










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.27 no.1 Santa Marta Jan./Dec. 1998
ISLA ARENA, UNA FORMACIÓN CORALINA SALUDABLE EN EL ÁREA DE INFLUENCIA DE LA PLUMA DEL RÍO MAGDALENA, PLATAFORMA CONTINENTAL DEL CARIBE COLOMBIANO.
ISLA ARENA, A HEALTHY CORAL FORMATION IN THE AREA INFLUENCED BY THE SEDIMENT PLUME OF THE MAGDALENA RIVER, COLOMBIAN CARIBBEAN CONTINENTAL SHELF.
Jorge H. Pinzón, Adriana M. Perdomo y Juan M. Díaz.
Instituto de Investigaciones Marinas y Costeras (INVEMAR), AA. 1016, Santa Marta, Colombia. E-mail: jmdiaz@invemar.org.co (JMD).
RESUMEN
Se describe la morfología, composición y estructura de la formación coralina desarrollada alrededor de Isla Arena, un pequeño islote localizado en la zona central de la plataforma continental del Caribe colombiano. A partir de fotografías aéreas del área y observaciones en el campo, se elaboraron mapas batimétricos, de las unidades de fondo, de acuerdo al sustrato y a la cobertura de la biota sésil dominante, del grado de exposición al oleaje y de sus patrones de dispersión y refracción. Mediante el empleo de una cámara de video submarina, se filmaron transectos a lo largo de una cadena de 10 m, que fueron analizados usando el método de punto interceptado para la caracterización de la estructura de la comunidad de organismos sésiles. Un total de 33 macroespecies conforman la comunidad bentónica. De éstas, 14 son corales pétreos, cinco son zoantídeos, un octocoral, una anémona, diez macroalgas y dos fanerógamas marinas. Se identificaron diez zonas ecológicas, cada una dominada por una o más especies. La distribución de las zonas está determinada principalmente por el tipo de sustrato y por la energía del oleaje. Las pocas especies de coral encontradas son particularmente resistentes a las condiciones subóptimas imperantes (alta turbidez y turbulencia). Además, el aparente buen estado de salud de los corales, en contraste con otros sectores arrecifales relativamente cercanos, sugiere que éstos puedan también defenderse contra afecciones patológicas cuando están sometidos a condiciones continuas de estrés.
PALABRAS CLAVE: Corales. Sedimentación. Isla Arena. Caribe colombiano.
ABSTRACT
The morphology, composition and structure of the coral community developed around Isla Arena, a tiny shingle cay located on the central area of the Caribbean continental shelf of Colombia, are described. Based on aerial photographs from the area and field surveys, maps of bottom relief, habitat units according to substrate type and cover of dominating sessile biota, wave exposure, and patterns of wave dispersion and refraction were drawn. Employing an underwater videocamera, 10 m-long transects chain were recorded and analized using the plotless method in order to characterize the sessile community structure. A total of 33 species compose the benthic community. Of these, 14 species are stony corals, 5 zoanthids, 1 octocoral, 1 anemone, 10 are macroalgae and 2 sea grasses. Ten ecological zones were identified, each of them dominated by one or more species. The spatial distribution pattern of the zones is mainly controlled by substrate and wave energy. The few coral species found seemed to be particularly adapted to meet the prevailing sub-optimal conditions (high water turbidity and turbulence). Furthermore, their healthy condition suggests that scleractinians growing under chronic stress may also to cope with infestation by common coral diseases when they are continuously subjected to natural stress.
KEY WORDS: Corals. Sedimentation. Arena Island. Colombian Caribbean.
INTRODUCCIÓN
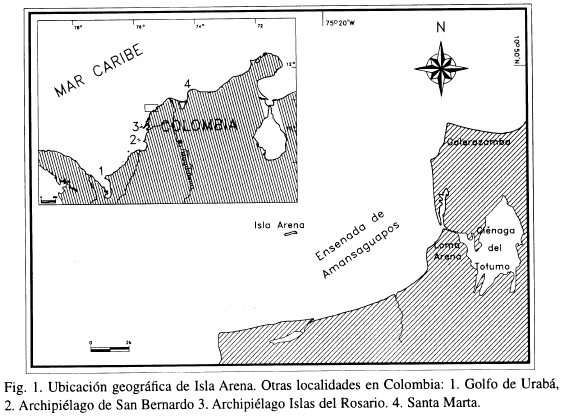
La plataforma y la costa continental del Caribe colombiano presentan varios sectores en los que las condiciones ambientales son favorables para el asentamiento y desarrollo de comunidades coralinas. Agrandes rasgos se distinguen tres sectores en los que se concentran las principales formaciones de este tipo (Figura 1) (Prahl y Erhardt, 1985; UNEP/UICN, 1988; Díaz et al., 1996). El primero está representado por una reducida área de la costa noroccidental del Golfo de Urabá, muy cerca de la frontera con Panamá. El segundo, y más extenso, está constituido por un cordón discontinuo de islas y bajos de origen diapírico, que se extiende por más de 100 km en la porción sur-central de la plataforma colombiana y del cual los archipiélagos de San Bernardo y de El Rosario son los elementos más conspicuos. El otro sector está representado por las formaciones coralinas desarrolladas a lo largo de la costa rocosa del área de Santa Marta. Estos sectores están separados por áreas más o menos amplias cuyas condiciones geomorfológicas e hidrográficas no son aptas para el desarrollo de estructuras hermatípicas. De éstas, la más amplia está fuertemente influenciada por las descargas del río Magdalena, cuyos aportes de sedimentos son el principal factor de control geomorfológico y ecológico en el área (Andrade y Thomas, 1988; Martínez, 1993; Molina et al., 1996).
El reciente hallazgo de una formación coralina relativamente bien desarrollada en torno a un pequeño islote, localizado en un área fuertemente influenciada por la pluma sedimentaria del Río Magdalena, ha motivado la realización de estudios sobre la estructura de la comunidad de organismos sésiles y de los factores que han permitido su desarrollo bajo condiciones aparentemente adversas. En el presente trabajo se describen los rasgos predominantes de la formación coralina, su zonación y estructura comunitaria.
ÁREA DE ESTUDIO
Isla Arena (10°44'20" N - 75°21'10" W) es un pequeño bajo emergido formado por la acumulación de arenas litobioclásticas y escombros calcáreos, localizado sobre la plataforma continental del Caribe colombiano, en la ensenada de Amansaguapos, a unos 8 km de la costa. El cayo tiene forma variable según la época del año en respuesta a la dinámica del viento, el oleaje y las corrientes, y en ocasiones se desagrega en varios cayos conectados por una barra arenosa de escasa profundidad (Figura 1).
Climáticamente, la región se caracteriza por un período lluvioso (junio-noviembre) y una época seca (diciembre-abril). Debido a los vientos alisios del NE, la pluma sedimentaria del Río Magdalena se desplaza hacia el SW durante la época seca (Andrade y Thomas, 1988; Molina et al., 1996), acarreando alrededor de 125 millones de m3 de sedimentos por año (Koopman, 1971), que hacen tránsito a lo largo del litoral y se acumulan en varios sectores (Correa, 1990; Molina et al., 1996). Por esta razón, las aguas en este sector son considerablemente turbias durante la mayor parte del año. El oleaje es generalmente fuerte y está determinado por el régimen de vientos, provenientes predominantemente del NNE, con alturas aproximadas de 2 m (Martínez, 1993; Molina et al., 1996).
Antepuesta a una línea de paleoacantilados de origen tectónico, la costa adyacente es baja, arenosa, construida por la acción del oleaje sobre los sedimentos transportados por la deriva litoral. La geomorfología de la costa evidencia una alta dinámica litoral, que se manifiesta en la formación de tómbolos, flechas arenosas y acrecimiento sedimentario en las playas (Correa, 1990; Molina et al., 1996).
La plataforma continental en este sector es más bien amplia (35 km), de escasa pendiente, y los fondos, con excepción del área en torno a Isla Arena, están predominantemente constituidos por arenas finas litoclásticas. Alrededor de Isla Arena se presentan en forma dispersa algunos bloques calcáreos de origen coralino, lodolitas y sedimentos de grano medio a fino no consolidados.
METODOLOGÍA
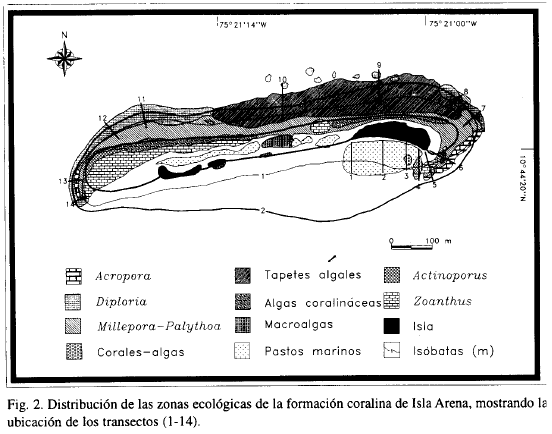
En el transcurso de dos visitas al área de Isla Arena, en septiembre y octubre de 1996, se hicieron observaciones generales acerca de la morfología, composición y zonación de las formaciones coralinas. La cobertura de los componentes bióticos sésiles fue cuantificada aplicando el método de punto intersectado cada 50 cm a lo largo de una cadena de 10 m de largo (Loya, 1976). Los transectos fueron registrados en video con una cámara submarina. La contabilización de la cobertura se efectuó en el laboratorio frente a un televisor a partir de las imágenes de video. Se tendieron un total 14 transectos perpendiculares a la línea de costa hasta la máxima profundidad en la que se observó la presencia de corales (Figura 2). Las zonas ecológicas fueron determinadas y nombradas de acuerdo a la especie o grupo de organismos dominantes en cobertura (Goreau, 1959; Scatterday, 1974).
Como unidad muestral se consideró los tramos de los transectos que se cruzaban con cada zona previamente definida. De acuerdo a las curvas de diversidad acumulada de Shannon obtenidas, las longitudes de los tramos fueron suficientes para determinar la estructura de la comunidad. Los datos fueron transformados mediante el algoritmo del arcoseno [sin-1 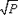 , (P = No. puntos por especie / Total de puntos por tramo)], posteriormente se sometieron a un análisis de clasificación normal mediante la técnica de ligamiento promedio empleando el índice de disimilaridad de Bray-Curtis para construir un dendrograma de similaridad (Field et al., 1982). Una vez obtenidos los grupos, se realizó un análisis inverso mediante la técnica propuesta por Kaandorp (1986), utilizando un 70% de valor acumulado.
, (P = No. puntos por especie / Total de puntos por tramo)], posteriormente se sometieron a un análisis de clasificación normal mediante la técnica de ligamiento promedio empleando el índice de disimilaridad de Bray-Curtis para construir un dendrograma de similaridad (Field et al., 1982). Una vez obtenidos los grupos, se realizó un análisis inverso mediante la técnica propuesta por Kaandorp (1986), utilizando un 70% de valor acumulado.
Los mapas de distribución de unidades ecológicas, topografía del fondo, esquema de dispersión - refracción e intensidad del oleaje fueron elaborados a partir de las observaciones en el campo, la carta de navegación No. 24501 de 1995 y fotografías aéreas pancromáticas del área (Instituto Geográfico Agustín Codazzi, 1974, 1984 y 1991), complementados con fotografías oblicuas tomadas durante un sobrevuelo del área realizado en julio de 1996, y digitalizados en un sistema de información geográfica (SIG-ILWIS) para su análisis. El posicionamiento en el campo se hizo utilizando un instrumento portátil de GPS.
RESULTADOS
En el mapa de la Figura 2 se muestra la distribución espacial de las zonas ecológicas discriminadas de acuerdo a la dominancia de las especies o sustratos y al grado de exposición al oleaje. Las 10 unidades o zonas ecológicas se describen a continuación:
Acropora
Esta zona está dominada ampliamente por una o más especies de corales ramificados del género Acropora [A. cervicornis (hasta 90% de cobertura por Tramo de Transecto (CpTT), A. palmata (hasta 80% de CpTT) y A. prolifera (setos aislados)]; el área total que ocupa esta zona (5114 m2 ) está repartida en cuatro parches discretos, distribuidos hacia la zona periférica a continuación de la zona de Millepora-Palythoa, entre 0 y 0.3 m de profundidad.
Diploría
Esta zona está dominada por amplias colonias de Diploría clivosa y D. strigosa, con abundancias comparables, que cubren el sustrato rocoso a manera de costras hasta en un 87,8% CpTT; esta zona bordea el conjunto de la formación coralina, extendiéndose entre 1.5 y 5 m de profundidad. El parche más extenso se localiza en la parte oeste y ocupa un área de 20300 m2.
Millepora-Palythoa
Los organismos dominantes son el hidrocoral Millepora complánala (30% CpTT) y el zoantídeo Palythoa caribaeorum (50% CpTT), éste último creciendo exuberantemente incluso sobre corales. A excepción de un pequeño parche aislado que se desarrolla a 3 m de profundidad, esta zona se presenta en aguas someras de hasta 1.5 m, extendiéndose principalmente en forma paralela a la línea de rompiente de las olas y ocupando en total un área aproximada de 45057 m2.
Corales-algas
Se caracteriza por la presencia de varias especies de coral (Diploria spp., Millepora complánala, Siderastrea sidérea, Acropora pálmala, Porites porites y P. astreoides), además de zoantídeos (Palythoa caribaeorum, P. mamillosa y Zoanthus sociatus), octocorales (Erythropodium caribaeorum) y macroalgas (Dictyota sp., Avrainvillea sp. y Caulerpa racemosa.), sin que ninguna de ellas sea notoriamente dominante; se presenta en el área de menor turbulencia del agua, del lado oriental de la isla, ocupando un área total de 1591 m2.
Tapetes algales
Las algas filamentosas y algunas macroalgas aisladas recubren densamente los bloques de roca abundantemente distribuidos sobre el fondo al norte de la isla, en profundidades que oscilan entre 0.5 y 3 m; esta zona es la de mayor extensión (61800 m2).
Algas coralináceas
Las algas coralináceas costrosas recubren el sustrato rocoso a lo largo de la cresta del arrecife en un 70% ; aisladamente se observan pequeños parches con colonias de M. complánala, P caribaeorum, Z. sociatus y Z. solanderi; la profundidad no supera 0,5 m y la turbulencia del agua es considerable.
Macroalgas
Diversas especies de macroalgas pardas y verdes conforman praderas poco densas que recubren hasta el 60% CpTT del sustrato, constituido éste primordialmente por gravas aluviales; las macroalgas aparecen entremezcladas o con predominancia de una de ellas, a profundidades no superiores a 1 m; se distinguen varios parches discretos en los que predominan una o dos de las especies presentes; así, la zona que se encuentra al sur de la isla está dominada por Bryopsis sp., mientras que en los dos parches localizados sobre el plano arrecifal dominan Padina sp. y Caulerpa racemosa y en un parche pequeño de la parte oriental domina Dictyota sp.
Pastos marinos
Es una pradera de fanerógamas marinas dominada por Thalassia testudinum que se localiza a sotavento de la isla, en aguas de poca agitación, sobre fondo de arena fina; hacia la parte más profunda de la pradera predomina Syringodium filiforme; el área aproximada de la pradera es de 26.626 m2.
Actinoporus
Es una zona de reducida extensión (algo más de 2000 m2 ), cercana al nivel de marea baja, en la que el sustrato rocoso está ocupado en un 90% por la anémona colonial A. cf. elegans.
Zoanthus
Los zoantídeos Z. sociatus y Z. solanderi son los organismos dominantes, cubriendo en conjunto aproximadamente el 85% del sustrato. La zona, repartida en tres parches, se localiza muy cerca del nivel de marea baja, ocupando un área total de 21100 m2.
En la Figura 3 se muestra el patrón de intensidad y dirección del oleaje; las olas tienden a hacerse paralelas a la línea de costa a medida que se acercan a la cresta del arrecife. Hacia los costados de éste se refractan y forman un núcleo de aguas de poca agitación en el costado de sotavento.
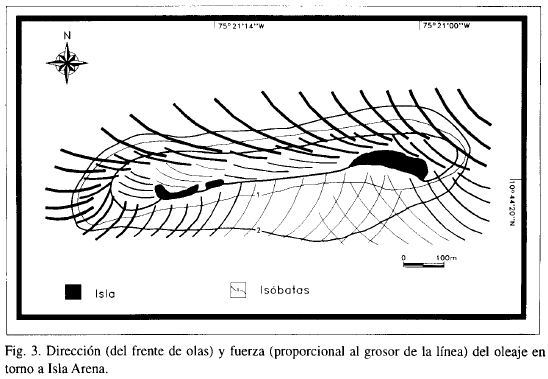
Este esquema de refracción es aparentemente constante a lo largo del año, como se deduce de las fotografías aéreas tomadas en diferentes épocas climáticas; sin embargo, la intensidad del oleaje sí muestra variaciones apreciables, dependiendo de la fuerza de los vientos. Las condiciones de intensidad del oleaje observadas durante las visitas corresponden a las de la época de menor turbulencia.
En la Tabla 1 se hace alusión a la abundancia estimada y a la cobertura relativa promedio de las especies (22) y categorías (5) que fueron cuantificadas en los tramos de los transectos.
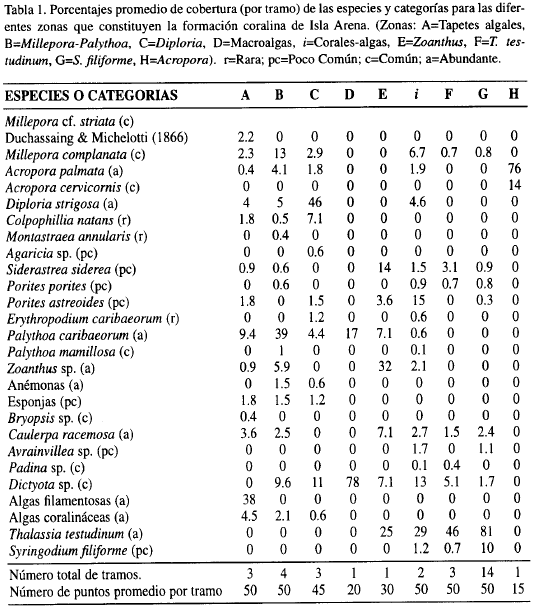
Un total de 33 especies bentónicas sésiles fueron encontradas, de las cuales 12 son corales escleractínios, 2 milleporínidos, 5 zoantídeos, 1 octocoral, 1 anémona, 10 macroalgas y 2 fanerógamas, además de algunas algas costrosas, algas filamentosas y esponjas no identificadas taxonómicamente.
El análisis de clasificación normal (Figura 4) causó el agolpamiento de los tramos en tres grupos y ocho subgrupos, casi todos ellos conformados por los tramos correspondientes a la zona respectiva. El primer grupo (A-D) estuvo constituido por las zonas Tapetes algales (A), Millepora-Palythoa (B), Diploria (C), Corales-algas (i) y Macroalgas (D); el segundo (E-G), lo conforman las zonas Fanerógamas marinas (F-G), Zoanthus (E) y Corales-algas (í); finalmente, el tercer grupo (H) corresponde exclusivamente a la zona de Acropora.
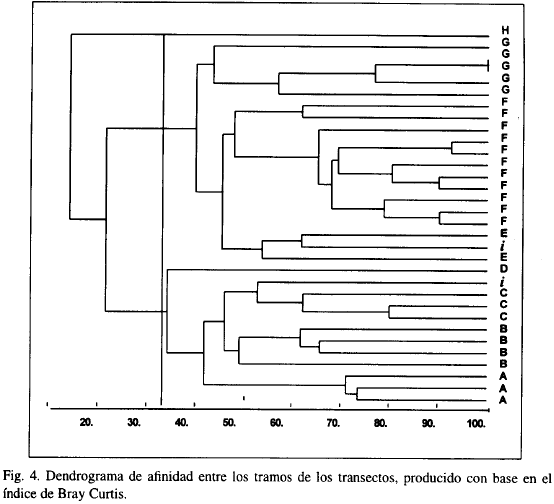
El agolpamiento de los tramos de los transectos en los análisis normal e inverso (Figura 5) deja entrever parcialmente la interferencia entre los factores sustrato y oleaje. Ello es particularmente evidente en el caso de los tramos de la zona Corales-Algas (i), que se encuentran tanto en el subgrupo E como en el subgrupo C, debido a que en esta zona el fondo es una combinación de sustrato rocoso (60 arenoso (40 %), permitiendo la presencia de especies características de ambos tipos de fondo.
Sobre sustrato rocoso consolidado, el orden idealizado de las zonas de acuerdo al grado de exposición es el siguiente (de mayor a menor energía del oleaje): Algas Coralináceas > Millepora-Palythoa, Zoanthus, Actinoporus > Acropora, Diploria > Tapete de Algas > Corales-Algas. La presencia de las zonas de Macroalgas y Praderas de Fanerógamas es claramente dependiente de los sustratos arenosos o de gravas, pero se presentan solamente en los lugares en los que la energía del oleaje es mínima, a sotavento de las áreas emergidas.
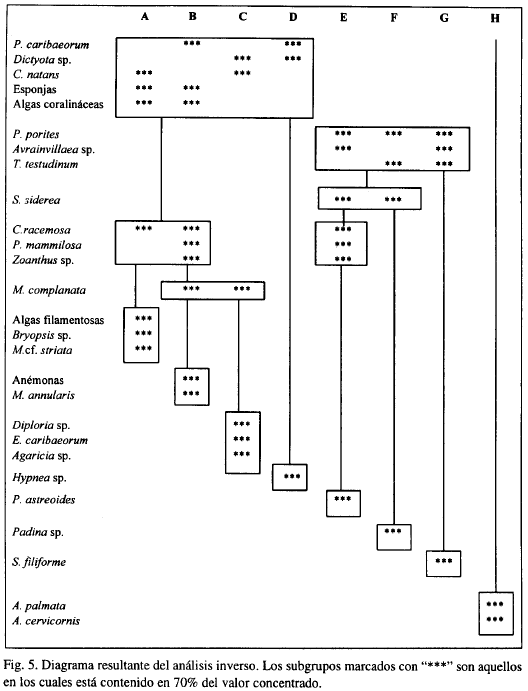
En la Figura 5 se muestra el diagrama resultante del análisis inverso, para definir las especies características se escogió un valor de concentración de la cobertura del 70%. Aquellas que además solo se encontraron en uno de los sub-grupos se les denominó exclusivas.
Para el primer grupo (A-D), las especies exclusivas del subgrupo (A) son algas filamentosas, Bryopsis sp. y M. cf striata, encontradas a barlovento de la isla; este está caracterizado por un sustrato mixto constituido por arena, grava de río y roca, y posee un oleaje moderado. En el subgrupo B las especies características y exclusivas son M. annularis y las anémonas, estas últimas presentándose sobre sustrato rocoso en sitios turbulentos. El subgrupo C está formado por la zona denominada Diploria y por tramos de la zona de Corales-algas (i). Las especies exclusivas allí son Diploria spp., E. caribaeorum y Agaricia sp.; el sustrato es una combinación de arena fina y roca. El subgrupo D tiene como especie característica y exclusiva a la macroalga Hypnea sp., la cual se encuentra en sitios de aguas calmas donde el sustrato predominante es arena fina.
Para el segundo grupo (E-G), el subgrupo (E) tiene como especie exclusiva a P. astreoides. En este subgrupo también se encuentran tramos de la zona de Corales-algas(i). El sustrato es de roca y arena fina, y la turbulencia va de moderada a fuerte. Para el subgrupo F la especie exclusiva es Padina sp., frecuente tanto en zonas de arena fina como de grava. Las aguas son calmas a sotavento de la isla. La fanerógama, S. filiforme, es exclusiva del subgrupo (G), sobre fondos de arena fina y en aguas de menor agitación
En la zona denominada Acropora (H), A. palmata y A. cervicornis son las formas exclusivas, presentes donde el fondo está constituido por roca y arena gruesa y la turbulencia del agua es considerable.
DISCUSIÓN
Es ampliamente conocido que en una misma área la distribución espacial de la biota arrecifal está en gran medida controlada por factores ambientales de naturaleza física y biológica. Junto con la topografía y el tipo de sustrato, los gradientes en el grado de exposición al oleaje y en la energía lumínica son los principales controladores del desarrollo de los diferentes tipos de arrecifes (Sheppard, 1982; Geister, 1983). Así, en los arrecifes del Caribe, determinadas especies forman zonas características que corresponden principalmente a diferentes grados de exposición al oleaje (Geister, 1977, 1982; Graus y Macintyre, 1989).
En el caso de Isla Arena, el efecto combinado del oleaje y del tipo de sustrato parecen ser los factores más determinantes, como se deduce de los agolpamientos resultantes en los análisis normal e inverso. La configuración espacial de las zonas de la comunidad coralina refleja la respuesta del crecimiento de la biota arrecifal preferencialmente en dirección de los vientos y el oleaje predominantes. Mayor oxigenación (turbulencia) y disponibilidad de alimento han sido sugeridos para explicar el mejor desarrollo de la biota hermatípica en el costado de barlovento de pequeñas islas y cayos (Busby, 1966). Igualmente, la pequeña y morfológicamente variable porción emergida del bajo de Isla Arena, compuesta por escombros arrecifales en el costado de barlovento y acumulaciones de arena en el de sotavento, demuestran la naturaleza dinámica de la construcción del cayo.
A pesar de que es evidente una tendencia de las zonas a disponerse en forma paralela, particularmente en el lado de barlovento, debido fundamentalmente al efecto de amortiguación del oleaje con la topografía, algunas de las unidades se presentan en parches discretos, distribuidos en mosaico. Ello se atribuye al efecto combinado de la dispersión-refracción del oleaje y a la distribución heterogénea de los fondos duros y blandos, especialmente en el lado de sotavento.
La relativa reducida cantidad de especies de corales hallada en la comunidad (2 milleporínidos y 12 escleractínios) parece deberse a las condiciones subóptimas para el desarrollo de la mayoría de las especies hermatípicas. La turbidez del agua, y probablemente una elevada tasa de sedimentación durante la mayor parte del año, imponen seguramente restricciones a la fijación de larvas y al desarrollo de los corales. La ausencia de colonias coralinas por debajo de 5 m de profundidad, pese a la presencia de sustratos aparentemente aptos para su fijación y desarrollo, es probablemente consecuencia de ello. Los números de especies registrados en otras áreas coralinas de la plataforma continental del Caribe colombiano son considerablemente más altos. Así, por ejemplo, en los arrecifes que rodean las Islas del Rosario se han registrado entre milleporínidos y escleractínios 51 especies (Pfaff, 1969; Sánchez y Ramírez, 1994), en torno a Isla Fuerte 27 (Díaz et al., 1996a) y en la región de Santa Marta 36 (Antonius, 1972). Inclusive en un área aislada, con pobre desarrollo coralino debido a la carencia de sustratos adecuados, elevada turbidez del agua y aguas de reducida temperatura a causa de una surgencia costera, en el extremo norte de la costa del Caribe colombiano, se ha registrado un mayor número de especies (19) (Solano, 1994).
En la formación coralina que rodea a Isla Arena, como es usual en áreas con condiciones semejantes (Loya, 1976; Cortés y Risk, 1984), los corales han desarrollado mecanismos para contrarrestar el estrés producido por las condiciones adversas. Por ejemplo, Diploria strigosa, D. clivosa, Porites astreoides y Colpophyllia natans son formas particularmente eficientes en la liberación de sedimentos mediante la producción de altas cantidades de mucus, que permiten limpiar continuamente las colonias (Bak y Elgershuizen, 1976). Adicionalmente, las colonias de Diploria suelen desarrollarse como costras recubrentes, aparentemente como estrategia para aprovechar mejor la escasa energía lumínica en condiciones de alta turbidez (Graus y Macintyre, 1982) y para ofrecer mejor resistencia a las fuerzas hidrodinámicas (Vogel, 1981). Ese tipo de crecimiento en Diploria es el predominante alrededor de Isla Arena. En el caso de las especies de Acropora, el crecimiento ramificado y su presencia en zonas de elevada turbulencia impiden la acumulación de sedimento sobre ellas (Bak y Elgershuizen, 1976; Stafford-Smith y Ormond, 1992). El crecimiento de Agaricia sp., en forma de láminas verticales, representa sin duda una estrategia eficiente para contrarrestar la acumulación de sedimentos (Bak y Elgerhuizen, 1976), en tanto que las colonias de Siderastrea sidérea adoptan formas globosas y achatadas en condiciones de alta turbidez (Cortés y Risk, 1984).
Montastraea annularis, una de las especies de mayor abundancia en las zonas de baja turbulencia de los complejos arrecifales oceánicos del Caribe (Geister, 1983; Díaz et al., 1996b), aunque también en los arrecifes coralinos que rodean pequeñas islas y cayos sobre la plataforma en otras áreas del Caribe (Busby, 1966; Rigby y Mclntire, 1966), es poco abundante en la comunidad coralina de Isla Arena, como era de esperarse en un área caracterizada por aguas de poca transparencia y de elevada turbulencia (Geister, 1977; Roberts y Murray, 1983; Cortés y Risk, 1984). Montastraea cavernosa es particularmente frecuente en las formaciones coralinas de la región de Santa Marta (Garzón y Cano, 1991), caracterizada también por aguas relativamente turbias. M. cavernosa está ausente en la comunidad coralina de Isla Arena, debido probablemente a que se distribuye preferencialmente en aguas de reducida turbulencia por debajo de los 5 m de profundidad (Garzón y Cano, 1991).
A diferencia de muchas comunidades coralinas del Caribe, en las que actualmente se registran signos de deterioro avanzado (Ginsburg, 1994; Hughes, 1994) en la comunidad coralina de Isla Arena no se observaron signos de mortalidad coralina reciente que pudiera atribuirse a blanqueamiento o a afecciones de orden patológico, ni siquiera en las especies usualmente más sensibles a estos agentes como A. cervicornis y A. pálmala (Gladfelter, 1982; Díaz et al., 1995). Ello sugiere la posibilidad de que en esta comunidad, desarrollada evidentemente bajo condiciones naturales subóptimas, se ha propiciado la adaptación de las colonias de las pocas especies de escleractínios que la conforman a un cierto nivel crónico de estrés causado por excesiva sedimentación y escasa iluminación, y que debido a ello no han resultado afectadas por las mortalidades masivas de coral ocurridas en las últimas décadas en el Mar Caribe (cf. Ginsburg, 1994). Adicionalmente, esta adaptación parece conferirles resistencia contra los agentes patógenos que afectan comúnmente a ciertas especies de coral en áreas donde las condiciones para su crecimiento son aparentemente óptimas (Garzón-Ferreira y Kielman, 1994; Díaz et al., 1995; Garzón-Ferreira et al., 1996). No se observaron abundancias o coberturas excesivas de algas como tampoco daños físicos de las colonias. La comunidad coralina no ha sido aparentemente afectada de manera significativa por actividades humanas. Isla Arena se localiza relativamente lejos de centros urbano-industriales y, además, la actividad pesquera en sus inmediaciones es aparentemente escasa.
Finalmente, resulta pertinente hacer algunas consideraciones acerca del escenario geológico que propició la formación de esta comunidad coralina. A lo largo de márgenes continentales activas, las arcillas no compactadas del prisma sedimentario pueden estar sujetas a estrés tectónico, provocando flujos verticales de lodo a lo largo de los ejes anticlinales o de los alineamientos de fallas, lo que hace que estas áreas sean altamente susceptibles a afloramientos de arcillas sinsedimentarias y diapirismo (Vernette et al., 1988). Está ampliamente documentado que algunos de los relieves positivos de la plataforma continental del Caribe colombiano se deben a elevaciones, en forma de domo, de arcillas no consolidadas que deforman los sedimentos suprayacentes, originadas por diapirismo de lodo (Vernette, 1989). Estudios de perfiles sísmicos y de alta resolución han mostrado la presencia de domos diapíricos sobre gran parte de la plataforma continental del Caribe colombiano (Shepard et al., 1968; Vernette, 1985,1989). El volcán de lodo de El Totumo y otros rasgos similares en la costa y sobre la plataforma continental frente a Galerazamba, en inmediaciones de Isla Arena, son también manifestaciones inequívocas de actividad diapírica en el área (Vernette, 1989; Carvajal, 1990). Al igual que otras áreas coralinas de la plataforma continental del Caribe colombiano (Islas del Rosario, Bajo Tortugas, Isla Fuerte, Bajo Bushnell; Vernette, 1985, 1989; Díaz et al., 1996a), la formación de una zona de desarrollo coralino en torno a Isla Arena parece haber sido posible gracias a la acumulación de material calcáreo y colonización de la cima de un domo diapírico por parte de la biota hermatípica.
AGRADECIMIENTOS
Los autores expresan sus agradecimientos a S. Zea por su generosa ayuda en el tratamiento de los datos. I. D. Correa, J. H. Carvajal, J. O. Martínez y A. Molina aportaron valiosos comentarios e información acerca de la geología y geomorfología del área de estudio. Al Instituto Alexander von Humboldt, especialmente a A. Repizo, agradecemos por habernos permitido efectuar el sobrevuelo sobre el área para la toma de fotografías. Este trabajo fue posible gracias al apoyo de COLCIENCIAS (proyecto 2105-09-023-93) y del Instituto de Investigaciones Marinas y Costeras, INVEMAR.
BIBLIOGRAFÍA1 Andrade, C. y Y. F. Thomas. 1988. Sedimentos en suspensión e hidrodinámica al sureste del delta del río Magdalena, Mar Caribe, Colombia. Bol. Cient. CIOH, 8: 27-34. [ Links ]
2 Antonius, A. 1972. Occurrence and distribution of stony corals (Anthozoa and Hydrozoa) in the vicinity of Santa Marta, Colombia. Mitt. Inst. Colombo-Alemán Invest. Cient., 6:89-103. [ Links ]
3 Bak, R. P. M. y J. H. B. W. Elgershuizen. 1976. Patterns of oil-sediment rejection in corals. Mar. Biol., 37:105-113. [ Links ]
4 Busby, R. F. 1966. Sediments and reef corals of Cayo Arenas, Campeche Bank, Yucatán, México. U.S. Naval Oceanogr. Off., Tech. Rep., 187: i-xi, 1-58. [ Links ]
5 Carvajal, J. H. 1990. Características sedimentológicas de la plataforma continental frente a Galerazamba. Mem. VII Semin. Nac. Cienc. Tecnol. Mar, CCO, Cali: 164-175. [ Links ]
6 Correa, I. 1990. Inventario de erosión y acreción litoral (1793-1990) entre los Morros y Galerazamba, Departamento de Bolívar, Colombia. Mem. I Sem. Andino de Geología Ambiental, Uribia: 129-142. [ Links ]
7 Cortés, J. y M. J. Risk 1984. El arrecife coralino del Parque Nacional Cahuita, Costa Rica. Rev. Biol.Trop., 32:109-121. [ Links ]
8 Díaz, J. M.; J. Garzón-Ferreira y S. Zea. 1995. Los arrecifes coralinos de la Isla de San Andrés, Colombia: estado actual y perspectivas para su conservación. Acad. Col. Cienc. Exac. Fis. Nat., Col. J. Alvarez Lleras, No. 7, Bogotá, 150 p. [ Links ]
9 Díaz, J. M.; J. A. Sánchez y G. Díaz-Pulido. 1996a. Geomorfología y formaciones arrecifales recientes de Isla Fuerte y Bajo Bushnell, plataforma continental del Caribe colombiano. Bol. Invest. Mar. Cost., 25: 87-105. [ Links ]
10 Díaz, J. M.; G. Díaz-Pulido; J. Garzón-Ferreira; J. Geister; J. A. Sánchez y S. Zea. 1996b. Atlas de los arrecifes coralinos del Caribe colombiano. I, Complejos arrecifales oceánicos. INVEMAR, Serie de publicaciones especiales, 2, 83p. [ Links ]
11 Díaz, J. M.; J. A. Sánchez y J. Geister. 1977. Development of lagoonal reefs in oceanic reef-complexes of the southwestern Caribbean: geomorphology, structure and distribution. Proc. 8th Int. Coral Reef Symp, 1: 779-784. [ Links ]
12 Field, J. G.; K. R. Clarke y R. M. Warwick. 1982. A practical strategy for analysing multispecies distribution patterns. Mar. Ecol. Prog. Ser., 8:37-52. [ Links ]
13 Garzón-Ferreira, J. y M. Cano. 1991. Tipos, distribución, extensión y estado de conservación de los ecosistemas marinos costeros del Parque Nacional Natural Tayrona. Manuscr. Inédito. VII Concurso Nac. Ecol. FEN Colombia, Bogotá, 82 p. [ Links ]
14 Garzón-Ferreira, J. y M. Kielman. 1994. Extensive mortality of corals in the Colombian Caribbean during the last two decades. En R.N. Ginsburg (compilador), Proc. Colloquium on Global Aspects of Coral Reefs : Health, Hazards and History, 1993 : 247-253. [ Links ]
15 Garzón-Ferreira, J; S. Zea y J. M. Diaz. 1996. Coral health assesment in four western Caribbean atolls. 8th Int. Coral Reef. Symp., abstracts: 68 p. [ Links ]
16 Geister, J. 1977. The influence of wave exposure on the ecological zonation of Caribbean coral reefs. Proc. 3rd Int. Coral Reef Symp. Miami: 23-29. [ Links ]
17 Geister, J. 1983. Holozäne westindische Korallenriffe: Geomorphologie, Ökologie und Fazies. Facies, 9:173-284. [ Links ]
18 Ginsburg, R. N. 1994 (Compilador). Proceedings of the Colloquium on Global Aspects of Coral Reefs : Health, Hazards and History, RSMAS, Univ. Miami, 1993. [ Links ]
19 Gladfelter, W. B. 1982. White-band disease in Acropora palmata: implications for the structure and growth of shallow reefs. Bull. Mar. Sci., 32(2): 639-643. [ Links ]
20 Goreau, T. 1959. The ecology of Jamaican coral reefs. I, Species composition and zonation. Ecology, 40:67-90. [ Links ]
21 Graus, R. R. y I. G. Macintyre. 1982. Variation on growth forms of the reef coral Montastrea annularis (Ellis and Solander): A quantitative evaluation of growth reponse to light distribution using computer simulation. Smithson. Contrib. Mar. Sci., 12:441-464. [ Links ]
22 Graus, R. R. y I. G. Macintyre. 1989. The zonation patterns of Caribbean coral reefs as controlled by wave and light energy input, bathymetric setting and reef morphology: computer simulation experiments. Coral Reefs, 8: 9-18. [ Links ]
23 Hughes, T. P. 1994. Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science, 265:1547-1551. [ Links ]
24 Kaandorp, J. A. 1986. Rocky susbtrate communities of the infralittoral fringe of the Boulonnais coast, littordNW France: a quantitative survey. Mar. Biol., 92:255-265. [ Links ]
25 Koopman, B. N. 1971. Interpretación de fotografías aéreas en morfología costera, relacionada con proyectos de ingeniería. Centro Interamericano de Fotointerpretación, Bogotá, 23 p. [ Links ]
26 Loya, Y. 1976. Effects of water turbidity and sedimentation on the community structure of Puerto Rican corals. Bull. Mar. Sei, 26(4): 450-466. [ Links ]
27 Martínez, J. O. 1993. Geomorfología y amenazas geológicas de la línea de costa del Caribe colombiano (Sector Cartagena-Bocas de Ceniza). Publ. geol. esp. Ingeominas, 19:1-62. [ Links ]
28 Molina, A.; A. Pelgrain; J. Suzunaga y L. Giraldo. 1996. Comportamiento de la dinámica marina en el sector costero entre Galerazamba y Cartagena. Bol. Cient. CIOH, 17: 73-78. [ Links ]
29 Pfaff, R. 1969. Las Scleractinia y Milleporina de las Islas del Rosario. Mitt. Inst. Colombo-Aleman Invest. Cient., 3: 16-24. [ Links ]
30 Prahl, H. von. y H. Erhard. 1985. Colombia corales y arrecifes coralinos. FEN-Colombia, Bogotá. 195 p. [ Links ]
31 Rigby, J. K. y W. G. Mcintyre. 1966. The Isla de Lobos and associated reefs, veracruz, Mexico. Brigham Young Univ. Geol. Stud., 13: 3-46. [ Links ]
32 Roberts, H. H. y S. P. Murray. 1983. Controls on reef development and the terrigenous-carbonate interface on a shallow shelf, Nicaragua (Cental America). Coral Reefs, 2: 71-80. [ Links ]
33 Sánchez, J. A. y A. Ramirez. 1994. Descripción, composición y estructura de las comunidades coralinas de los arrecifes de isla Tesoro. Caribe colombiano. Ans. Inst. Invest. Mar. Punta Betín, 24: 55-77. [ Links ]
34 Scatterday, J. W. 1974. Reefs and associated coral assemblages off Bonaire, Netherlands Antilles and their bearing on Pleistocene and Recent reef models. Proc. 2nd Int. Coral Reef Symp, Brisbane: 85-106. [ Links ]
35 Solano, O. 1994. Corales, formaciones arrecifales y blanqueamiento de 1987 en Bahía Portete (Guajira, Colombia). An. Inst. Invest. Mar. Punta Betín, 23: 149-163. [ Links ]
36 Shepard, F.; R. Dill y B. Heezen. 1968. Diapiric intrusions in foreset slope sediments off the Magdalena Delta, Colombia. Bull.Amer. Ass.Petrol.Geol., 52(11): 2197-2207. [ Links ]
37 Sheppard, C. R. C. 1982. Coral populations on reef slopes and their major controls.Mar. Ecol. Prog. Ser., 7: 83-115. [ Links ]
38 Stafford-Smith, M. G. y R. F. G. Ormond. 1992. Sediment-rejection mechanisms of 42 species of Australian scleractinian corals. Aust. J. Mar. Freshwater Res., 43: 683-705. [ Links ]
39 UNEP/IUCN. 1988. Coral reefs of the world Vol. 1, Atlantic and eastern Pacific. UNEP Regional seas directories and bibliographies. IUCN. Gland, Switzerland and Cambridge, UK/UNEP, Nairobi, Kenya, xlvii +373pp. 38maps. [ Links ]
40 Vernette, G. 1985. La plateforme continentale caraibe de Colombie (du debuché du Magdalena au Golfe de Morrosquillo), importance du diapirisme argileux sur la morphologie et la sedimentation. Thèse de Doctorat d'Etat, Univ. Bourdeaux, 387 p. [ Links ]
41 Vernette, G. 1989. Examples of diapiric control on shelf topography and sedimentation patterns on the Colombian Caribbean continental shelf. J. South Amer. Earth Sci., 2(4): 391-400. [ Links ]
42 Vernette, G; G. Lè Blanc; L. Briceño; H. Carvajal; C. Faugeres; J. Gayet; E. Gonthier; R. Griboulard y A. Molina. 1988. Manifestaciones tectónicas en márgenes activas, comparaciones entre dos sectores del Mar Caribe (margen colombiana y prisma de Barbados). Mem. VI Semin. Nac. Cienc. Tecnol. Mar, CCO, Bogotá: 240-252. [ Links ]
43 Vogel, S. 1981. Life in moving fluids. The phisical biology of flow. Princeton University Press, Princeton, XV+352 pp. [ Links ]
FECHA DE RECEPCIÓN: 05/05/1997 FECHA DE ACEPTACIÓN: 29/12/1997
