










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.26 no.1 Santa Marta Jan./Dec. 1997
ESTRUCTURA DE LA TAXOCENOSIS ANNELIDA-MOLLUSCA EN LA REGION DE MINGUEO, GUAJIRA (CARIBE, COLOMBIANO)
STRUCTURE OF THE TAXOCOENOSIS ANNELLIDA-MOLLUSCA IN THE MINGUEO AREA, GUAJIRA (COLOMBIAN CARIBBEAN)
Angela I. Guzmán-Alvis y Oscar David Solano
Instituto de Investigaciones Marinas y Costeras (INVEMAR), A. A. 1016, Santa Marta, Colombia. E-mail: aguzman@invemar.edu.co (A.I.G.A.). odsolano@invemar.edu.co (O.D.S.)
RESUMEN
En una red de 10 estaciones que se ubicaron entre los 6 y 21.5 m de profundidad en la plataforma continental comprendida entre Dibulla y el Río Palomino (Colombia), se hizo una caracterización puntual de las comunidades del macrozoobentos de fondos blandos. En cada estación se hicieron cuatro lances de una draga van Veen, de los cuales uno se destinó para el análisis granulométrico y los tres restantes para el biológico. Los resultados que se presentan, incluyen el setenta y cinco por ciento de los organismos recolectados, comprendiendo los poliquetos el 69%, y moluscos el 6%. Los análisis se hicieron a nivel de familia de los taxa antes mencionados. Con fin de definir las taxocenosis, se realizó un análisis de clasificación entre estaciones y los grupos allí obtenidos se confrontaron en un ordenamiento multidimensional no métrico. Para cada ensamblaje se recalcularon abundancias, número de familias, familias dominantes, características y diversidad con base en las curvas de k-dominancia. Igualmente, se construyeron curvas ABC, con el fin de observar el grado de perturbación en cada ensamblaje. Finalmente, el patrón de distribución biológico hallado se relacionó con variables ambientales como profundidad y tamaños de grano del sedimento, usando la técnica del BIOENV. Los resultados mostraron dos taxocenosis, denominadas Cossuridae y Lumbrineridae dado que estas familias de poliquetos fueron características de cada una. La comunidad Cossuridae estaba asentada en cienos, localizada más lejos de la costa, fue la más diversa y presentó mayor biomasa que la comunidad Lumbrineridae. Esta última, estuvo asentada sobre arenas muy finas y era más somera. Las curvas ABC mostraron que ambos ensamblajes estaban perturbados, se cree que esta perturbación es de origen natural dado que en la zona no existe actividad antrópica. La profundidad y arenas muy finas fueron las variables que mejor explicaron el patrón de distribución espacial hallado.
ABSTRACT
A punctual characterization of the soft-bottom macrobenthos was conducted on the continental shelf between Dibulla and Río Palomino (Colombia). A net of 10 sample sites, 4 van Venn grabs per site, placed in deeps between 6 and 21.5 m, was sampled for this purpose. The results include 75% of the collected organisms, of which 69% were polychaeta and 6% mollusks. Analysis were performed at family level, in order to identify assemblages, a classification analysis between sample sites was performed and the groups there were confronted with the picture obtained from a non metric multidimensional scaling (NMDS). For each assemblage abundance, number of families, family dominance and diversity (K-dominance) were determined. By means of ABC curves the disturbance status of the assemblages was assessed. Finally, the pattern of distribution was related with water column depth and grain size of the sediments by means of the BIOENV technique. Results showed two assemblages called "Cossuridae" and "Lumbrineridae" on the basis of the polychaeta families that characterize them. The "Cossuridae" assemblage occurs in silt sediments offshore. It was the most diverse and showed the highest biomass levels. The Lumbrineridae assemblage occurs in fine sand sediments and shallow waters. The ABC curves analysis suggests that both assemblages are disturbed by natural causes since there is no human activity in the area. Water depth and fine sand were the environmental variables with the highest explanatory power in relation to the spatial distribution of the assemblages.
INTRODUCCIÓN
El estudio del bentos tiene gran importancia en la evaluación de los sistemas acuáticos, con relación a la interacción existente entre éstos organismos y el medio abiótico que los rodea y sirve de hábitat. Además, la poca movilidad y ciclos de vida relativamente cortos de los organismos que lo conforman, los hacen ideales para el estudio de cambios ambientales naturales o inducidos por la actividad humana (Clarke y Warwick, 1994). En un futuro, la Guajira se perfila como el punto de desarrollo de puertos mineros y explotación de recursos energéticos; por tanto un mejor conocimiento de éste tipo de ecosistemas, conducirá a un mejor planeamiento de los estudios de línea base y monitoreo que necesitará éste tipo de obras para el desarrollo de la región.
La mayoría de los trabajos sobre la estructura de comunidades de fondos blandos en el Caribe colombiano se concentran en la plataforma del departamento del Magdalena (García et al., 1990; Guzmán-Alvis y Díaz, 1993; Guzmán-Alvis y Díaz, 1996; Guzmán y García, 1996), conociéndose muy poco sobre otras regiones. Los estudios realizados en la plataforma de la Guajira, son principalmente descriptivos o limitados a determinadas taxocenosis, como los de Bahía Portete (Dueñas, 1988; Díaz, 1992); el estudio de los ostrácodos desarrollado por Ramírez (1983) frente a Riohacha y la caracterización de moluscos de Puyana (1992).
En los últimos años, se ha venido acumulando evidencia sobre estudios que sugieren que para detectar efectos de contaminación en comunidades macrozoobénticas marinas es suficiente trabajar con niveles taxonómicos mayores a especie (Warwick, 1988a, b; Warwick et al., 1990; Ferraro y Colé, 1992a; Agard et al., 1993; Warwick y Clarke, 1993). Guzmán y García (1996), concluyen que donde existen gradientes naturales, se pueden describir patrones en comunidades a niveles taxonómicos mayores a especie. La agregación taxonómica a nivel de familia ha mostrado en los análisis multivariados poca pérdida de información en gradientes de contaminación y naturales, disminuyendo tiempo de procesamiento en laboratorio y costos entre otros.
La presente investigación es una caracterización espacial de la taxocenosis Annelida-Mollusca de fondos blandos encontrada en Mingueo (Guajira) y de su relación con algunas variables ambientales como la profundidad y el tipo de sedimento. Se escogieron estos dos grupos porque constituyen el 72% del total de organismos encontrados y por tratarse de grupos poco móviles, ligados en mayor o menor grado a determinado tipo de sedimento.
ÁREA DE ESTUDIO
Se encuentra ubicada en el departamento de la Guajira, que está sujeto durante casi todo el año a la acción de los vientos alisios del noreste; estos arrastran las nubes hacia el sudoeste, provocando que se presenten períodos de sequía y aridez (IGAC, 1986). La precipitación total anual es inferior a los 500 mm; y sigue el régimen global para la región del Caribe colombiano, que se caracteriza por ser unimodal, es decir, un máximo de lluvias entre septiembre y noviembre (que representa el mayor aporte con un promedio de 55%), y con algunas precipitaciones cortas y moderadas entre junio y agosto (con un aporte de 20%) (IGAC, 1986; Corpes, 1992).
Las corrientes marinas presentan una dirección E-W cuando los vientos alisios soplan persistentemente (período seco); cuando la fuerza del alisio es menor (período lluvioso), esta corriente es contrarrestada por una contracorriente W-E originada a la altura del Golfo de Urabá en el Caribe Suroccidental (Corpes, 1992).
Presenta una temperatura media anual del aire superior a los 24°C. La amplitud térmica anual rara vez pasa de 3.5°C, es decir, que para un mismo sitio las diferencias entre el mes más caluroso y el más frío son de ese valor (IGAC, 1986).
La plataforma continental de la Guajira es la más extensa del Caribe colombiano, alcanzando 64 km en su máxima amplitud; en general los fondos de esta plataforma son arenosos, rocosos, coralinos y de praderas submarinas que hacen que sea un medio propicio para la pesca de arrastre de camarón, langostas y esponjas principalmente (INDERENA, 1977 en IGAC, 1986; Corpes, 1992). La costa de este sector es predominantemente plana y el litoral de tipo arenoso (Corpes, 1992).
MATERIALES Y MÉTODOS
Muestreo y Análisis de Laboratorio
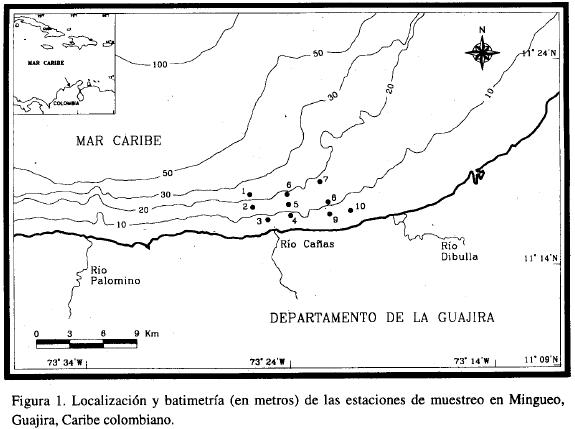
Se recogieron un total de 40 dragados, repartidos en 10 estaciones entre los 6 y 21.5 m de profundidad (Figura 1). En cada estación se tomó un dragado para el análisis granulométrico del sedimento y tres para el análisis biológico (composición, abundancia y biomasa).
Las muestras colectadas para el análisis granulométrico fueron secadas inicialmente a temperatura ambiente y posteriormente en estufa a 80°C hasta obtener un peso constante. Se utilizaron dos metodologías diferentes, dependiendo de si el sedimento era arena o limo. Para el primero se utilizó un tamizado en seco y para el segundo en húmedo (Folk, 1980; Stirn, 1981; Holme y Mclntyre, 1984).
Los dragados destinados al análisis biológico fueron tamizados a través de un cedazo de 0.5 mm, lavando con abundante agua para separar los organismos del sedimento. El material retenido en el tamiz fue preservado en formalina al 5%. Posteriormente, este material fue colocado en una bandeja a fin de separar los organismos de las conchas, restos de material vegetal, restos calcáreos etc., además de agruparlos por taxa mayores. Se identificaron hasta nivel de familia; además, se obtuvieron datos de abundancia (N°. de organismos/ 0.15 m2) y biomasa (gramos/ 0.15 m2) por estación. Esta se determinó con base en el peso seco y en el peso libre de cenizas para cada filum encontrado.
Análisis Matemático
Con base en los conteos realizados se determinaron para cada estación las siguientes características estructurales: número de individuos, número de familias, familia dominante y su porcentaje.
Con el fin de definir las taxocenosis y posteriormente hacer comparaciones entre ellas, la matriz básica se redujo teniendo en cuenta las familias que presentaron porcentajes de dominancia iguales o superiores al 2%. Se realizó un análisis de clasificación entre estaciones, que las agrupa en el espacio, (Field et al, 1982; Crisci y López, 1983), aplicando el índice de Bray-Curtis con transformación de log (x+1). El ligamiento promedio no ponderado (UPGMA) se empleó para unir los grupos (Field et al., 1982; Matteucci y Colma, 1982; Ludwig y Reynolds, 1988; Clarke y Warwick, 1994). Estos grupos se confrontaron con los formados por la técnica de ordenamiento; el análisis utilizado fue el escalamiento multidimensional no métrico (NMDS), partiendo de la matriz de similaridad empleada para el análisis normal (Field et al., 1982; Clarke y Warwick, 1994).
Con el objeto de poder comparar las asociaciones, se recalculó para cada una de ellas la familia dominante y su porcentaje, las familias características, el número promedio de individuos, el número de familias, y la biomasa promedio. Para caracterizar faunísticamente las taxocenosis, se siguió la técnica descrita por Kaandorp (1986) y adicionalmente se recalcularon por taxocenosis la dominancia y frecuencia, así como la dominancia y frecuencia intrafamilia. La diversidad se determinó por taxocenosis, utilizando como herramienta las curvas de k-dominancia (Lambshead et al., 1983); Estas se calcularon con base en la dominancia de todas las familias de poliquetos y moluscos involucrados.
Para estimar perturbaciones naturales o antrópicas en las taxocenosis, se utilizaron las curvas de abundancia y biomasa (ABC) introducidas por Warwick (1986), que comparan simultáneamente la distribución de estos parámetros entre las especies; en el presente estudio las curvas se trazaron con base en la abundancia y biomasa por filum (Warwick, 1988b).
Finalmente, con el propósito de relacionar las variables ambientales con la estructura de la taxocenosis, se aplicó el BIOENV (Clarke y Ainsworth, 1993; Clarke y Warwick, 1994). Las variables tenidas en cuenta fueron: profundidad (P), arena muy gruesa (AMG), arena gruesa (AG), arena media (AM), arena fina (AF), arena muy fina (AMF) y cieno (C). Se construyeron varias matrices triangulares de disimilaridad usando la distancia euclidiana y se relacionaron con la biológica por medio de la correlación harmónica de Spearman. De aquí se obtuvieron una serie de coeficientes que corresponden a la combinación de la(s) variable(s) tomadas en grupos secuenciales de k variables. El máximo coeficiente obtenido de todas las posibles combinaciones indica que ésta es la que 'mejor explica' el patrón biológico obtenido en el ordenamiento (Clarke y Ainsworth, 1993; Clarke y Warwick, 1994).
RESULTADOS
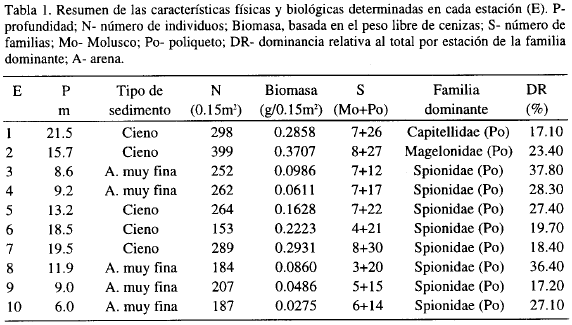
La Tabla 1 resume las características físicas y biológicas de cada estación. En general, se presentaron dos tipos de sedimentos: cienos que dominaron en las estaciones 1, 2, 5, 6 y 7 (más profundas); y arenas muy finas que sobresalieron en las 3, 4, 8, 9 y 10 (que se encuentran más cerca a la costa). El número de individuos por estación varió entre 153 y 399, observándose las mayores abundancias y el mayor número de familias en las estaciones 1, 2, y 7 al igual que las biomasas más altas. Las familias de poliquetos superaron en número a las de moluscos. La familia Spionidae es la que aparece como dominante en la mayoría de las estaciones (Tabla 1).
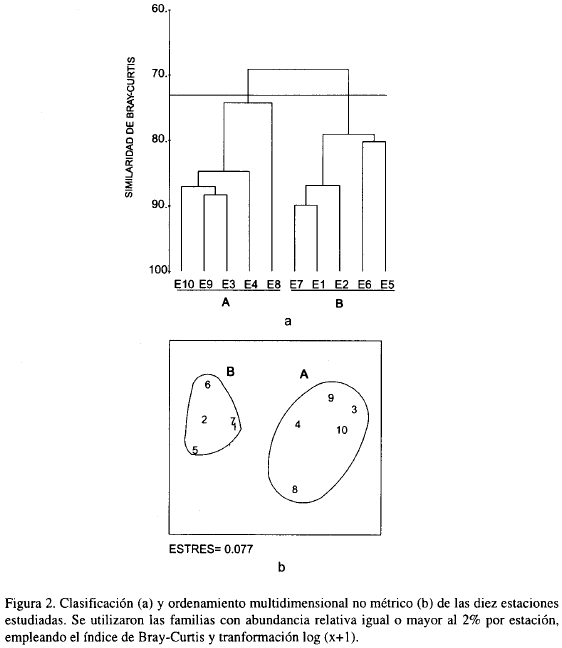
La Figura 2 muestra la agrupación entre las diez estaciones. En la clasificación básicamente se forman dos grupos, el primero constituido por las estaciones 1, 7, 2, 5 y 6 y el segundo por 3, 4, 9, 10 y 8 (Figura 2a). En el análisis de ordenamiento NMDS se establecieron los mismos grupos, con un valor de estrés muy bajo (Figura 2b).
Se encontraron dos taxocenosis, que físicamente se distinguen en profundidad y tipo de sustrato (Tabla 2). La taxocenosis A se establece en arenas muy finas y más cerca a la costa y posee un menor número de individuos, familias y biomasa que la B. Esta última, se asienta en cienos a mayores profundidades y tiene un mayor número de individuos, familias y biomasa con relación a la taxocenosis A.

El análisis inverso muestra la caracterización por familias de la taxocenosis (Figura 3). Se definen como familias características aquellas que tienen un porcentaje de dominancia dentro del grupo superior al 70% (nivel 'g') y frecuencia por encima del 80%. En el grupo principal AB, las primeras once, se encontraron en todas la estaciones, por lo que se denominan familias generalistas; su frecuencia dentro del estudio fue del 100% y también tuvieron densidades promedio altas (Tablas 3 y 4). Las siguientes cuatro familias poseen porcentajes de frecuencia dentro de cada asociación un poco más bajos y densidad promedio dentro de cada taxocenosis muy baja.
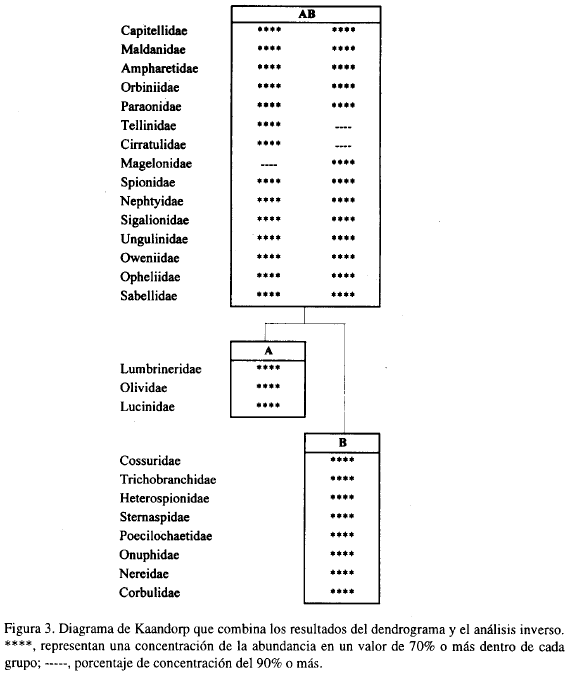
El grupo A tiene como familias características a Lumbrineridae y Lucinidae porque cumplen los porcentajes en dominancia y frecuencias para definirlas como tal; la familia de moluscos Olividae no lo es, puesto que aunque presenta una dominancia del 90% su frecuencia dentro del grupo es del 60% (Tabla 3). Cossuridae, Trichobranchidae, Heterospionidae y Sternaspidae se presentan sólo en el grupo B, por lo que se definen como exclusivas (Tabla 4). Aunque la familia Sternaspidae es exclusiva, su frecuencia dentro de B es muy baja debido a que sólo se encontró una vez, no pudiéndose definir también como característica.
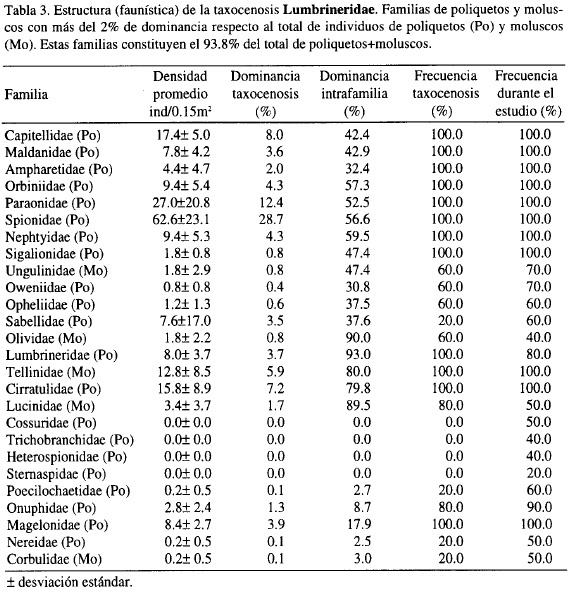
Las demás familias que muestra la Figura 3 para el grupo tienen porcentajes de dominancia intrafamilia y frecuencia por encima del 70% y 80% respectivamente, siendo características de B, al igual que Cossuridae, Trichobranchidae y Heterospionidae (Tabla 4).
La taxocenosis A, denominada en adelante Lumbrineridae, por presentar los mayores porcentajes en dominancia y frecuencia y la taxocenosis B, se denominará Cossuridae.
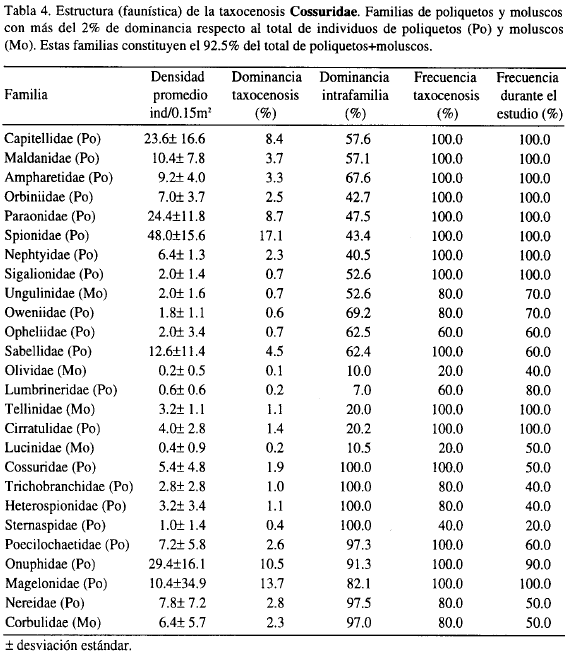
Comparando los valores de diversidad entre las dos taxocenosis se observa que la taxocenosis Cossuridae es más diversa que la taxocenosis Lumbrineridae, porque todos los valores de k-dominancia de ésta siempre se encuentran por debajo de los valores de la taxocenosis Lumbrineridae (Figura 4).
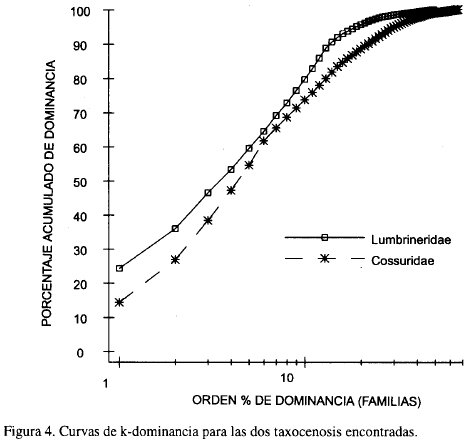
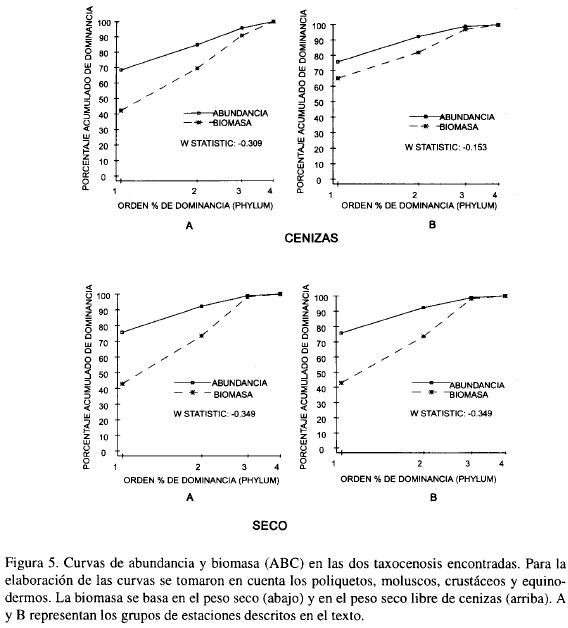
Las curvas ABC (abundancia-biomasa) para las dos taxocenosis encontradas muestran el mismo comportamiento, es decir, las curvas de biomasa se encuentran por debajo de las de abundancia; Los poliquetos constituyen aproximadamente el 70% de abundancia pero presentan biomasas muy bajas (Figura 5). Según modelos propuestos por Warwick (1986), las curvas ABC que siguen este comportamiento corresponden a taxocenosis que se encuentran perturbadas (Warwick, 1986; Clarke, 1990; Agard et al., 1993; Warwick, 1993).
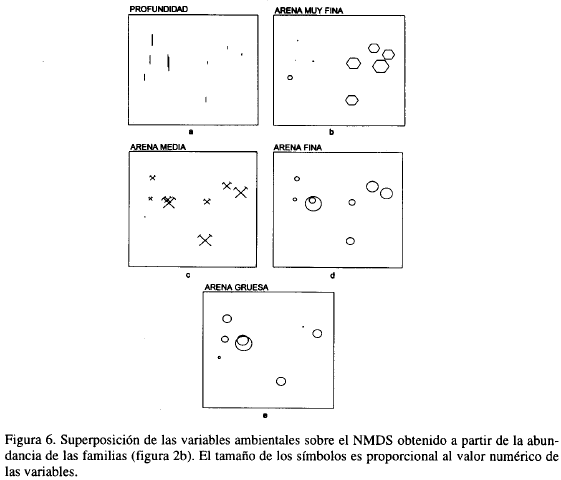
Con el fin de observar gradientes ambientales que estuvieran relacionados con la estructura de las taxocenosis encontradas, se graficaron sobre el NMDS original, y por separado, la profundidad y diferentes tamaños de grano. La Figura 6 muestra claramente que la distribución hallada responde a un gradiente en profundidad y en el contenido de arenas muy finas en los sedimentos.
Para combinar simultáneamente las variables antes mencionadas, se aplicó el BIOENV. Del análisis se excluyeron AMG y C por presentar coeficientes de correlación por encima de 0.95, puesto que éstas no proporcionan información adicional al análisis; además, de acuerdo con los "Draftsman plots", las variables no requirieron ser transformadas (Figura 7). El máximo coeficiente de correlación harmónica se obtuvo para la secuencia profundidad (P)-arenas muy finas (AMF), siendo ésta la combinación que "mejor explica" el patrón de distribución biológico hallado (Tabla 5).

DISCUSIÓN Y CONCLUSIONES
A diferencia del tipo de comunidad que se registra en los trabajos de la plataforma del área de Santa Marta (Guzmán-Alvis y Díaz, 1996; Guzmán-Alvis y Córdoba, 1996), aquí se presentan dos taxocenosis bien definidas, con varias familias características cada una. Se conserva en ambas regiones, un patrón de distribución espacial relacionado con la profundidad y, además, se repiten varias de las familias que son dominantes y frecuentes en ambas comunidades.
La menor diversidad hallada en la taxocenosis Lumbrineridae puede ser explicada de la siguiente forma: Warwick et al., (1990) sugieren que la disminución en la diversidad de la macrofauna puede estar marcadamente influenciada por la inestabilidad del sedimento, esta comunidad presenta estaciones más cercanas a la costa, sometidas a una mayor resuspensión del fondo respecto a aquellas de la taxocenosis Cossuridae que se encuentran a mayor profundidad; la remoción del fondo es producida por los fuertes vientos alisios que inciden en el lugar. Esta inestabilidad física del medio es una posible causa de la disminución en diversidad.
Al observar las curvas ABC se podría pensar que un nivel taxonómico menor (especie, género o familia) podría producir un patrón diferente que el nivel de filum; sin embargo, Warwick (1988a, b) sugiere que cuando existen gradientes de polución o naturales (Ferraro y Colé, 1990b; Guzmán y García, 1996) se reflejan los mismos patrones tanto a niveles taxonómicos altos como bajos, sin pérdida de información. Para el caso de las taxocenosis de Mingueo no se puede hablar de gradientes de polución, puesto que en la zona no hay aparentemente actividad humana que produzca contaminación, pero se pueden argumentar gradientes naturales como la profundidad y el contenido de arenas muy finas como lo muestran las Figuras 6a, b. Igualmente es posible, que el patrón que se observa en las curvas ABC pudiera ser el reflejo de la perturbación que causa el continuo movimiento y resuspensión del fondo producto de los vientos alisios.

Al igual que para otros trabajos realizados en el Caribe colombiano (Tabla 6), la profundidad es importante en la distribución de taxocenosis infaunales; el valor del coeficiente de correlación harmónica es de los más altos en comparación con otros estudios del Caribe. Al cotejarlo con valores de zonas templadas se podría pensar que tiene un poder explicativo bueno, teniendo en cuenta que se encuentra entre los más altos de la tabla. La heterogeneidad de las variables medidas en el presente estudio pueden ayudar a explicar los mayores valores encontrados en comparación con las otras áreas del Caribe colombiano, en donde las variables medidas fueron relativamente homogéneas.
La profundidad influye sobre el tipo de sedimento, originando dos tipos de ambientes en donde se asientan taxocenosis diferentes estructuralmente en diversidad, biomasa y composición de las familias características.
AGRADECIMIENTOS
Agradecemos al Instituto de Investigaciones Marinas y Costeras "José Benito Vives de Andreis" INVEMAR por la financiación de la investigación. Al Dr. J. M. Díaz por la identificación de los moluscos. A la tripulación del B/I Ancón por su colaboración en los muestreos.
BIBLIOGRAFIA
1 Agard. J. B. R., J. Gobin y R. M. Warwick. 1993. Analysis of marine macrobenthic community structure in relation to pollution, natural oil seepage and seasonal disturbance in a tropical environment (Trinidad, West Indies). Mar. Ecol. Prog. Ser., 92: 233-243. [ Links ]
2 Clarke, K. R. 1990. Comparisons of dominance curves. J. Exp. Mar. Biol. Ecol., 138: 143-157. [ Links ]
3 Clarke, K. R. y R. M. Warwick. 1994. Change in marine communities: an approach to statistical analysis and interpretation. Natural Environment Research Council, UK, 141 p. [ Links ]
4 Clarke, K. R. y Ainsworth. 1993. A method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser., 92: 205-219. [ Links ]
5 Crisci, J. y M. López. 1983. Introducción a la teoría y práctica de la taxonomía numérica. OEA, monografía No 26, Washington D.C., 132 p. [ Links ]
6 CORPES, 1992. El Caribe Colombiano: realidad ambiental y desarrollo. Consejo Regional de Planificación de la Costa Atlántica, Bogotá, 350 p. [ Links ]
7 Díaz, J. M. (Ed). 1992. Descripción inicial de unidades de monitoreo de ecosistemas marinos en la Bahía de Portete. Determinación del área de influencia de Puerto Bolívar afuera de la Bahía de Portete. INVEMAR. Informe final, Santa Marta, 67 p. [ Links ]
8 Dueñas, R. 1988. Comunidades bénticas de fondos sedimentarios, 39-75 pp. En: Díaz, J. M. (Ed.). Diagnóstico actual de las comunidades marinas de la Bahía de Portete, análisis de efectos reales para la construcción y operación de las instalaciones portuarias. INVEMAR. Informe final, Santa Marta, 156 p. [ Links ]
9 Ferraro S. y F, Colé. 1990a. Taxonomic level sufficient for assessing a moderate impact on macrobenthic communities in Puget Sound, Washington, USA. Can. J. Fish. Aquat. Sci., 49: 1184-1188. [ Links ]
10 Ferraro S. 1990b. Taxonomic level and sampling size sufficient for assessing pollution impacts on the Southern California Bight macrobenthos. Mar. Ecol. Prog. Ser., 67: 251-262. [ Links ]
11 Field, J.; K. Clark y R. Warwick. 1982. A practical strategy for analysing multispecies distribution patterns. Mar. Ecol. Prog. Ser., 8: 37-52. [ Links ]
12 Folk, R. L 1980. Petrology of sedimentary rocks. Hemphill Publishing Company, Austin, Texas, 182 p. [ Links ]
13 García, C. B.; J. H. Sandoval y H. Salzwedel. 1990. Caracterización puntual de las comunidades macrozoobénticas en la plataforma continental de Ciénaga, Caribe colombiano. Mem. VIII Sem. Nac. Cienc. Tecnol. Mar., Santa Marta, 2: 591-599. [ Links ]
14 Guzmán-Alvis, A. y J. M. Díaz. 1993. Distribución espacial de las taxocenosis Annelida-Mollusca en la plataforma continental del Golfo de Salamanca, Caribe colombiano. An. Inst. Inv. Mar. Punta Betín, 22: 45-59. [ Links ]
15 Guzmán-Alvis, A. y J. M. Díaz. 1996. Soft-bottom macrobenthic assemblages of Santa Marta, Caribbean coast of Colombia. Cahb. J. of Sci., 32:176-186. [ Links ]
16 Guzmán-Alvis, A. y C. B. Garcia. 1996. Taxonomic aggregation and the detection of patterns in a tropical marine benthos data set. Rev. Biol. Trop., 44(2): 865-868. [ Links ]
17 Guzmán-Alvis, A. y M. Córdoba-Tejada. 1996. La comunidad de fondos blandos de la ensenada de Pozos Colorados y áreas aledañas (Caribe, colombiano). Resumen Mem. X Semin. Cienc. Tecnol. Mar, CCO, Santa Fe de Bogotá. [ Links ]
18 Holme, N. A. y A. D. Mclntyre. 1984. Methods for the study of marine benthos. Second edition. Blackwell Scientific Publications, Oxford, 387 p. [ Links ]
19 IGAC, 1986. La Guajira aspectos geográficos. Ministerio de Hacienda y Crédito Público- Instituto Geográfico Agustín Codazzi y la Subdirección de Investigación y Divulgación Geográfica, Bogotá, 151 p. [ Links ]
20 Kaandorp, J. A. 1986. Rocky substrate communities of the infralittoral fringe of the Boulonnais coast, NW France: a cuantitative survey. Marine Biology, 92: 255-265. [ Links ]
21 Lambshead, P.; H. Piatt y K. Shaw. 1983. The detection of differences among assemblages of marine benthic species based on an assessment of dominance and diversity. J. Nat. Hist., 17: 859-874. [ Links ]
22 Ludwig, J. A. y J. F. Reynolds. 1988. Statistical ecology a primer on methods and computing. John Wiley and Sons, New York, 337 p. [ Links ]
23 Matteucci, S. y A. Colma. 1982. Metodología para el estudio de la vegetación. OEA. Monografía No 22, Washington D.C., 168 p. [ Links ]
24 Puyana, M. 1992. Caracterización de la malacofauna de la península de la Guajira (Caribe colombiano). Resumen Mem. VIII Semin. Cienc. Tecnol. Mar. y Congreso Centroamericano y del Caribe en Ciencias del Mar, Santa Marta, Tomo II: 565-580. [ Links ]
25 Ramírez, M. 1983. Contribución al conocimiento de los ostrácodos (Arthropoda-Crustácea) actuales de la plataforma continental frente a Riohacha y su relación con factores del medio ambiente. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá, 135 p. [ Links ]
26 Stirn, J. 1981. Manual of methods in aquatiac environment research. Part 8, Ecological assessment of pollution effects. FAO Fish. Tech. Pap., 209: 1-70. [ Links ]
27 Warwick, R. 1986. A new method for detecting pollution effects on marine macrobenthic communities. Mar. Biol., 92: 557-562. [ Links ]
28 Warwick, R. 1988a. Analysis of community attributes of the macrobenthos of Frierfjord/Langesundfjord at taxonomic levels higher than species. Mar. Ecol. Prog. Ser., 46: 167-170. [ Links ]
29 Warwick, R. 1988b. The level of taxonomic discrimination required to detect pollution effects on marine benthic communities. Mar. Poll. Bull., 19(6): 259-268. [ Links ]
30 Warwick, R. 1993. Environmental impact studies on marine communities: pragmatical considerations. Aust. J. Ecol., 18: 63-80. [ Links ]
31 Warwick, R.; H. M. Plat; K. R. Clarke; J. Agard y J. Gobin. 1990. Analysis of macrobenthic and meiobenthic community structure in relation to pollution and disturbance in Hamilton Harbour, Bermuda. J. Exp. Mar. Biol. Ecol., 138:119-142. [ Links ]
32 Warwick, R. y K. R. Clarke. 1993. Comparing the severity of disturbance: a meta-analysis of marine macrobenthic community data. Mar. Ecol. Prog. Ser., 92: 221-231. [ Links ]
FECHA DE RECEPCIÓN: 11/12/1996 FECHA DE ACEPTACIÓN: 08/07/1997
