










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.23 no.1 Santa Marta Jan./Dec. 1994
PRESENCIA DE LOS OCTOCORALES STYLATULA DIADEMA BAYER (PENNATULACEA) Y CARIJOA RIISEI (DUCHASSAING Y MICHELOTTT) (TELESTACEA) EN LA COSTA CARIBE COLOMBIANA
Juan Armando Sánchez M.
Instituto de Investigaciones Marinas de Punta de Betín-INVEMAR, A.A, 1016, Santa Marta, Colombia.
RESUMEN
Con el fin de colectar octocorales telestáceos y penatuláceos se revisaron diferentes ecosistemas entre 0 y 30 m de profundidad, mediante buceo SCUBA y simple a pulmón que comprendían litorales rocosos, fondos arenosos y arrecifes coralinos, en un área que incluye a Cartagena, las Islas del Rosario, Tierra Bomba, Barú y San Bernardo, los bancos de Salmedina y Bushnell, las ensenadas de Zapzurro y Capurganá (8°20'-10°45' N; 75°50'-77°25' W), en la región de Santa Marta (11°14'50" N; 74°15' W) y la Guajira (11°56'58" N; 72°16'18" W), en el Caribe colombiano. Se encontró por primera vez en el mar Caribe y la costa colombiana a Stylatula diadema Bayer (Virgulariidae: Pennatulacea) que se halló solamente en fondos arenosos blandos entre 25 y 30 m a la salida de la Bahía de Cartagena y en el bajo Mangles en las islas de San Bernardo. Se amplió notablemente la distribución geográfica para el mar Caribe y Colombia de Carijoa riisei (Duchassaing y Michelotti) (Telestidae: Telestacea), con un gran rango batimétrico (0.5-30 m) encontrado sobre todos los habitats revisados. La poca presencia de telestáceos y penatuláceos en aguas someras de esta región, sugiere que al igual que en todo el mar Caribe, los octocorales están mejor representados por los gorgonáceos que por los demás órdenes, mostrando un comportamiento inverso con la fauna del Indopacífico, en donde dominan los alcyonáceos sobre los demás órdenes.
ABSTRACT
Through SCUBA and skin diving various shallow water ecosystems (rocky shores, soft bottoms and coral reefs, 0-30 m deep) were surveyed to collect telestaceans and penatulaceans octocorals; the areas considered were Cartagena, the Rosario islands, Tierra Bomba island, Barú island, San Bernardo islands, Bushnell and Salmedina banks, Capurgana and Zapzurro harbours (8°20'-10°45' N; 75°50'-77°25' W), the Santa Marta area (11°14'50" N; 74°15' W) y and the Guajira (11°56'58" N; 72°16'18" W), Colombian Caribbean. Stylatula diadema Bayer (Virgulariidae: Pennatulacea) is first recorded for the Caribbean sea and has been found inhabiting soft sand bottoms between 25 and 30 m, at the outer end of Cartagena Bay and the Mangles Bank in San Bernardo islands. The geographic distribution of Carijoa riisei (Duchassaing y Michelotti) (Telestidae: Telestacea) was widen from the Caribbean and Colombia, it has been found in a broad bathymetric range (0.5-30 m) in all the habitats surveyed. Few records of telestaceans and pennatulaceans in this región, as in the rest of the Caribbean, suggest that the octocorallian fauna is mostly represented by gorgonians than by other orders, and shows an inverse relationship with the Indopacific fauna, where the alcyonaceans are the dominant order.
INTRODUCCIÓN
Los octocorales son antozoarios sésiles generalmente coloniales caracterizados por tener sus pólipos con ocho tentáculos pinados, presentar un esqueleto formado por espículas calcáreas en todos los órdenes y estructuras axiales rígidas de sustancias córneas, fibrocristalinas o de espículas fusionadas dependiendo del orden. Además, poseen una compleja red de interconexión gastrodérmica o solenia, colocándose como uno de los grupos más evolucionados entre los celenterados sésiles. Se distribuyen por todos los mares, encontrándose desde aguas someras hasta fondos abisales (Bayer 1956, 1961).
El mar Caribe se ha caracterizado por la gran diversidad y abundancia de octocorales en especial de gorgonáceos, siendo la región geográfica con más número de especies de Plexauridae y Gorgoniidae (Kinzie, 1973). El orden Alcyonacea es mucho más diversificado hacia el Indopacífico y los demás órdenes tienen una distribución más global, pero siempre con una tendencia pantropical (Sammarco y Coll, 1992). Los penatuláceos aunque son un grupo relativamente reducido tienen una amplia distribución geográfica en especial hacia el Atlántico e Indopacífico. En el Atlántico occidental estos se encuentran representados con dos subórdenes, tres familias y cinco géneros: Acanthoptilum, Renilla, Stylatula, Virgularia y Sclerobelemnon (Bayer, 1957, 1959, 1961). Caso contrario ocurre con los telestáceos que aunque son mas conocidos en estas aguas, solo se encuentra la familia Telestidae, representada por dos géneros en aguas someras: Telesto y Carijoa. Anteriormente se consideraba que todas las especies del Caribe se incluían dentro del género Telesto, pero Carijoa riisei, Coperculata y Crupicula presentaban espículas y estructuras diferentes con respecto a las demás especies de Telesto, suficientes como para ubicarlas dentro de un género aparte (Bayer, 1981), posición que se había retardado en esclarecer, a causa de una descripción muy somera de las especies tipo de Telesto y Carijoa (Bayer, 1961).
En el mar Caribe se desconocen informes sobre octocorales penatuláceos. De los telestáceos en el Caribe colombiano se conoce la presencia de C. operculata en el Morro Grande, Bahía de Santa Marta (Botero, 1987) y una observación sin localidad exacta de C. riisei para el archipiélago del Rosario (Manjarres, 1979). A causa del relativo desconocimiento de estos dos órdenes en esta área, así como la gran cantidad de arrecifes coralinos y ecosistemas relacionados aptos para el desarrollo de octocorales en la costa Caribe colombiana, se revisó la zona para colectar e identificar las especies de aguas someras.
METODOLOGÍA
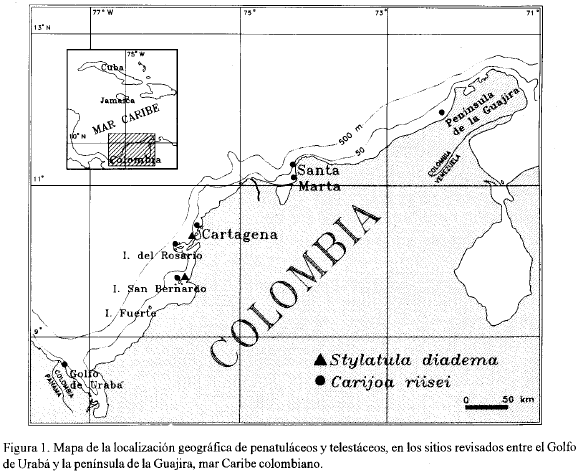
Durante 1990-1992 se exploró periódicamente mediante buceo SCUBA y a pulmón en arrecifes coralinos, litorales rocosos y fondos arenosos dentro de un área que comprende las costas de Cartagena, la islas de Tierra Bomba, Barú, del Rosario y el banco de Salmedina (10°45'-10°20' N; 75°30'-75°50' W), las islas de San Bernardo (9°40'-9°50' N; 75°45'-75°55' W), el banco Bushnell (9°25'08" N; 76°2'00" W), las ensenadas de Zapzurro y Capurganá en el golfo de Urabá (8°20'-8°35' N; 77°00,-77°25' W). Adicionalmente durante 1994 se realizaron algunas revisiones en el área de la bahía de Santa Marta (Punta de Betín y el Morro: 11°14'50" N; 74°15' W), la plataforma continental entre Ciénaga y Santa Marta (pesca de arrastre en el B/I ANCON) y la Guajira (11°56'58" N; 72°16'18" W), todas las localidades sobre el Mar Caribe suroccidental en Colombia (figura 1).
Los octocorales penatuláceos fueron fijados en seco para su identificación, y los telestáceos en seco y húmedo mediante etanol al 70 %. Adicionalmente se anotó la profundidad, coloración y hábitat de las especies colectadas. Se utilizaron para su identificación las claves de Bayer (1961). Las medidas de las colonias fueron realizadas con un calibrador de precisión hasta de 0.01 mm. Las espículas de los octocorales se dibujaron mediante calco sobre proyección de placas y se midieron por medio de una placa-escala Leitz™ de un milímetro con precisión de 0.001 mm, también calcada y usada para medir sobre los dibujos. Para cada movimiento del microscopio de proyección se calcaba una escala diferente y además se trató de que la proyección fuese a 40 cm de distancia y que la circunferencia proyectada tuviese máximo 20 cm de diámetro, o el ancho de una hoja tamaño carta. Las muestras identificadas fueron confirmadas por el Dr. F.M. Bayer del Instituto Smithsonian de Washington D.C. y depositadas en el Museo Nacional de Historia Natural (USNM) de la misma ciudad. Una colección de trozos de las mismas muestras enviadas a Washington, se ubicó en la colección del Instituto de Investigaciones Marinas de Punta de Betín (INVEMAR-OCTO), Santa Marta, Colombia.
Subclase OCTOCORALLIA Haeckel,1866 Orden PENNATULACEA Verrill, 1865
Suborden SUBSELLIFLORAE Kükenthal, 1915
Familia VIRGULARHDAE Verrill, 1868
Genero Stylatula Verrill, 1864
Stylatula diadema Bayer, 1959
Stylatula diadema Bayer 1959, p.38 (Surinam) - Bayer 1961, p.308 (Surinam).
Diagnóstico: Stylatula delgado con 8-12 pares de pólipos frondosos hasta de 2 cm, con 25-30 autozooides y dispuestos en dos hileras alternadas en zigzag dorsal y ventralmente. La base polipar esta compuesta por 8-12 largas y gruesas agujas entre 2.2-2.7 mm de largas y numerosas agujas pequeñas. No presenta espículas en las paredes corporales ni en los tentáculos. Las espículas sobre el raquis se disponen en dos angostas bandas laterales que parten desde la base espicular del pólipo. Los sifonozooides se disponen bajo la superficie de la fronda polipar, dentro de uno o dos círculos cerca a la región distal del vientre de la fronda. El axis es muy rígido, de sección redondeada a cuadrangular y tiene un leve canalete a cada lado. (Bayer, 1959; 1961).
Descripción: El material examinado (USNM 92965, INV. OCTO-289, 92966 INV. OCTO-290) presenta características similares, con un corallum entre 30-50 cm de largo, una coloración del raquis blanca y el tejido vivo incoloro levemente blanco, de sección redondeada cuadrangular con dos leves surcos ventral y dorsalmente bien marcados, en especial hacia la región basal, con 1-3 mm de grosor cerca al pedúnculo basal y generalmente de 1 mm hacia la región apical. Las colonias se colectaron perfectamente erguidas y enterradas unos 2.5-4 cm en la arena. Las frondas polipares se disponían hacia la región ventral del raquis pero de manera bilateralizada entre 1-2 cm a cada lado como una pluma. El material en seco presentaba claramente los abanicos de 8-12 espículas que soportaban las frondas polipares compuestos de agujas gruesas y lisas generalmente mayores a 1 mm de largo (figura 2j), las espículas pequeñas algunas de forma similar y otras en forma de agujas delgadas, teniendo entre 0.1-0.5 mm de longitud (figura 2k).

Comentario: Aunque las muestras de Stylatula diadema fijadas en seco no son el mejor material para una identificación hasta especie y pueden llegar a confundirse con especies de Acanthoptilum, los abanicos espiculares se conservan intactos en las muestras recientes, especialmente en la región basal, en donde la forma de las agujas y su tamaño son muy similares a las de S. diadema en condiciones de fijación en húmedo. Además, esta especie es el único representante de los virguláridos en la costa Norte de Sudamérica (Bayer, 1961).
Habitat: Fondos arenosos blandos y llanos en aguas claras entre 25-30 m de profundidad en donde es muy abundante.
Distribución: Océano Atlántico: Surinam; Mar Caribe (primer registro): Colombia, canal navegable de Bocachica entre la Bahía de Cartagena y la isla de Tierra Bomba, y el bajo Mangles en las islas de San Bernardo.
Material examinado: USNM No.92966, INVEMAR OCTO-289: Dos colonias del Bajo Mangles, al este de isla Mangles en el Archipiélago de San Bernardo, prof. 25 m, 26 de julio de 1991 (color blanco con pólipos entre blanco y transparente, muy abundante en fondo arenoso). USNM No.92967, INV. OCTO-290: Cuatro 4 colonias del canal de Bocachica a la salida de la Bahía de Cartagena en la isla de Tierra Bomba, prof. 30 m, 20 de marzo de 1991 (color blanco con pólipos entre blanco y transparente, muy abundante en fondo arenoso).
Orden TELESTACEA Hickson, 1930
Familia TELESTIDAE Milne Edwards y Haime, 1857
Género Carijoa Müller, 1867
Carijoa riisei (Duchassaing y Michelotti, 1860)
Clavularia rusei Duchassaing y Michelotti 1860, p.34 (St. Thomas)-Kölliker 1865, p.131 (St. Thomas).
Clavularia riisei Duchassaing y Michelotti 1864, p.23 (St. Thomas).
Telesto riisei Verrill 1870, p.372- Hargitt y Rogers 1901, p.278 (Puerto Rico)- Laackmann 1909, p.78 (St. Thomas, St. John, Dry Tortugas)- Bayer 1959, p.3 (Surinam, Guayana francesa)- Rodríguez 1959, p.274- Bayer 1961, p.39 (Curasao, Venezuela, Jamaica, Puerto Rico, St. Lucia, Guayana francesa, Río de Janeiro: Brasil)- González-Brito 1970a, p.63 (Puerto Rico)- González-Brito 1970b, p.80 (Venezuela)- Rees 1972, p.115 (Puerto Rico)- Manjarrés 1979, p.24 (listado para Islas del Rosario, Colombia)- Bardales 1981, p.38 (Puerto Rico).
Carijoa riisei Bayer 1981, p.906- Humann, 1993, p. 83.
Diagnóstico: Carijoa con canales longitudinales en la pared corporal muy notorios, espículas alargadas y varillas ramificadas semifusionadas. Armadura antocodial débil compuesta de aproximadamente dieciséis hileras longitudinales y ocho septas e interseptas. Coloración de los pólipos exertos blanca y la colonia viva desde blanco hasta naranja. En alcohol color blanco o café claro.
Descripción: Las muestras analizadas presentan formas densamente ramificadas y arborescentes (USNM No. 92965; INVEMAR OCTO-288), arborescentes pero con ramas muy cortas y rastreras (USNM No.92963; INV. 287), todas generalmente partiendo de estolones comunes a la colonia. Las colonias no superaban los 18 cm de altura y generalmente se encontraron entre 10-12 cm. Los cálices laterales con 1-5 mm de largo y los tentáculos del pólipo permanecen semiexertos en las muestras conservadas en alcohol con una coloración blanca. El antocodio se dispone en ocho hileras hacia la base de los tentáculos y ocho septales bajo estos. Las varillas antocodiales (figura 2b, c, h) varían entre 0.08-0.4 mm de largas, las de mayor talla presentan procesos espinosos y curvaturas. Las espículas de la pared del cuerpo presentan 2 formas: varillas largas y curvas con procesos espinosos (figura 2a, d, g) generalmente entre 0.2-0.6 mm de largo siendo más grandes en la muestra colectada en Salmedina (USNM No.92965; INV. OCTO-288), y espículas gruesas y cortas entre 0.1-0.35 mm de largo con 0.05-0.09 mm de grueso (figura 2e, f, i) con tubérculos y procesos complejos de formas irregulares, las espículas más pequeñas (figura 2f, i) pueden encontrarse algunas veces semifusionadas. Las muestras vivas se encontraron generalmente de color naranja a rojo a excepción de la muestra del canal de Bocachica (USNM 92963; INV. OCTO-286) que presentó una coloración entre blanca a naranja pálido.
Comentario: Aunque las espículas de Carijoa riisei tienen cierta afinidad con C. operculata, esta última contiene una armadura antocodial con corona y collar muy desarrollados lo cual diferencia claramente estas dos especies. Carijoa riisei y C. rupicula se han considerado como la misma especie, sin embargo, C. rupicula contiene gran cantidad de espículas fusionadas en las paredes del cuerpo. Cabe notar que las muestras colectadas sobre arrecifes coralinos en Salmedina y Capurganá presentan similitudes con algunas formas de crecimiento de C. rupicula cuando estas se encuentran sobre paredes y muelles sumergidos, pero lamentablemente a causa de que fueron fijadas en seco no se pudo tener una posición clara para incluirlas dentro de esa especie (Bayer, com. per). La muestras analizadas al microscopio se observaron generalmente junto con espículas de poríferos y aunque siempre se encontraron muy asociadas con esponjas exteriormente, sectores limpios presentaban estas espículas en especial hacia las regiones coloreadas de naranja, y no se descarta la posibilidad de algún tipo de asociación con esponjas del género Mycale, como se ha encontrado en octocorales estoloníferos del Indopacífico (Van Soest y Verseveldt, 1987).
Hábitat: Esta especie se encuentra en una gran diversidad de habitats, tipos de substrato y formas de crecimiento. Se le halló en infralitoral rocoso frente a la muralla de Cartagena y en los espolones de Marbella en la misma ciudad, en aguas con alta energía del oleaje y generalmente turbias entre 0.5-2 m de profundidad, habitando dentro de cavidades sombreadas de rocas en compañía de abundantes esponjas e hidroides, donde formaba densos tapetes de colonias cuya orientación estaba relacionada con la dirección del oleaje de manera similar a lo observado por Rees (1972). En el canal navegable de Bocachica en la isla de Tierra Bomba a 30 m, en cercanías al Bajo Mangles en las isla de San Bernardo del Viento a 25 m y en la región de Santa Marta frente al aeropuerto Simón Bolívar a 15 m, se encontró como algo muy raro a nivel mundial en telestáceos, hallarlos sobre fondos arenosos llanos, donde eran muy abundantes y se expandían por estolones irregulares. Se presentó también en arrecifes coralinos bajo corales de plato y oquedades arrecifales en isla del Tesoro en las islas del Rosario (talud externo norte, 20 m), el banco de Salmedina (20 m) y Capurganá (15 m), y dentro de barcos hundidos en Punta de Betín (Santa Marta) a 8 m y en el banco de Salmedina a 2 y 25 m. González-Brito (1970b) y Rees (1972) encuentran esta especie adherida a raices de Rhizophora mangle y ambientes lagunares muy someros.
Distribución: Océano Atlántico: Surinam, Río de Janeiro y Bahía en Brasil; Mar Caribe: Jamaica, Puerto Rico, Antillas, Curaçao, Venezuela y Colombia en toda la costa Caribe desde: la ensenada de Capurganá (golfo de Urabá) bajo Mangles (islas de San Bernardo), Cartagena, isla del Tesoro (islas del Rosario), isla de Tierra Bomba, Banco de Salmedina, Santa Marta (Punta de Betín y plataforma continental frente al aeropuerto Simón Bolívar) y península de la Guajira (figura 1).
Material examinado: USNM No.92963; INVEMAR OCTO-286: Una colonia; Canal de Bocachica, isla de Tierra Bomba-Bahía de Cartagena, prof. 30 m, 20 de marzo de 1991 (color naranja pálido con pólipos blancos, creciendo sobre estolones irregulares sobre arena). USNM No.92964; INV. OCTO-287: Una colonia; costa paralela a la ciudad amurallada de Cartagena hacia el Caribe, prof. 1.5 m, 6 de noviembre de 1990 (color naranja, con pólipos blancos, sobre oquedades rocosas en un ambiente de alta energía del oleaje). USNM No.92965; INV. OCTO-288: Una colonia; Banco de Salmedina, al SW de Cartagena; talud externo de barlovento en 27 m de prof, 26 de julio de 1991 (Color naranja con pólipos blancos, bajo corales de forma de plato de Agaricia agaricites y Montastraea annularis) Ident. F.M. Bayer. INV. OCTO-291: Una colonia; Punta de Betín, Bahía de Santa Marta, dentro de un barco hundido, 5 m prof. 15 de marzo de 1994, asociado a Mycale sp. de color naranja. INV. OCTO-292-3: Dos colonias; Plataforma continental frente al aeropuerto Simón Bolívar, Santa Marta, mar Caribe, 15 m prof, marzo de 1994, sobre fondos arenosos, pesca de arrastre, B/I ANCON. INV. OCTO-294: Una colonia, de la península de la Guajira entre Cardón y Ahuyama (11°56'58" N; 72°16'18" W), prof: 10 m, 9 de septiembre de 1994 B/I ANCON (sobre un parches denso de gorgonáceos entre arena y cascajo).
DISCUSIÓN
Siendo el área estudiada relativamente extensa y con numerosas formaciones coralinas, se esperaba encontrar más especies de los órdenes Telestacea y Pennatulacea, ya que estudios sobre Santa Marta registran cerca de 37 especies de gorgonáceos y en las islas del Rosario y Barú más de 40, presentes generalmente sobre arrecifes coralinos (Botero, 1987; Conde y Londofio, 1991; Sánchez, 1993; Sánchez y Ramírez, 1994). Sin embargo, un patrón similar ocurre en todo el mar Caribe, en donde el mayor número de octocorales está dentro del orden Gorgonacea, contando toda la provincia del Atlántico occidental aproximadamente con 90 especies y formas contra tan solo 7 especies de telestáceos, 6 de penatuláceos y 4 alcyonáceos (Bayer, 1959; 1961; Verseveldt y Bayer, 1988), mientras en el Indopacífico se encuentran cerca de 70 especies de alcyonáceos (Sammarco y Coll, 1992), pero el número de especies de gorgonáceos no es superior al Atlántico occidental, y el de telestáceos y penatuláceos es similar.
Estas interesantes diferencias están posiblemente ligadas a la historia geológica de las dos regiones, la gran extensión del océano Pacífico y sus largos períodos de estabilidad climática, facilitaron la radiación específica de los alcyonáceos, los cuales soportan un estrecho rango climático y por ende de latitud. Por otro lado, los fuertes cambios climáticos ocurridos en la evolución del océano Atlántico causaron probablemente la extinción de muchas especies de alcyonáceos, mientras que órdenes que toleran diferentes temperaturas y por ende altas latitudes, como los gorgonáceos, tuvieron una relativa rápida colonización y radiación específica (Klopfer, 1959; Sammarco y Coll, 1992).
Para el caso de los penatuláceos, que han habitado el Atlántico occidental desde el terciario, presentando pocas diferencias entre las especies actuales con sus registros fósiles (Bayer, 1955), es posible que gracias a su amplia distribución latitudinal (Bayer, 1961) hayan soportado los fuertes cambios climáticos ocurridos en las épocas geológicas de manera similar que los gorgonáceos, permaneciendo así en esta región. Sin embargo, la razón de su escasa especiación sigue siendo desconocida, presentándose un caso similar con el orden Telestacea, el cual mundialmente contiene tan solo dos familias conocidas.
Excluyendo el orden Gorgonacea, los octocorales que se han registrado hasta el momento en la costa Caribe colombiana son: el alcyonáceo Nidalia rubripunctata Verseveldt y Bayer (frente a Carrizal en la Guajira en 79-82 m de profundidad: Verseveldt y Bayer, 1988); el penatuláceo Stylatula diadema Bayer (25-30 m de prof, siendo su primer registro para el Mar Caribe) y los telestáceos: Carijoa operculata Bayer (Santa Marta, 15 m prof: Botero, 1987) y C. riseii (Duchassaing and Michelotti), en una extensa área comprendida entre el Golfo de Urabá y la península de la Guajira (0-30 m).
AGRADECIMIENTOS
Un especial agradecimiento al Dr. Frederick M. Bayer del Instituto Smithsonian de Washington D.C. por su invaluable ayuda A la Facultad de Biología Marina de la Universidad de Bogotá Jorge Tadeo Lozano seccionales del Caribe y Santafé de Bogotá. Al Centro de Investigaciones Oceanógraficas e Hidrográficas de la Armada Nacional (CIOH). A Jaime Garzón-Ferreira, Leonor Botero y un revisor anónimo por sus críticas. Al Instituto de Investigaciones Marinas de Punta de Betín-INVEMAR, Santa Marta. A Juan Manuel Díaz y Francisco Borrero del proyecto "Evaluación ecológica y prospección pesquera preliminar de los bancos naturales de ostra perlífera (Pinctada imbricata y Pieria colymbus) en el norte del Caribe colombiano" durante el cual se colectó el material de la Guajira. A la empresa de buceo El Caracol, Cartagena.
BIBLIOGRAFÍA
1 Bardales, A.T. 1981. Reproductive patterns of three species of octocorals (families Telestidae, Briareidae, Plexauridae) in the vicinity of la Parguera, Puerto Rico. MSc. Thesis. Univ. Mayaguez, Puerto Rico, 86p. [ Links ]
2 Bayer, F.M. 1955. Remarkably preserved fossil sea-pens and their recent counterparts. Jour. Wash. Acad. Scl., 45(9): 294-300. [ Links ]
3 Bayer, F.M. 1956. Octocorallia: 166-231. En R. Moore (ed) Treatise on invertebrate paleontology. Part. F. Coelenterata. Geol. Soc. America- Univ. Kansas Press. [ Links ]
4 Bayer, F.M. 1957. Aditional records of Western Atlantic Octocorals. Journ. Wash. Acad. Sci., 47: 379-390. [ Links ]
5 Bayer, F.M. 1959. Octocorals from Surinam and the adjacent coasts of South America. Stud. Fauna Curacao and other Guyanas, 6: 1-43. [ Links ]
6 Bayer, F.M. 1961. The shallow water Octocorallia of the west Indian region. Stud. Fauna Curacao, 12(55): 1-373. [ Links ]
7 Bayer, F.M. 1981. Key to the genera of Octocorallia exclusive of Pennatulacea (Coelenterata: Anthozoa) with diagnosis of a new taxa. Proc, Biol. Soc. Wash., 94:902-947. [ Links ]
8 Botero, L. 1987. Zonación de octocorales gorgonáceos en el área de Santa Marta y Parque Nacional Tayrona, costa Caribe colombiana. An. Inst. Inv. Mar. Punta de Betín, 17:61-60. [ Links ]
9 Conde, S. y R, Londono, 1991. Inventario preliminar de gorgonáceos (Cnidaria: Anthozoa, Octocorallia) en el Bajo del Medio y otras localidades de las islas del Rosario Cartagena, Colombia. Mem. VII. Sem. Nal. Cienc, y Tee. Mar, 316-324. [ Links ]
10 Duchassaing, P. y J. Michelotti. 1860. Memoire sur les coralliaires des Antilles. Mem. Reale Accad. Sci. Torino, 2(19): 279-365. [ Links ]
11 Duchassaing, P. 1864. Suplement au memorie sur les corallaires des Antilles. Mem. Reale Accad. Sci.Torino. 2(23): 97-206. [ Links ]
12 González-Brito, P. 1970a. Una lista de los octocorales de Puerto Rico. Carib. J. Sci., 10(1-2): 63-69. [ Links ]
13 González-Brito, P. 1970b. Algunos octocorales de la isla de Margarita, Venezuela. Bol. Inst. Oceanogr. Univ. Oriente, 9(1-2): 79-92. [ Links ]
14 Hargitt, O. y C. Rogers. 1901. The Alcyonarian of Porto Rico. Bull. U. S. Fish. Comm., 20 (2): 265- 287. [ Links ]
15 Humann, P. 1993. Reef coral identification. New World Publications, Inc., Jacksonville, Florida. 237 p. [ Links ]
16 Kinzie, R.A. 1973. The zonation of West Indian gorgonians. Bull. Mar. Sci., 23: 93-155. [ Links ]
17 Klopfer, P.H. 1959. Environmental determinants of faunal diversity. Am. Nat., 93: 337-342. [ Links ]
18 Kölliker, R.A. 1865. Die bindesubstanz der Coelenlenteraten. Icones histologicae oder atlas der vergleichenden Gewebelehre. Leipzig, 2(1): 87-181. [ Links ]
19 Laackmann, H. 1909. Zur kenntnis der Alcyonarien-Gattung Telesto Lmx. Zool. Jahr., 1(2): 41-104. [ Links ]
20 Manjarrés, G.A. 1979. Observaciones sobre los Anthozoa (excepto Madreporaria) en las islas del Rosario. An. Ins. Inv. Mar. Punta Betín, 11: 23-24. [ Links ]
21 Rees, J.T. 1972. The efect of current on growth form in an octocoral. J. exp. mar. Biol. Ecol., 10: 115-123. [ Links ]
22 Rodríguez, G. 1959. The marine communities of Margarita island, Venezuela. Bull. U.S. Fish. Comm., 20(2): 265-287. [ Links ]
23 Sammarco, P.W y J.C. Coll. 1992. Chemical adaptations in the Octocoraliia: evolutionary considerations. Mar. Ecol. Prog. Ser., 88: 93-104. [ Links ]
24 Sánchez, J.A. 1993. Las comunidades del bajo arrecifal Imelda, isla Barú, Caribe colombiano. V Estructura de la comunidad de gorgonáceos (Cnidaria: Octocoraliia). Mem. VIII Sem. Nal. Cien. Tec. Mar Santa Marta, Colombia. I: 328-340. [ Links ]
25 Sánchez, J.A. y A. Ramírez. 1994. Descripción, composición y estructura de las comunidades coralinas en los arrecifes de isla del Tesoro, Caribe colombiano. Tesis de grado, Univ. de Bogotá Jorge Tadeo Lozano, Santafé de Bogotá, D.C. 130p. [ Links ]
26 Van Soest, R y J. Verseveldt. 1987. Unique symbiotic Octocoral-sponge association from Komodo. Indo-Malayan Zoology, 4: 27-33. [ Links ]
27 Verrill, A.E. 1870. Contributions to zoology from the Museum of Yale College: 7. Description of new corals. Amer. Jour. Sci. Arts., 2(49): 370-375. [ Links ]
28 Verseveldt, J. y F.M. Bayer. 1988. Revision of the genera Bellonella, Euletherobia, Nidalia, and Nidaliopsis (Octocoraliia: Alcyoniidae and Nidalliidae), with descriptions of two new genera. Zoologische Verhandenlinge, 245: 3-131. [ Links ]
