










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
versão impressa ISSN 0122-9761
Bol. Invest. Mar. Cost. v.25 n.1 Santa Marta jan./dez. 1996
CRECIMIENTO DEL CORAL ACROPORA PALMATA (LAMARCK, 1886) EN EL PARQUE NACIONAL NATURAL CORALES DEL ROSARIO, CARIBE COLOMBIANO
GROWTH OF THE CORAL ACROPORA PALMATA (LAMARCK, 1886) IN THE CORALES DEL ROSARIO NATIONAL NATURAL PARK, COLOMBIAN CARIBBEAN
Rocío del Pilar García U.1, Elvira M. Alvarado Ch.1 y Alberto Acosta M.2
1Museo del Mar - Universidad de Bogotá Jorge Tadeo Lozano, Apartado Aéreo 34185, Bogotá, Colombia (R. G. U. y E. M. A. Ch).
2Museo del Mar - Universidad de Bogotá Jorge Tadeo Lozano, Apartado Aéreo 59629 Bogotá, Colombia (A. A. M.)
RESUMEN
Se determinó la tasa de crecimiento durante 10 meses (septiembre 1992-Julio 1993) para 30 colonias de Acropora palmata en la zona norte del Parque Nacional Natural Corales del Rosario, Caribe Colombiano. Se emplearon tres técnicas de medición: medida manual directa, tinción con el colorante rojo de alizarina y lectura de bandas de crecimiento a través de fotografía de rayos X. El estimativo anual de crecimiento para la especie en el Parque fue de 5.2 ± 1.3 cm y se encontró inversamente correlacionado con el tamaño de las colonias (largo, ancho y alto). El crecimiento presentó una variación espacial, siendo Isla Caleta la estación con menor tasa de crecimiento e Isla Caribarú la de mayor. El máximo crecimiento se encontró en la época seca (diciembre-febrero). La medida manual directa no presentó diferencias significativas con la medida con el rojo de Alizarina. La técnica de rayos X mostró que Acropora palmata no presenta bandas anuales de crecimiento discernibles.
ABSTRACT
The growth rate in ten months (September 1992 - july 1993) for 30 colonies of Acropora palmata along the northern zone of the Corales del Rosario national natural park, Colombian Caribbean, was determined. Three measuring techniques were used: by hand, by staining with alizarin red, and from growth bands by X ray photography. The annual growth estimate for the species in the Park was 5.2 ± 1.3 cm and was found inversely correlated to the size of the colony (lenght, width and height). The growth presented a spatial variation. The smallest growth was found at Caleta Island and the largest at Caribarú Island. The maximum growth rate was observed in the dry season (December-February). No significant differences were found between measurement by hand and by alizarin red. The X-ray technique showed that Acropora palmata does not present discernible annual growth bands.
INTRODUCCIÓN
Acropora palmata (Lamarck, 1886) es una especie dominante frente a la zona de la rompiente en los arrecifes del Caribe. Debido a los ajustes que presenta en su geometría colonial (alineamiento de las ramas en dirección de la corriente y engrosamiento del tronco), resiste la fuerte turbulencia a la que está sometida durante su vida (Schumacher y Plewka, 1981).
En el Parque Nacional Natural Corales del Rosario, Caribe colombiano, las poblaciones de Acropora spp. han registrado mortalidades cercanas al 90% (Ramírez, 1984). En los arrecifes del lado norte dominan los esqueletos muertos de A. palmata y en todo el Parque esta especie tiene tan solo una cobertura viva menor al 5% (Sarmiento et al., 1989) lo cual ha sido atribuido principalmente al uso de dinamita (Ramírez, 1984) y al efecto de alta turbidez, sedimentación (Alvarado et al., 1986) y nutrientes provenientes del Canal del Dique (Alvarado y Corchuelo, 1992).
Se ha demostrado que la tasa de crecimiento de esta especie difiere marcadamente entre localidades y que su morfología varía en respuesta a diferentes factores ambientales y fisiológicos (Dodge y Vaisnys, 1975; Antonius, 1981; Hudson, 1981; Schumacher y Plewka, 1981; Randall, 1981; Cortés et al., 1984; Hughes y Connell, 1986; Bythell, 1990; Jordán, 1992). Existen referencias sobre las tasas de crecimiento de corales en diferentes partes del mundo (Lewis et al., 1986; Shinn, 1966), pero este es el primer estudio realizado en Colombia sobre la tasa de crecimiento de la especie A. palmata.
Durante 10 meses de muestreo en campo, se midió la tasa de crecimiento de 30 colonias ubicadas en tres estaciones diferentes del norte del Parque, por medio de la medida manual directa del largo de las ramas (extensión esqueletal) y, se comparó con el incremento observado a partir de la línea teñida con rojo de alizarina. Adicionalmente, y con el propósito de medir el crecimiento a través de las bandas en A. palmata y contrastar este resultado con los obtenidos con las otras mediciones, se realizaron tomas de rayos X. Para no alterar la población actual de la especie y debido a su alto estado de deterioro en el Parque, la escogencia de las colonias en las estaciones de muestreo estuvo dada por la disponibilidad tanto en tamaño (tallas grandes) como en número.
ÁREA DE ESTUDIO
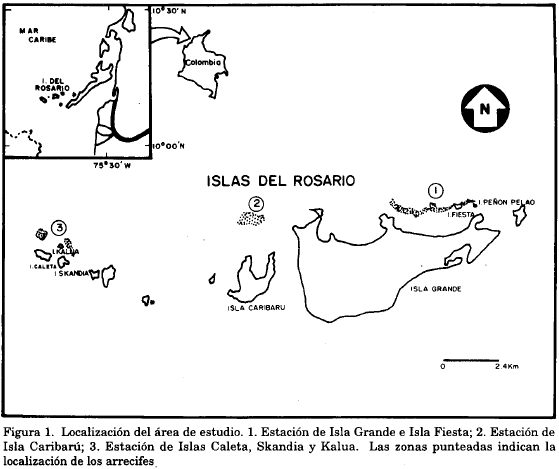
El Parque Nacional Natural Corales del Rosario está situado a 28 millas náuticas al suroeste de Cartagena, Colombia, en un área comprendida entre los 10°08' y 10°15' de latitud norte y entre los 75°40' y 75°48' de longitud oeste (Leblé y Cuignon, 1987) (Fig. 1)
En general el clima de la zona se clasifica como caliente tropical subhúmedo-seco (Duque, 1983). Las épocas climáticas son tres: una seca (de diciembre a abril) dominada por los fuertes vientos Alisios; una época intermedia (de mayo a julio) y una época de lluvia (de agosto hasta noviembre) caracterizada por vientos suaves, en su mayoría de dirección suroeste (Leblé y Cuignon, 1987). La precipitación anual promedio es de 800 mm, la evaporación promedio anual es de 623 mm, la temperatura media anual del ambiente es de 27.7°C y la nubosidad del área es baja con un promedio anual de 5/8 (Alvarado et al., 1989).
El estudio se realizó en tres estaciones del norte del Parque en las cuales se encontraron colonias de A. palmata en buen estado. En Isla Grande y los Islotes de Peñón Pelao e Isla Fiesta (estación 1) la cobertura de coral vivo, era en 1989 de 29.3% y 18% respectivamente; en Isla Caribarú (estación 2), era de 15.1% y en las Islas Caleta, Skandia y Kalua (estación 3), la cobertura de coral vivo era de 10.8%, 4.7% y 13.1% respectivamente (Sarmiento et al., 1989). Los mismos autores señalan para la especie A. palmata un porcentaje de cobertura viva así: 0.19 para el conjunto de islas Caguamo, Bonaire, Pirata, Fiesta y Grande, 0.27% para las Islas Naval, Caribarú, Roberta y Macabí y de 3.6 para las islas Pajarales, Relleno 1, Relleno 2, Majayura, Cubanos, Gloria, Caleta, Kalúa, Skandia, San José, San Juan, San Martín y Yomara.
MATERIALES Y MÉTODOS
A lo largo de la cresta arrecifal se escogieron las mejores colonias teniendo en cuenta que estas presentaran características similares de tamaño y buen estado. Las colonias se marcaron con boyas de icopor y rótulos de acrílico codificados de acuerdo a la estación, tratamiento y número de colonia.
Entre julio y septiembre de 1992 se coloreó con rojo de alizarina una rama de cada una de las 30 colonias (15 de la estación 1; 5 de la estación 2; y 10 de la estación 3) siguiendo la metodología de Brown y Scoffin (1986). A partir del momento de la tinción y hasta julio de 1993 se midió mensualmente, con un metro plástico de costura, el largo a partir de la bifurcación de la rama, el ancho en el punto de bifurcación y el ancho a nivel apical. Igualmente, en cada muestreo se midió el largo y ancho en el plano horizontal y alto en el plano vertical de las colonias que contenían las ramas experimentales. Para los análisis de resultados solo se tomó en cuenta el largo de las ramas (incremento en longitud) ya que el incremento en ancho fue bajo.
Todas las ramas teñidas se colectaron en el último muestreo (julio de 1993), se etiquetaron y se lavaron con agua dulce con el fin de remover el tejido vivo y exponer en el esqueleto, la marca de tinción. Posteriormente se introdujeron en un recipiente con hipoclorito de sodio comercial durante 24 horas y por último se secaron al sol (Guzmán y Cortés, 1989). El crecimiento se tomó como la máxima distancia desde la línea teñida con rojo de alizarina hasta su ápice (Brown y Scoffin, 1986; Guzmán y Cortés, 1989).
Adicionalmente, con el fin de observar la presencia de bandas de crecimiento, las ramas de las colonias marcadas con el rojo de alizarina, una vez fueron medidas, se cortaron con sierra y pulidora en láminas longitudinales con un espesor aproximado de 5 mm (Hubbart y Scaturo, 1985; Guzmán y Cortés, 1989). Estas fueron expuestas a Rayos X a diferentes tiempos de exposición (10 mAs a 40 KvoHs por 0.5 seg, 25 mAs a 40 KvoHs por 0.7 seg y 16 mAs a 42 KvoHs por 0.7 seg). Los mejores negativos de rayos X fueron copiados por contacto directo en papel fotográfico y ampliados al doble para observar las bandas.
Previo al análisis estadístico se seleccionaron solo 16 de las 30 colonias iniciales, por cuanto solo estas tenían un registro completo del crecimiento a lo largo del estudio: 9 de 15 de la estación 1, 4 de 5 de la estación 2, y 3 de 10 en la estación 3.
Teniendo en cuenta los test de homogeneidad de varianzas y normalidad (test de Lilliefors y Shapiro-Wilks), se realizaron los análisis de varianza (ANOVA) de una vía con un nivel de confianza del 95% para probar las hipótesis de igualdad de crecimiento obtenido con la medición directa en campo en las tres estaciones de muestreo, así como la de igualdad de crecimiento obtenido con la técnica de tinción con el colorante rojo de alizarina en las tres estaciones de muestreo. Igualmente se realizó un análisis de varianza a dos vías (ANOVA) de comparaciones pareadas para probar la hipótesis nula de igualdad de crecimiento con las dos técnicas utilizadas.
Con el tamaño promedio total de cada colonia (largo, ancho y alto), así como con los intervalos de tallas y su respectivo crecimiento, se realizó una matriz de correlación para determinar si existía alguna relación directa entre el crecimiento y el tamaño de cada colonia.
RESULTADOS
La tabla 1 registra el incremento promedio total (10 meses) en longitud para cada rama y los promedios por zona a partir de la medida manual directa; el ancho no se incluye debido a que el incremento no fue apreciable. En la estación 1 (Isla Grande e Isla Fiesta) el incremento promedio total del largo de las colonias fue de 4.4 ± 1.8 cm.
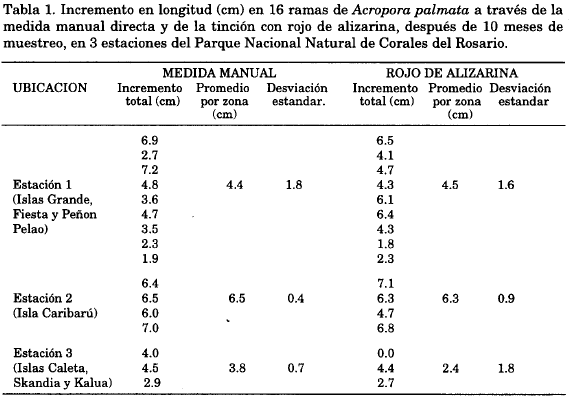
Para la estación 2 (Isla Caribarú) fue de 6.5 ± 0.4 cm y para la estación 3 (Islas Caleta, Skandia y Kalua) fue de 3.8 ± 0.7 cm.
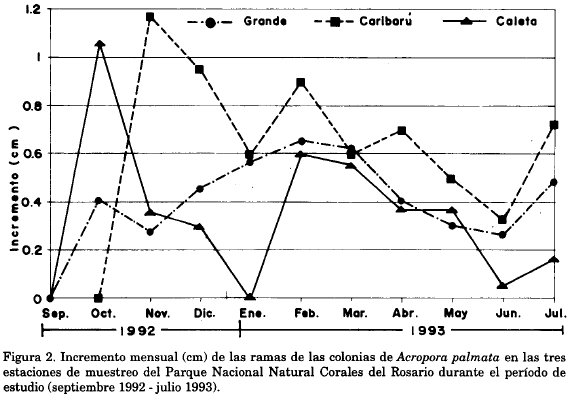
La figura 2 muestra que en los primeros meses de muestreo, el incremento mensual fue diferente y variable para las 3 estaciones. Sin embargo, a partir de febrero se nota un patrón generalizado de cambio en el crecimiento, que va disminuyendo hacia junio y comienza nuevamente a aumentar hacia julio. En esta misma figura se puede notar que la diferencia de crecimiento entre las estaciones coincide con las épocas climáticas. En época de lluvias fuertes (septiembre a noviembre) la variabilidad es mayor que en la época seca (febrero-abril).
Al comparar los valores obtenidos entre estaciones, se observó que las colonias de Caribarú presentaron un mayor crecimiento. No obstante, al realizar el análisis de varianza de una vía (ANOVA, P = 0.0536; F=3.696) no se encontraron diferencias significativas en el incremento del crecimiento entre estaciones, pero como el valor estuvo tan cercano al crítico, se cree que la hipótesis planteada sobre la igualdad de crecimiento entre estaciones, no es del todo aceptable.
Los incrementos del crecimiento del largo de las colonias en las tres estaciones obtenidos a través de la técnica de tinción con el colorante rojo de alizarina, igualmente se observan en la Tabla 1. En la estación de isla Grande e isla Fiesta el incremento promedio total fue de 4.5 ±1.6 cm.
Para las colonias ubicadas en isla Caribarú fue de 6.3 ± 0.9 cm y para las que se encontraban en las islas Caleta, Skandia y Kalua fue de 2.4 ± 1.8 cm. Se pudo observar que el incremento en crecimiento fue mayor en la estación de Caribarú y el menor fue en las islas Caleta, Skandia y Kalua. Esto se comprobó a través del resultado del análisis de varianza de una vía (ANOVA, P=0.0382; F=4.630), y por lo tanto se acepta que existen diferencias en las tasas de crecimiento entre estaciones. Al comparar las zonas, por separado, mediante los análisis de varianza de una vía, hubo diferencia entre las estaciones 2 y 3 (ANOVA, P=0.0268; F=9.63), pero no entre las estaciones 1 y 2 (ANOVA, P=0.0883; F=3.485) y entre las estaciones 1 y 3 (ANOVA, P=0.1059; F=3.158).
Al comparar las dos técnicas utilizadas para medir el incremento del crecimiento de las colonias (medida manual mensual y rojo de alizarina) y una vez examinada la normalización de los datos, se comprobó que con una probabilidad de 0.708 (análisis a dos colas de comparaciones pareadas), no se puede rechazar la hipótesis nula de igualdad en el incremento entre las dos técnicas. Esto valida el uso de la medida manual en especies de rápido crecimiento como A. palmata.
Los intervalos de talla para la especie estuvieron entre 96 y 249 cm de largo, 59 y 183 cm de ancho y entre 30 y 72 cm de alto, con un promedio total de 158.3 x 107.4 x 52.2 cm. La matriz de correlación de las medidas de los tamaños de cada colonia (largo, ancho, alto) en relación con el crecimiento, mostró en un modelo general y uno al tomar los intervalos de tallas (>182 cm; entre 103 y 183 cm y <102 cm), que existe relación inversa entre el largo, ancho y alto de la colonia con el crecimiento de la rama (Tabla 2).
En los análisis de las fotografías de rayos X tomadas a las ramas no se observaron bandas de crecimiento.
DISCUSION
Tasa de Crecimiento
El valor máximo de crecimiento en A. palmata de 7.2 cm, así como el incremento promedio de 4.4 cm en diez meses (equivalente a 8.2 y 5.3 cm /año respectivamente) es comparable en magnitud, aunque menor, con lo registrado para otras regiones del Caribe: Vaughan (1915 en Huston, 1985), obtuvo en la Florida 10 cm/año; Shinn (1972) registró igualmente para la Florida 10 cm/año; Bak (1976), registró en Curazao un incremento medio de 7.4 cm/año y un máximo de 8.8 cm/año; y Gladfelter et al. (1978) obtuvieron en las islas Vírgenes valores entre 4.7 y 8.3 cm/año. Teniendo estos datos en mente, se puede pensar que aunque las condiciones fisicoquímicas del agua en el Parque han sido alteradas en las últimas dos décadas, el crecimiento registrado en el período de estudio (septiembre 1992 - julio 1993) señala que la especie se está manteniendo y está desarrollando sus procesos biológicos.
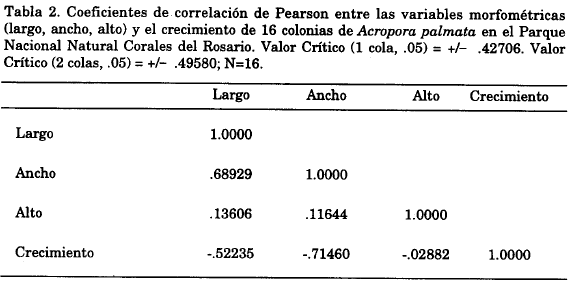
La correlación de las medidas tomadas en cada colonia (largo, ancho y alto) con su crecimiento, evidenció un comportamiento inverso, es decir, que a mayor tamaño, menor crecimiento. Este es concordante con las observaciones realizadas por Hughes y Jackson (1985) quienes afirmaron que el crecimiento de las colonias es dependiente de su tamaño inicial; es decir, que las colonias pequeñas aumentan proporcionalmente más su área de tejido vivo que las colonias grandes. Sin embargo, esta correlación no se puede generalizar para A. palmata, pues no siempre se presenta en la especie; de hecho Bak (1976) indicó que las colonias grandes (viejas) acumulan más rápidamente carbonato que las pequeñas (jóvenes) y por lo tanto aunque la tasa de calcificación total es dependiente del tamaño, es el tamaño de la superficie y no el volumen de la colonia la que es determinante en el crecimiento.
Crecimiento Entre Zonas
El crecimiento de las colonias en las tres estaciones fue diferente. Independientemente de la técnica utilizada (medida manual - rojo de alizarina) se observó que en la estación 2 (Isla Caribarú) se obtuvo el mayor incremento seguido de la estación 1 (Isla Grande e Isla Fiesta) y por último la estación 3 (Islas Caleta, Skandia y Kalua) (Tabla 1). Con los análisis de varianza se confirman estas diferencias para la medida con el rojo de alizarina entre las estaciones 2 y 3. No obstante, dados los valores de desviación estándar se puede pensar que las diferencias en el crecimiento por estación pueden ser debidas a la desigualdad en el número de ramas resultantes en cada estación al final del estudio (mortalidad y pérdida), a las tasas inconstantes de crecimiento (no crecimiento) en algunas ramas en cualquiera de las tres estaciones en diferentes meses, a la mayor o menor exposición al oleaje o, a la ausencia de datos en los primeros meses de muestreo. En cuanto se refiere al menor crecimiento en la estación 3, es probable que sea debido a que solo se tomaron en cuenta tres ramas sobrevivientes y, además, en estas se presentaron meses en los cuales no se observó crecimiento. De acuerdo a las épocas climáticas y a la consecuente incidencia del oleaje sobre el crecimiento, en la figura 2 se puede observar que existe una tendencia a presentar un incremento en los meses de vientos y por consiguiente en los meses de mayor oleaje. Esto es concordante con lo expuesto por Cintrón et al. (1994) quienes afirman que "los corales crecen mejor en aguas agitadas y constantemente renovadas y que por esta razón el mejor crecimiento de los corales ocurre en la vertiente orientada hacia la dirección de donde proviene el viento y el oleaje". La variabilidad en el crecimiento entre estaciones en época de lluvias obedece más a la ausencia de datos en los primeros meses que a la incidencia de las lluvias. Este vacío de información se debió a la imposibilidad de localizar las colonias por la pérdida de las marcas de localización.
Diferencias Entre Técnicas
Las técnicas utilizadas para determinar el crecimiento (medida manual directa mensual y tinción con rojo de Alizarina) fueron igualmente efectivas y no mostraron diferencias significativas. No obstante, las dos técnicas tienen sus ventajas y desventajas. La primera incluye ventajas como la simplicidad y bajo costo del equipo de medición así como la permanencia de las colonias y sus ramas en el parche o arrecife medido. Dentro de las desventajas está la dificultad para realizar la medición a partir de un punto preciso de referencia. Dentro de las ventajas de la medición con rojo de alizarina están la exactitud de la medida a partir de un punto de referencia y la fácil aplicación en campo ya que la incorporación al carbonato fue efectiva y, dentro de las desventajas, están la manipulación de la colonia en el momento de la tinción, el efecto que causa a la rama la aplicación del rojo de alizarina y el sacrificio final de las ramas para la medición.
Por lo tanto, aunque la tasa de crecimiento es difícil de evaluar directamente, las dos técnicas de medición fueron efectivas pues el incremento en extensión del esqueleto de las ramas pudo ser usado como un indicador directo de la suma de nuevo tejido producido. Sin embargo, para futuros estudios se deben tomar ramas de características de calcificación similares, ya que presumiblemente la punta de la rama es el sitio de producción activa de tejido y por lo tanto en cualquier tipo de medición se pueden causar sesgos en su determinación (Gladfelter et al., 1985). Así, para obtener buenos resultados al determinar el crecimiento es aconsejable: 1) utilizar un punto de mareaje preciso e inamovible; 2) escoger ramas con espacios apreciables para evitar que durante las mediciones se presente la fusión con ramas vecinas; 3) tomar más de una rama por colonia para comparar intracolonialmente el crecimiento y 4) tener la precaución de tomar ramas que posean las mismas características de crecimiento apical (puntas coloreadas o no).
Rayos X
La presencia de bandas anuales de crecimiento no fue detectada. En el presente estudio se observaron sitios claros y oscuros en las fotografías de rayos X y, en las copias a papel se leyeron patrones de densidad de pólipos por superficie. Es probable que este hecho obedezca al pequeño tamaño de los fragmentos que se tomaron (10 cm de largo) ya que de acuerdo a los máximos valores de crecimiento reportados para la especie (10 cm/año) y el máximo encontrado en esta especie (7.2 cm) es muy factible que no se alcanzara a observar el anillo del año y por lo tanto tampoco se alcanzaran a observar bandas de años anteriores. Otro factor pudo ser que el ángulo de corte y el grueso de las láminas no fue el apropiado para la observación a través de rayos-X o, finalmente, a que esta especie no presenta bandas de crecimiento, tal como lo afirmaron Weber et al. (1975) y Huston (1985).
CONCLUSIONES
Los registros de crecimiento en Colombia para A. palmata durante 10 meses (4.4 cm en promedio y un valor máximo de 7.2 cm) son comparables, aunque más bajos, con lo registrado para otras áreas del Caribe. El crecimiento de la especie se observa principalmente en el incremento en longitud a nivel apical de la rama y en relación al tamaño de las colonias (largo ancho y alto), se encontró inversamente correlacionado.
Las técnicas de medida manual directa y de tinción con rojo de alizarina son viables y dan resultados que permiten establecer la tasa de crecimiento de manera confiable. Sin embargo, aunque la tinción con el rojo de alizarina presenta una mayor exactitud, los resultados de este estudio permiten recomendar la utilización de la medida manual directa, al menos en poblaciones con una baja cobertura, ya que evita el estrés que causa la tinción en los pólipos, no hace necesaria la extracción y el sacrificio de las ramas medidas y es una técnica de fácil manejo para especies de rápido crecimiento.
AGRADECIMIENTOS
Los resultados de este estudio son parte del proyecto "Crecimiento y regeneración de algunas especies de coral, manglar y fanerógamas marinas en el Parque Nacional Natural Corales del Rosario" financiado por la Universidad Jorge Tadeo Lozano, COLCIENCIAS, el Fondo para la Promoción de la Investigación y la Tecnología del Banco de la República, ECOPETROL, ARSEG, CEINER y el CIOH. Los autores desean agradecer al personal del Museo del Mar, del departamento de Planeación, de Investigaciones Científicas y de la Facultad de Biología Marina (Cartagena) de la Universidad Jorge Tadeo. A Rafa Vieira, José Magardo Blanquisette, Jaime Polanía, Alberto Ramírez, Sven Zea y a los revisores anónimos que ayudaron a mejorar considerablemente los análisis y presentación del manuscrito.
BIBLIOGRAFÍA
1 Alvarado, E. M. y M. C. Corchuelo. 1992. Los nutrientes, la temperatura y la salinidad provenientes del Canal del Dique como factores de deterioro en el Parque Nacional Natural Corales del Rosario (Cartagena - Colombia). Mem. VIII Semin. Nal. Cien. Tecnol. Mar, CCO, Bogotá: 277-287. [ Links ]
2 Alvarado, E. M.; F. Duque.; L. Florez y R. Ramírez. 1986. Evaluación cuantitativa de los arrecifes coralinos de las Islas del Rosario (Cartagena-Colombia). Bol. Ecotrópica, (15): 1-30. [ Links ]
3 Alvarado, E.M.; G. Pinilla y T. León. 1989. Plan de manejo Parque Nacional Natural Corales del Rosario. Inf. Proy., Univ. Jorge Tadeo Lozano - INDERENA, Bogotá, Tomos I y II. [ Links ]
4 Antonius, A. 1981. The "Band" disease in coral reefs. Proc. 4th Int. Coral Reef Symp., Manila 2: 7-14. [ Links ]
5 Bak, R. P. M. 1976. The growth of coral colonies and the importance of crustose coralline algae and bourrowing sponges in relation with carbonate accumulation. Journal Sea Res., 10(3): 285-337. [ Links ]
6 Brown, B. E. y T.P. Scoffin. 1986. Measuring growth rates of reef corals as an indicator of the effects of pollution and environmental disturbance: 12-24. En B. E. Brown (ed.): Human induced damage to coral reefs. UNESCO Rep. Mar. Sci., 40. [ Links ]
7 Bythell, J. C. 1990. Nutrient uptake in the reef -building coral Acropora palmata at natural environmental concentrations. Mar. Ecol. Prog. Ser., 68: 65-69. [ Links ]
8 Cintran, G.; J. R. García y F. Geraldes. 1994. Manual de métodos para la caracterización y monitoreo de arrecifes de coral. WWF, Gland, 67 p. [ Links ]
9 Cortés, J.; M. M. Murillo.; H. M. Guzmán y J. Acuña. 1984. Pérdida de zooxanthelas y muerte de corales y otros organismos arrecifales en el Caribe y Pacífico de Costa Rica. Rev. Biol. Trop., 32(2): 227-231. [ Links ]
10 Dodge, R. E. y J. R. Vaisnys. 1975. Hermatipic coral growth banding as environmental recorder. Reprinted Nat., 258(5537): 706-708. [ Links ]
11 Duque, F. 1983. Parque Nacional Natural Corales del Rosario. Inf. Tec, INDERENA/CIP, Cartagena, 25 p. [ Links ]
12 Gladfelter, E. H.; R. K. Monahan y W. B. Gladfelter. 1978. Growth rates of five reef-building corals in the northeastern Caribbean. Bull. Mar. Sci., 28:720-734. [ Links ]
13 Gladfelter, E. H.; G. Micheol y A. Sanfelier. 1985. Metabolic gradients along a branch of the reef coral Acropora palmata. Bull. Mar. Sci., 28: 728-734. [ Links ]
14 Guzmán, H. M. y J. Cortés. 1989. Growth rates of eight species of scleractinian corals in the eastern Pacific (Costa Rica). Bull. Mar. Sci., 44(3): 1166-1173. [ Links ]
15 Hubbard, D. K. y D. Scaturo. 1985. Growth rates of seven species of scleractinian corals from Cane Bay Salt River, St. Croix, Usvi. Bull. Mar. Sci., 36(2): 325-338. [ Links ]
16 Hudson, J. H. 1981. Response of Montastrea annularis to environmental changes in the Florida Keys. Proc. 4th Int. Coral Reefs Symp., Manila, 2: 233-240. [ Links ]
17 Hughes, T. P. y J. B. C. Jackson. 1985. Population dynamics and life histories of foliaceous corals. Ecol. Monog.,55 (2): 141-166. [ Links ]
18 Hughes, T. P. y J. H. Connell.1986. Population dynamics based on size or age. A reef - coral analysis. Amer. Nat., 129(6): 818-829. [ Links ]
19 Huston, M. 1985. Variation in coral growth rates with depth at Discovery Bay, Jamaica. Coral Reefs; 4: 19-25. [ Links ]
20 Jordan, E. 1992. Recolonization patterns of Acropora palmata in a marginal environment. Bull. Mar. Sci., 5(1): 104-117. [ Links ]
21 Leble. S. y R. Cuignon. 1987. El archipiélago de las Islas del Rosario. Estudio hidrodinámico y sedimentológico. Bol. Cient. C.I.O.H., Cartagena. 7: 37-51. [ Links ]
22 Lewis, J. B.; F. Axelsen.; J. Goodbody.; C. Page y G. Chislett. 1986. Comparative growth rates of some reef corals in the Caribbean. Research Dept. Navi. Biol., Washington, 22 p. [ Links ]
23 Ramírez, G. A. 1984. Ecología descriptiva de las llanuras madreporarias del Parque Nacional Natural Los Corales del Rosario. FEN Colombia, Bogotá, 71 p. [ Links ]
24 Randall, R. H. 1981. Morphologic diversity in the scleractinian genus Acropora. Proc. 4th Int. Coral Reef Symp., Manila, 2:157-164. [ Links ]
25 Sarmiento, E.; F. Flechas y G. Alvis. 1989. Evaluación cuantitativa del estado actual de las especies coralinas del Parque Nacional Natural Corales del Rosario. Cartagena (Colombia). Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá, 187 p. [ Links ]
26 Schumacher, H. y M. Plewka. 1981. The adaptative significance of mechanical properties versus morphological adjustments in skeletons of Acropora palmata and Acropora cervicomis (Cnidaria, Scleractinia). Proc. 4th Int. Coral Reef Symp., Manila, 2:121-128. [ Links ]
27 Shinn, E. A. 1966. Coral growth rate: an environmental indicator. J. Paleontol., 40: 233-240. [ Links ]
28 Shinn, E. A, 1976. Coral reef recovery in Florida and the Persian Gulf. Environ. Geol., 1: 241-254. [ Links ]
29 Weber, J. N., E. W. White y P. H. Weber. 1975. Correlation of density banding in coral skeletons with environmental parameters: the basis for interpretation of chronological records preserved in the coralla of corals. Paleobiology, 1:137-149. [ Links ]
