundidades de incubación superiores debido a que éstas se encontraron en una capa homogénea, ahorrando así trabajo y material. Las otras 2 muestras se tomaron a las profundidades en las cuales luego fueron incubados los frascos.
Debido a que en el Fiordo de Schlei se encontró una capa eufótica homogénea, se tomó una sola muestra a 0.5 m con la cual se llenaron los frascos para todas las profundidades de incubación. Para el segundo y tercer experimento en el Fiordo de Kiel (17.8. y 24.8.) también se llenaron los frascos de la cinco profundidades de incubación con una sola muestra (a 2 m) a pesar de que hubo una estratificación, ya que con estos experimentos solamente se pretendió estudiar el efecto de diferentes intensidades de luz sobre la misma población (abundancia y composición) de fitoplancton. A cada profundidad de incubación se colocaron un par de botellas de vidrio transparente de 100 ml (volumen nominal) para el método de O2 y un par de frascos de policarbonato transparente de 30 ml para el método de 14C, procurando que los recipientes no se dieran sombra. Las botellas y los frascos oscuros acompañantes (2 para cada profundidad en el caso del método de O2 y 1 para cada profundidad para el método de 14C) se incubaron en tubos de PVC oscuro a temperatura ambiental. La duración de las incubaciones se indica en la tabla.
Método de O2. Se empleó el método de Winkler para la determinación del oxígeno usando la versión de Green y Carrit (1966). Se tituló todo el contenido de las botellas con tiosulfato de sodio 0.O2 N y se utilizó como indicador almidón. La titulación se realizó con una bureta semi-automática (Schott, T 90/20) a más tardar 1 día después de la fijación del O2. El error de titulación (diferencia media entre las replicas, todas las muestras combinadas) fue solamente 0.15 % en el caso de la concentración inicial, 0.20 % en las botellas oscuras y 0.25 % en las botellas claras. El error es poco mayor al que se consigue con las versiones sofisticadas de la determinación de oxígeno (0.05 - 0.1 %), las cuales emplean una detección fotométrica o amperométrica del punto final (Bryan et al., 1976; Williams y Jenkinson, 1982; Bell y Kuparinen, 1984). Para la conversión de masa de O2 en masa de carbono orgánico se aplicó un cociente fotosintético de 1.2 y un cociente respiratorio de 0.84, lo que significa que 1 mg de O2 liberado o consumido es igual a 0.313 mg de carbono producido o respirado.
Método de 14C. Se adicionaron 4 μ Ci de NaH14CO3 (Amersham Buchler) a los frascos a incubar. Inmediatamente transcurrida la incubación se filtró todo el contenido de los frascos (Fiordo y Bahía de Kiel) o solamente 5 ml (Schlei) por filtros de membrana de 0.2 μ (Sartorius, acetato de celulosa). Los filtros se expusieron al vapor de HCl por 15 minutos en un desecador para eliminar el 14CO2 no incorporado. Después se determinó la radioactividad a través de un contador de centelleo líquido (Tri Carb, Packard). El "quench" se determinó con el método de "extemal-standard-channel-ratio", obteniéndose una muy buena reproducibilidad. Se aplicó un factor de discriminación isotópica de 1.06. La concentración de carbono inorgánico se calculó a través de la alcalinidad, la temperatura, la salinidad y el pH (Grasshoff et al., 1983).
Variables ambientales. La temperatura y salinidad se determinaron con una sonda automática (LF 191, WTW) el pH con el pH-96 (WTW) y la profundidad del disco Secchi con un disco blanco de 40 cm de diámetro. Para la determinación de la alcalinidad se agregaron 10 ml de HCl (0.01 N) a 40 ml de muestra, se midió el pH resultante y se calculó la alcalinidad según Grasshoff et al. (1983).
RESULTADOS Y DISCUSIÓN
Algunos datos metodológicos e hidrográficos se muestran en la tabla 1. Se reconoce bien el carácter salobre de los tres lugares de estudio. La Bahía de Kiel tuvo el valor más alto de salinidad con 20.4 %o en la superficie, seguido por el Fiordo de Kiel que fue un poco menor (17.8 %o), mientras que el Schlei tuvo solamente 7.3 ‰. En el Fiordo y la Bahía de Kiel se observó una estratificación marcada de la columna eufótica tanto en la temperatura como en la salinidad, la cual no se encontró en el Schlei, ya que en éste el espesor de la capa eufótica no sobrepasó 1 m.
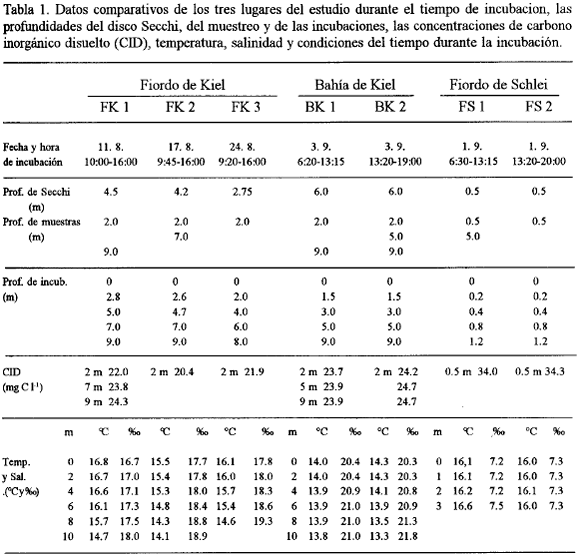
Hubo una diferencia marcada con respecto a la turbidez del agua indicada por las profundidades del disco Secchi, que fue de 6 m en la Bahía de Kiel y solamente 0.5 m en el Schlei. El alto grado de turbidez del Schlei se debe sobre todo a la presencia de concentraciones altas de plancton y detrito, mientras la presencia de partículas inorgánicas no tiene mucha importancia en su turbidez (Ehrhardt, 1970; Lenz, 1970).
La Fig. 2 muestra una diferencia marcada con respecto a la producción primaria entre las 3 áreas de estudio (determinado con el método de 14C). Sobre todo la PP por volumen muestra esta diferencia. La capa superficial del Schlei sobrepasó a la del Fiordo de Kiel por 9 veces y la de la Bahía de Kiel por más de 40 veces (valores promedios). La PP y la profundidad del disco Secchi tuvieron un comportamiento inverso, pero no existieron relaciones lineales (Tabla 1. y Fig. 2.). La situación cambia considerablemente cuando no contemplamos la PP por unidad de volumen sino por área Todavía predomina el Schlei, pero se ve que las diferencias ahora son relativamente pequeñas. En cuanto a la PP por volumen la relación entre la Bahía de Kiel, el Fiordo de Kiel y el Schlei fue de 1:4.8:43, la relación por área fue 1:2.6:3.1. En el Schlei, entonces, la producción primaria por m2 solamente fue 3 veces mayor que en la Bahía de Kiel. Esto dependió del hecho que el grosor de la capa eufótica en el Fiordo y la Bahía de Kiel fue mucho más grande que en el Schlei. Así la profundidad de compensación en el Schlei se encontró a 0.8 m mientras la del Fiordo se encontró a 5.2 m y la de la Bahía a 5.5 m (valores promedios).
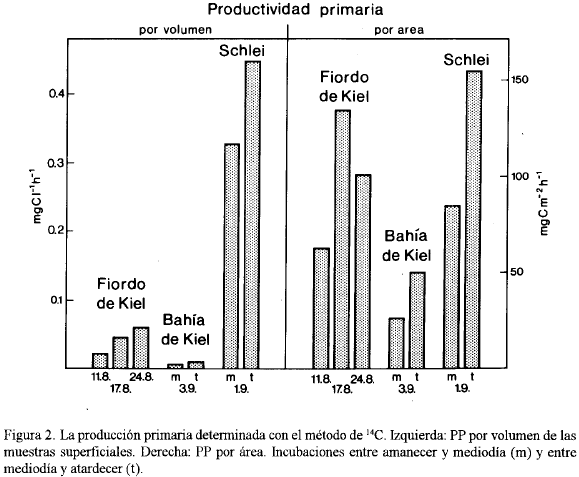
De la Fig. 3 se reconoce la distribución vertical de algunas de las determinaciones efectuadas comparativamente con el método de O2 y de 14C. En el Fiordo de Kiel se encontró una gran semejanza entre la producción primaria determinada con 14C (14C-PP) y la productividad primaria neta (PN) determinada con el método de O2, sobre todo en las 3 profundidades de incubación superiores. Más abajo las diferencias tienen que aumentar, ya que hasta la profundidad donde todavía existe un poco de luz la 14C-PP tiene que dar valores positivos aún cuando la PP fuera menor que la respiración. En la Bahía de Kiel (Fig. 3) la 14CPP se situó aprox. entre la PN y PB. Los valores que arrojó el método de 14C, entonces, fueron bastante mayores que la PN y menores que la PB. Además se encontró una diferencia grande entre la PN y la PB debido a la magnitud de la respiración la cual era pequeña en términos absolutos pero alta en relación a la producción primaria. Completamente diferente fue el resultado en el hipertrófico Fiordo de Schlei (Fig. 3), donde la 14C-PP fue más alta tanto que la PN como la PB, por lo menos en las tres profundidades superiores, o sea, en aquellas donde todavía hubo una producción neta.
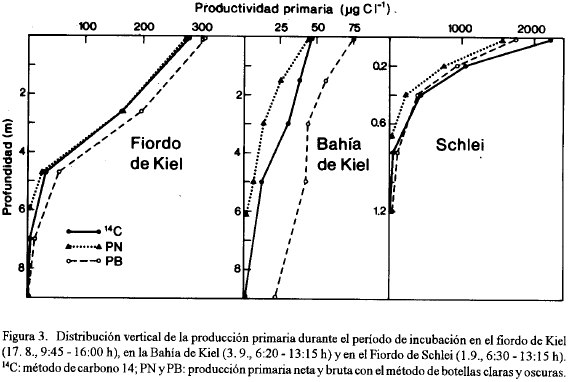
La Fig. 4 muestra la producción primaria por unidad de área. Debido a que nos interesa especialmente la relación entre la PN, la PB y la 14C-PP, se presentan no solamente los valores absolutos sino también lo los valores relativos tomando la 14C-PP como 100 %. En el Fiordo de Kiel se encontró una buena semejanza entre los resultados. Para el valor promedio de las 3 determinaciones en este lugar se obtuvo una PN que fue 9 % menor y una PB 36 % mayor que la 14C-PP lo que coincide bien con la teoría según la cual la 14C-PP tendría que ser más similar con la PN que con la PB después de una incubación prolongada.
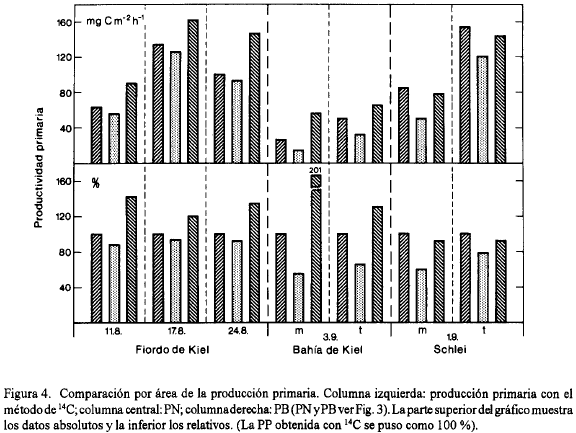
En la Bahía de Kiel la PN fue 40 % menor y la PB 72 % mayor que la 14C-PP. Las diferencias fueron marcadas, pero todavía la 14C-PP se encontró entre la PN y la PB. Esto no fue el caso para el Schlei donde la PN fue 32 % menor y la PB también 7 % menor que la 14C-PP. En esta área hipertrófica el método de 14C, entonces, dio el resultado más alto de la producción primaria lo que está difícil de entender como muestra la consideración siguiente:
En un cultivo de algas sin organismos heterotróficos el resultado que se obtiene con el método de 14C está influido por el tiempo de la incubación, ya que de la duración de la fotosíntesis depende la radioactividad específica de las sustancias orgánicas producidas. Con una incubación breve de pocas horas el método de 14C tendría que dar un resultado que debería asemejarse mucho a la PB, ya que las sustancias celulares respiradas (y junto con esto el CO2 liberado) apenas son radioactivas. En cambio, en el caso de una incubación prolongada (por lo menos 24 h según Hobson et al., 1976) los compuestos de la célula deberían haber llegado a un equilibrio isotópico, o sea, el CO2 liberado por la respiración debería haber tenido la misma actividad específica como el CO2 incorporado por la fotosíntesis. Esto significaría que la 14CPP tendría que ser casi igual a la PN. En el caso de una exudación significativa, la producción primaria obtenida con el método de 14C incluso debería ser menor que la PN ya que el método usual del 14C comprende solamente la PP particulada.
En los biótopos naturales las algas siempre están acompañadas por organismos heterótrofos, o sea, por el zooplancton y/o las bacterias. La producción primaria neta entonces no es la del fitoplancton sólo, sino la de la comunidad planctónica total; o sea que la PN del método de O2 toma en cuenta tanto la respiración de las algas como la de la parte heterótrofa del plancton. Los compuestos celulares respirados por los organismos heterótrofos (por lo menos los del zooplancton) no están en equilibrio isotópico ya que estos organismos requerirían una incubación de varios días para obtenerla, dado a la tasa relativamente reducida de su crecimiento. Esto significa que el método de 14C casi no es influido por la respiración del zooplancton. Así, incluso después de una incubación de varias horas cuando las algas ya han adquirido hace mucho el equilibrio isotópico, la PP obtenida con el método de 14C debería exceder a la PN obtenida con el método de O2 ya que la última incluye la respiración del zooplancton y de las bacterias.
Por otra parte una alta tasa de exudación bajaría la estimación de la 14C-PP. En la literatura existen valores muy divergentes de la exudación porcentual. Según Sharp (1977) la exudación constituye solamente un bajo porcentaje de la producción primaria, en cambio, otros autores opinan que podría constituir hasta la mitad de la producción total. Fogg (1983) da un resumen sobre los aspectos de la exudación. Según estudios propios (Gocke, en prep.) la exudación en la Bahía de Kiel se encontró entre 1.7 y 31 %, en el Schlei se encontraron solamente valores entre 0.3 y 3.0 %. Es obvio que por lo menos en el Schlei la baja exudación casi no disminuye el valor de la 14C-PP y por lo tanto puede ser una de las razones por las cuales el método de 14C dio valores relativamente altos en este lugar.
Sin embargo, según las consideraciones teóricas mencionadas arriba, la 14C-PP no debería ser más alta que la PB, como se observó en el Schlei (Fig. 4). Una de las razones podría ser, que aquí el cociente fotosintético, con el cual se convirtieron las unidades de O2 en unidades de C, no tenia el valor de 1.2 como en las otras 2 áreas. Normalmente las algas prefieren NH4+ como fuente de nitrógeno lo que resultaría en un CF de 1.2 (Eppley et al., 1969; Axler et al., 1982; Berman et al., 1984). Según Bell y Kuparinen (1984) el CF subiría a 1.57, si las algas utilizaran N03" como única fuente de compuestos nitrogenados, lo que equivaldría a 0.239 mg de C en vez de 0.313 mg por 1 mg de O2 como en el caso de un CF de 1.2. Sin embargo, en el caso del Schlei el CF no debería ser mayor de 1.2 sino más bien menor, ya que un CF por encima de 1.2 disminuiría la PN y PB. Si las algas producen solamente carbohidratos, el CF valdría 1. Esta situación podría ser posible en el caso de fitoplancton senescente. A finales del verano es muy posible que una situación semejante pudo haber ocurrido en el Schlei. Si solamente una cierta parte del fitoplancton tuviera un CF cercano a 1, el cociente fotosintético integral para todo el fitoplancton caería un poco debajo de 1.2 lo que bastaría para explicar la discrepancia observada entre el resultado observado con los 2 métodos.
CONCLUSIONES
Usando los métodos de oxígeno y de 14C para la determinación de la producción primaria se encontró en los dos lugares menos eutroficados una progresión: producción primaria neta < producción primaria determinada con 14C < producción primaria bruta empleando un cociente fotosintético a 1.2. Aplicando el mismo cociente fotosintético la 14CPP sobrepasó ligeramente la producción primaria bruta determinada con el método de oxígeno en el lugar hipertrófico. Esta desviación de la teoría se podría explicar con un fitoplancton parcialmente senescente resultando en un cociente fotosintético un poco menor de 1.2. Como resultado del estudio comparativo, se puede decir que los dos métodos dan valores comparables en biótopos acuáticos de diferentes grados de eutroficación.
AGRADECIMIENTOS
Agradecemos mucho la ayuda financiera del Servicio Alemán de Intercambio Académico (DAAD) que hizo posible la estadía del segundo autor en Alemania. Expresamos nuestros agradecimientos al capitán y a la tripulación del barco científico "Littorina" por su valioso apoyo durante los cruceros. Además nos es grato expresar nuestros sinceros agradecimientos a tres evaluadores incógnitos que contribuyeron mucho a la elaboración del artículo por sus valiosas críticas y sugerencias.
BIBLIOGRAFÍA
1 Axler, R.P.; R.M. Gersberg y C.R. Goldmann. 1982. Inorganic nitrogen assimilation in asubalpine lake. Limnol. Oceanogr., 27: 53-65. [ Links ]
2 Bell, R.T. y J. Kuparinen. 1984. Assessing phytoplankton and bacterioplankton production during early spring in Lake Erken, Sweden. Appl. Environ. Microbiol., 48: 1221-1230. [ Links ]
3 Berman, T.; B.F. Sherr, E. Sherr; D. Wynne y J.J. McCarthy. 1984. The characteristics of ammonium and nitrate uptake by phytoplankton in lake Kinneret. Limnol. Oceanogr., 29: 287-297. [ Links ]
4 Bodungen, B. v. 1975. Der Jahresgang der Nahrsalze und der Primarproduktion des Planktons in der Kieler Bucht unter der Berücksichtigung der Hydrographie. Tesis Doct. Univ. Kiel, Alemania, 116 p. [ Links ]
5 Bryan, J-R.; J.P. Riley y P.J.LeB. Williams. 1976. A Winkler procedure for making precise measurements of oxygen concentration for productivity and related studies. J. Exp. Mar. Biol. Ecol., 21: 191-197. [ Links ]
6 Davies, J.M. y P.J.LeB. Williams. 1984 Verification of 14C and O2 derived primary organic production measurements using an enclosed ecosystem. J. Plankton Res., 6: 457-474. [ Links ]
7 Ehrhardt, M. 1970. Partikularer organischer Kohlenstoff und Stickstoff sowie gelöster organischer Kohlenstoff. En Chemische, mikrobiologische und planktologische Untersuchungen in der Schleiim Hinblick auf deren Abwasserbelastung. Kieler Meeresforsch., 26: 138-144. [ Links ]
8 Eppley, R.W.; J.L. Coatworth y L. Solorzano. 1969. Studies on nitrate reductase in marine phytoplankton. Limnol. Oceanogr., 14: 194-205. [ Links ]
9 Fogg, G.E. 1983. The ecological significance of extracellular products of phytoplankton photosynthesis. Bot. Mar., 26: 3-14. [ Links ]
10 Gaarder, T. y H.H. Gran. 1927. Investigations of the production of plankton in the Oslo Fjord. Rapp. et Proc. Verb., Cons. Internat. Explor. Mer., 42: 1-48. [ Links ]
11 Gocke, K. y G. Rheinheimer. 1991. Influence of eutrofication on bacteria in two fjords of the Western Baltic. Int. Revue ges. Hydrobiol., 76: 371-385. [ Links ]
12 Grande, D.K; P.J.LeB Williams; J. Marra; D.A. Purdie; K. Heinemann; R. W. Eppley y M.L. Bender. 1989. Primary production in the North Pacific Gyre: a comparison of rates determined by the 14C, O2 concentration and 180 methods. Deep-Sea Res. 36: 1621 1634. [ Links ]
13 Grasshoff, K.; M. Ehrhardt y K. Kremling (Eds.). 1983. Methods of seawater analysis, Verlag Chemie, Weinheim, Alemania, 419 p. [ Links ]
14 Green, E.J. y D.E. Carrit. 1966. An improved iodine determination flask for whole bottle titration. Analyst., 91: 207-208. [ Links ]
15 Hobson, L.A.; W.J. Morris y K.T. Picquet. 1976. Theoretical and experimental analysis of the 14C technique and its use in studies of primary production. J. Fish. Res. Bd. Canada, 33: 1715-1721. [ Links ]
16 Irwin, B. 1991. Coulometric measurement of primary production, with comparison against dissolved oxygen and 14C methods in a seasonal study. Mar. Ecol. Prog. Ser. 71: 97-102. [ Links ]
17 Kandier, R. 1959. Hydrographische Beobachtungen in der Kieler Förde 1952-1957. Kieler Meeresforsch., 15: 145-156. [ Links ]
18 Lenz, J. 1970. Seston-, Chlorophyll- und Eiweißgehalt. En Chemische, mikrobiologische und planktologische Untersuchungen in der Schlei im Hinblick auf deren Abwasserbelastung. Kieler Meeresforsch., 26: 180-192. [ Links ]
19 Lenz, J. 1977. Hydrographie conditions. En G. Rheinheimer (Ed.): Microbial ecology of a brackish water environment. Springer Verlag Berlin, Heidelberg, New York: 12-25. [ Links ]
20 Margalef, R. 1986. Ecología. Edit. Omega, Barcelona, España, 951 p. [ Links ]
21 Moigis, A.G. 1983. Zur Größenstruktur und Ökologie des Phytoplanktons in der Kieler Förde unter dem besonderen Aspekt einer möglichen Ölverschmutzung. Tésis Doct., Univ. Kiel, Alemania, 171 p. [ Links ]
22 Nellen, W. y G. Rheinheimer. 1970. Salzgehalt. En Chemische, mikrobiologische und planktologische Untersuchungen in der Schlei im Hinblick auf deren Abwasserbelastung. Kieler Meeresforsch., 26: 123-126. [ Links ]
23 Parsons, T.R.; M. Takahashi y B. Hargrave. 1984. Biological océanographie processes. 3rd. edit. Pergamon Press, Londres, 330 p. [ Links ]
24 Peterson, B.J. 1980. Aquatic primary productivity and the 14C-CO2 method: a history of the productivity problem. Annual Rev. Ecol. Systems, 11: 359-385. [ Links ]
25 Raine, R.C.T. 1983. The effect of nitrogen supply on the photosynthetic quotient of natural phytoplankton assemblages. Bot. Mar., 26: 417-423. [ Links ]
26 Raymont, J.E.G. 1980. Plankton and productivity in the oceans. 2nd edit. Pergamon Press, Londres, 489 p. [ Links ]
27 Schiemann, S., 1974. Die Primärproduktion des Phytoplanktons der Schlei und des Windebyer Noors im Jahre 1972. (Ein Vergleich von Methoden und Biotopen.) Tésis Doct., Univ. Kiel, Alemania, 173 p. [ Links ]
28 Sharp, J.H. 1977. Excretion of organic matter by marine phytoplankton - Do healthy cells do it?. Limnol. Oceanogr., 22: 381 -399. [ Links ]
29 Steemann Nielsen, E., 1952. The use of radioactive carbon (C14) for measuring organic production in the sea. J. Cons. Perm. Int. Explor. Mer, 18: 117-140. [ Links ]
30 Strickland, J.D.H. 1960. Measuring the production of marine phytoplankton. Bull. Fish. Res. Bd. Canada, 122: 1-172. [ Links ]
31 Vollenweider, R.A. 1974. Methods for measuring production rates. En R.A. Vollenweider (Ed.): Primary production in aquatic environments. IBP Handbook No 12, Blackwell Scientific Publications, Oxford, Londres, 225 p. [ Links ]
32 Williams, P.J.LeB., R.C.T. Raine y J.R. Bryan. 1979. Agreement between the 14C and oxygen methods of measuring phytoplankton production: reassessment of the photosynthetic quotient. Oceanol. Acta, 2: 411-416. [ Links ]
33 Williams, P.J.LeB. y N.W. Jenkinson. 1982. A transportable microprocessor-controlled precise Winkler titration suitable for field station and shipboard use. Limnol. Oceanogr., 27: 576-584. [ Links ]
34 Williams, P.J. LeB.; K.R. Heinemann; J. Marra y D.A. Purdie. 1983. Comparison of 14C and O2 measurement of phytoplankton production in oligotrophic waters. Nature, 305: 49-51. [ Links ]
