










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.23 no.1 Santa Marta Jan./Dec. 1994
ASPECTOS BIOLÓGICOS DEL CALAMAR ARGENTINO ILLEX ARGENTINUS (CASTELLANOS) CAPTURADO POR LA FLOTA CUBANA DE PESCA EN EL TALUD PATAGÓNICO DURANTE 1987
Rafael Tizol, Claudio García y Julio López
Centro de Investigaciones Pesqueras, Barlovento, Santa Fe, Playa, La Habana, CUBA.
RESUMEN
En 1987, se inicia un programa de muestreos biológicos del calamar argentino illex argentinus a bordo de barcos comerciales de la flota cubana de pesca que operan en la plataforma patagónica. La talla de los organismos para ambos sexos presenta un comportamiento estratificado por profundidades, los ejemplares de mayor tamaño se encuentran a profundidades superiores y viceversa, lo cual se ajusta a una recta de regresión (r2 = 0.88; α = 0.05). Las hembras alcanzaron largos promedio del manto superior es a los machos, incrementándose la presencia de machos en las capturas desde enero a abril. Se estimaron los largos promedio de primera maduración, como 20.5 y 26.5 cm de largo del manto para machos y hembras respectivamente. Se observó un aumento de la proporción de individuos maduros de febrero a abril, haciéndose más marcado este fenómeno en los machos, que en su mayoría presentaban abundantes espermatóforos en el saco. Se obtuvo la relación largo-peso para hembras y machos y para la combinación de ambos sexos.
ABSTRACT
In 1987 a program for the biological sampling of squid illex argentinus, was begun an board of a fishing ship belonging to the Cuban Fisheries Fleet, wich operates in the Patagonian Shelf. The lengths of both sexes presented a estratified pattern with individuals of larger size found in higher depths and viceversa; this pattern is adjusted to a regression line (r2 = 0.88; α = 0.05). Females reached average length larger than males and the rate of males in catches increased from January to april. The first maturation average length was estimated about 20.5 cm for males and 26.5 cm for females. An increase of the rate of mature individuals was observed between february and april emphazasing in males which showed abundant espermatophores in the sac.
INTRODUCCIÓN
El calamar argentino Illex argentinus (Castellanos, 1960), constituye uno de los objetivos pesqueros más importantes de la plataforma patagónica. A partir de 1978 se comienzan a obtener volúmenes significativos de esta especie en el área, con 226000 ton en 1985 (Sato y Hatanaka, 1983), con Polonia y Japón como principales productores (FAO, 1987). Cuba se incorpora a esta pesquería ese mismo año realizando sus faenas en las zonas fuera de las 200 millas de Zona Económica Exclusiva.
Las principales características biológicas del calamar argentino han sido descritas por diferentes autores. Brunetti (1982) Y Hatanaka, et al (1985) han investigado su ciclo de vida, distribución, etc. En Cuba hasta el momento no se había comenzado trabajo alguno de investigación sobre esta especie, debido en primer lugar, al carácter prospectivo de nuestras pesquerías. En 1987, considerando la necesidad de apoyar la actividad de la Flota Cubana de Pesca en esta área con una base científica, se comenzó el primer programa de investigación del Illex argentinus. Los resultados preliminares del mismo se exponen en el siguiente trabajo.
MATERIALES Y MÉTODOS
Entre el 1ro. de enero y el 12 de abril de 1987, se llevaron a cabo muestreos biológicos del calamar argentino, a bordo de barcos comerciales cubanos del tipo TACSA 95, arrastreros por la popa de 106 m de eslora. Las pesquerías se realizaron al oeste de la República Argentina, en la zona del talud fuera de los límites de las 200 millas de Zona Económica Exclusiva, entre los 44° y los 47° de latitud sur y los 61° y 59° de longitud Este, en profundidades entre 100 y 160 m.
Durante el período de trabajo se muestrearon 200 ejemplares en cada lance efectuado, considerando el largo dorsal del manto, el sexo y el estadio de madurez sexual. El largo dorsal del manto (desde la protuberancia anterodorsal del manto hasta el apex de la aleta caudal) se tomó al 0.5 cm más cercano, mientras que la información sobre los aspectos restantes se recopiló mediante la observación interna de los especímenes. En total se muestrearon 13568 ejemplares: de estos 6555 hembras y 7013 machos. Además, se obtuvo una muestra estratificada por clase de largo y sexo de 1076 ejemplares para el cálculo de la relación largo-peso, de estos 446 machos (largo del manto entre 14.5 y 28.7) y 630 hembras (largo del manto entre 14.5 y 35.5 cm). Para determinar la madurez sexual de las hembras se utilizó la escala de Burukovski (1984) y la de Nigmatullin et al (1984) para evaluar la madurez sexual de los machos. Los datos obtenidos en los muestreos fueron agrupados en períodos de 10 días. Se estudió la variación de la composición por largo durante toda la etapa de trabajo, con referencia a los estadios de madurez sexual y a las profundidades, relacionando este último con el largo mediante un análisis de regresión. Se compara, además, el largo medio del manto de especímenes capturados en diferentes latitudes.
Se calculó la relación largo-peso para machos y hembras combinados y para cada sexo por separado. Las relaciones por sexo se compararon mediante un análisis de covarianza para encontrar diferencias estadísticamente significativas entre las mismas (a=0.05).
La proporción porcentual de hembras y machos así como la de los diferentes estadios gonadales en el período de muestreo es también analizada. Para la obtención del largo de primera maduración se consideraron los especímenes en el estadio V, teniendo en cuenta dos variantes del mismo parámetro, el largo mínimo de primera maduración y el largo del manto a partir del cual el 50 % de los ejemplares están maduros.
RESULTADOS Y DISCUSIÓN
Las hembras alcanzaron largos del manto máximos y medios mayores que los machos durante todo el período de muestreo. En la tabla 1 se ofrecen los datos de tallas mínimas, máximas y medias, así como la dispersión por décadas. La talla media va en aumento a partir de la primera década de febrero, mientras que en el mes de enero permanece con poca variación.
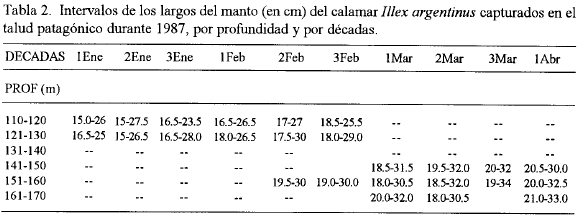
La profundidad es un factor importante en la distribución del calamar (Ngai y Uosumi, 1980; Hatanaka, et al, 1985). Los ejemplares de mayor talla fueron encontrados a profundidades superiores y viceversa (tabla 2), el análisis de regresión ofrece un buen ajuste a una línea recta (r2=0.88; α= 0.05). Si asumimos que el movimiento de la flota pesquera coincide con las áreas de mayores concentraciones, en el mes de enero y la primera década de febrero los calamares se distribuyeron en profundidades entre 100 y 130 m. Por otra parte, en la segunda década de ese mes se observa la aparición de ejemplares de mayor talla a profundidades superiores y con un desarrollo gonadal avanzado.
Podemos considerar entonces que las concentraciones de calamares en el mes de enero y la primera década de febrero, con una talla menor (largo medio del manto entre 19.8 y 20.5 cm) e inferior desarrollo gonadal no corresponden a agrupaciones de reproducción, mientras que el segundo grupo con un largo medio entre 22.4 y 35.1 cm, que aparece posteriormente, con un desarrollo gonadal superior, corresponde a concentraciones predesove. Brunetti (1982) también hace alusión a la presencia en este periodo de cardúmenes predesove en el talud de la plataforma argentina. Las migraciones de desove son más densas e involucran a la mayor parte de la población, por tanto al suceder éstas es que ocurren los mayores rendimientos en la pesquería. La aparición de estos cardúmenes ocurre de una forma rápida, lo que en ocasiones no permite tener dispuestas las condiciones necesarias para aplicar un esfuerzo máximo de pesca Este aspecto es de importancia y debe ser considerado, ya que la presencia de estos indicadores biológicos (como es la aparición de ejemplares de mayor talla y avanzado estado de madurez sexual), nos da la posibilidad de conocer con anticipación el inicio de la etapa de altos rendimientos y acondicionar la flota para obtener los mejores rendimientos ya que posteriormente, como se conoce, éstos descienden tan bruscamente como aumentaron.
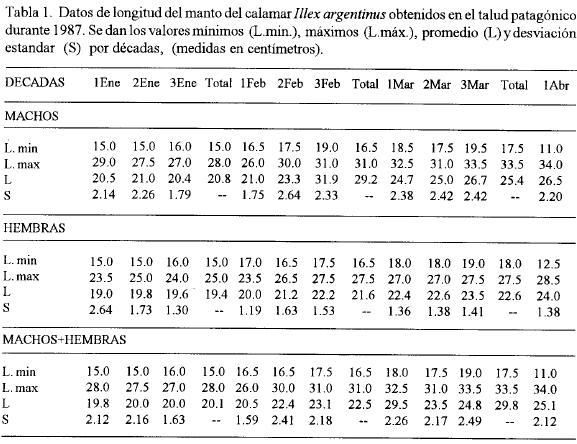
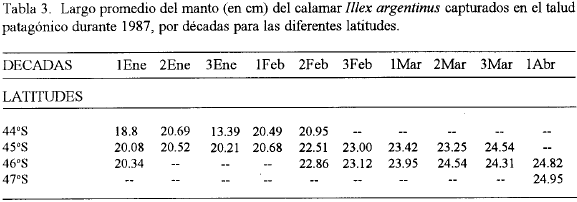
Los largos medios del manto distribuidos por latitudes (tabla 3), muestran que los ejemplares capturados más al Sur presentan mayores tallas que los encontrados en latitudes inferiores. Brunetti (1982), menciona esta característica, atribuyéndola al desfase del desove en el tiempo, comenzando por el extremo sur del área de distribución y no simultáneamente en toda la zona y asociándolo con la variación de la temperatura. La proporción de hembras y machos fue favorable a las primeras durante los meses de enero y febrero. La presencia de machos en las capturas fue superior entre un 5 y un 12% a partir de la primera década de marzo. En general, en ningún caso un sexo predominó sobre el otro en más del 18% y la mayor parte del tiempo estas diferencias se mantuvieron entre un 2 y un 7 %.
Los estadios gonadales se comportaron para los machos con un predominio en la primera década de febrero de los estadios I, II y III (23.6, 31.6 y 31.1% respectivamente). Los ejemplares que presentan los estadios IV y V de madurez sexual tienen largos del manto superiores a los 21 cm y su proporción en ésta época varió entre 3.5 y 10.2%. A partir de la segunda década de febrero se produce un aumento brusco del número de machos en el estadio V (62.75%), mientras que los estadios I, II y III alcanzan proporciones inferiores a los obtenidos en la década anterior (1.7, 16.6 y 9.8% respectivamente). Este crecimiento del número de especímenes en estadio V se mantiene hasta alcanzar un máximo de 97.4% en la tercera década de marzo, disminuyendo en la primera década de abril a un 75.6%, distribuido el resto entre los estadios II, III y IV.
En el caso de las hembras ocurre un proceso similar, aunque manteniendo la presencia de diferentes estadios de maduración y sin llegar a valores extremos como es el caso de los machos. En la primera década de febrero predomina el estadio III (49.3%) seguido del II y del IV (23.6 y 21.2%), mientras que el estadio V tiene solo un 2%. En la segunda década sin embargo, la producción de ejemplares en este estadio alcanza un 33.5%, la cual se mantiene durante todo el período posterior alcanzando un máximo de 55% en la tercera década de marzo, distribuido el resto entre los estadios II, III y IV.
Estas variaciones de los estadios gonadales en los muestreos están muy relacionadas con los movimientos de la flota pesquera que realiza sus faenas buscando las áreas de mayores concentraciones de la especie. De esa forma en el mes de enero y la primera década de febrero, las pesquerías se desarrollaron a menores profundidades sobre cardúmenes que no son de reproducción. Posteriormente en la segunda década de febrero a la pesquería se incorporan paulatinamente los cardúmenes de reproducción, por lo que es posible encontrar en las capturas ejemplares de mayor talla y con un nivel superior de desarrollo gonadal. Finalmente el arribo masivo de las agrupaciones predesove provocan un salto brusco en la proporción de calamares en estadios avanzados de desarrollo gonadal y un aumento de los rendimientos. Ya en la primera década de abril ocurre una disminución de la proporción de ejemplares en estadio V junto con los rendimientos en las capturas, aparentemente como resultado del movimiento de los primeros grupos de desovantes hacia mayores profundidades para su reproducción. En nuestro caso los muestreos no fueron continuados, pero se conoce que la flota pesquera se traslada a profundidades superiores a los 400 m, donde continúa la pesquería sobre estos cardúmenes antes de su movimiento hacia el océano abierto donde se produce el desove (Brunetti, 1982; FAO, 1983; Hatanaka et al, 1985).
Se ha señalado (FAO, 1983) la marcada influencia entre las proporciones de los estadios de madurez sexual, para machos y hembras, especialmente en el estadio V, en el cual se puede capturar una elevada proporción de machos en los momentos cercanos al desove, mientras que en las hembras aunque el porcentaje en este estadio puede ser relativamente alto, no alcanza esta proporción. Según nuestros resultados, los machos poseen un largo de primera maduración de 17.6 cm, mientras que las hembras tienen un largo de primera maduración de 21.6 cm. Por otra parte los valores de Lm50 se presentan marcadamente superiores para las hembras con 26.4 cm, en comparación con un 20.2 cm para los machos (tabla 4).

Las ecuaciones de relación largo-peso se ofrecen en la tabla 4. El análisis de covarianza no arrojó diferencias estadísticamente significativas (Fteórica = 3.00; Fcalculada= 0.172) entre las ecuaciones para hembras y machos, aunque se ha observado que los machos, especialmente los que poseen un largo del manto superior a los 21 cm tienen un peso mayor.
BIBLIOGRAFÍA
1 Brunetti, N. 1982. Estado actual sobre el conocimiento del calamar (Mimeografiado). [ Links ]
2 Burukovski, R.N., V.N. Freman y C.N. Nigmatullin. 1984. Reproductive biology and scale of maturity stage of reproductive system of female squid (illex ilticebrosus) NAFO SCR Doc 84/IX/120. [ Links ]
3 Hatanaka, H., S. Kawahara, Y. Uozumi y S. Kasehara. 1985. Comparison of the cycles of five Ommastrephid squid fishes by Japan: Thodarodes pacificus; Hlex illicebrosus; Nototodarus sloani sloani; and Nototodarus sloani gouedi. NAFO Sci. Coun. Studies, 9. [ Links ]
4 FAO. 1983. Informe del grupo ad hoc de trabajo sobre los recursos pesqueros de la plataforma continental patagónica. FAO Inf. Pesca, 297. [ Links ]
5 FAO. 1987. Anuario estadístico de pesca. Capturas y desembarques. Colección FAO Estadística, 60. [ Links ]
6 Ngai, T. y Y. Uozumi. 1980. Estimation of the abundance index for Hlex based on Japanese fishery operations along the edge of the Scotian Shelf in 1979. NAFO SCR Doc 80/XX/9. [ Links ]
7 Nigmatullin, Ch.N., R.N. Savirov y Y.M. Boeman. 1984. Reproductive biology and scale of maturity stages of the reproductive system of male squid, illex illicebrosus. NAFO SCR Doc IX/119. [ Links ]
8 Sato, Y. y H. Hatanaka. 1983. Progresos realizados en la evaluación mundial de cefalópodos. FAO Doc. Tec. de Pesca 231. [ Links ]
