










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.27 no.1 Santa Marta Jan./Dec. 1998
DINÁMICA POBLACIONAL DEL CHIVO MAPALE CATHOROPS SPIXII EN LA CIÉNAGA GRANDE DE SANTA MARTA Y COMPLEJO DE PAJARALES, CARIBE COLOMBIANO
POPULATION DYNAMICS OF THE MADAMANGO SEA CATFISH CATHOROPS SPIXII AT THE CIÉNAGA GRANDE DE SANTA MARTA AND PAJARALES COASTAL LAGOONS, COLOMBIAN CARIBBEAN
Rocío Tíjaro1, Mario Rueda2 y Adriana Santos-Martínez3
1Instituto de Investigaciones Marinas y Costeras, INVEMAR, A.A. 1016, Santa Marta, Colombia (R.T.).
2CINVESTAV - IPN, Dpto. Recursos del Mar, CP. 97310, A.R 73, Cordemex, Mérida, México. E-mail: mrueda@kin.cieamer.conacyt.mx (M.R.).
3Instituto de Estudios Caribeños, Univ. Nal. Colombia, Hansa Reef 6, San Andrés y Providencia, Colombia (A.S.-M.).
RESUMEN
Durante 1994 el chivo mapalé (Cathorops spixii) fue el segundo recurso pesquero más abundante en la Ciénaga Grande de Santa Marta y el Complejo de Pajarales, principal sistema laguno-estuarino de Colombia. A partir de estadísticas de capturas comerciales y prospecciones pesqueras, se determinaron los parámetros básicos de su población: ciclo reproductivo, crecimiento, mortalidad, rendimiento y biomasa por recluta. Se presentaron individuos maduros durante todo el año con un pico de maduración y desove de junio a agosto cubriendo las épocas de lluvia menor y seca, respectivamente. La talla media de madurez se estimó en 23 cm. Los parámetros de crecimiento de la función de von Bertalanffy fueron: L∞ = 32.5 cm y K = 0.38 año-1, en tanto que el índice de desempeño del crecimiento φ' fue 2.6. Las tasas de mortalidad total (Z), natural (M) y por pesca (F) fueron estimadas en 2.83, 0.96 y 1.87 año-1, respectivamente. La tasa de explotación actual (E = 0.66) indica que el recurso se encuentra sobre-explotado; por consiguiente se recomienda regular la pesquería, reduciendo la tasa de mortalidad por pesca a F = 1.0 año-1 y aumentando la talla media de captura a 23 cm.
PALABRAS CLAVE: Cathorops spixii. Dinámica poblacional. Manejo de recursos pesqueros. Pesca artesanal. Lagunas costeras.
ABSTRACT
During 1994, the catfish "chivo mapalé" (Cathorops spixii) was the second most abundant fishing resource in the "Ciénaga Grande de Santa Marta" and the "Complejo de Pajarales", the main lacustrine-estuarine system of Colombia. Using commercial catch statistics and fishing explorations, the population parameters were determined reproductive cycle, growth, mortality, yield and biomass per recruit. Mature individuals were present all year around with peaks of ripeness and spawning from June to August during periods of little rain and dry, respectively. The mean maturity size was estimated at 23 cm. The growth parameters of the Von Bertalanffy function were: L∞ = 32.5 cm and K = 0.38 year-1, with a growth performance index φ' of 2.6. Total (Z), natural (M) and fishing (F) mortality rates were estimated as 2.83, 0.96 and 1.87 year-1, respectively. The exploitation rate E = 0.66 indicates that the resource is overfished. Therefore it is recomended a fishery regulation by decreasing the mortality rate to F = 1.0 year-1 and increasing the cath mean size to 23 cm.
KEY WORDS: Cathorops spixii. Population dynamics. Management of fishery resources. Artisanal fishing. Coastal lagoons.
INTRODUCCIÓN
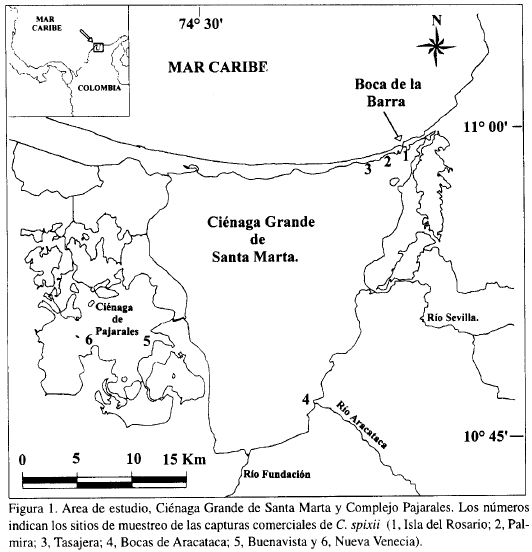
La Ciénaga Grande de Santa Marta (CGSM) y el Complejo de Pajarales (CP) conforman el sistema laguno-estuarino de mayor interés socio-económico de Colombia. Este se encuentra localizado en la costa Caribe colombiana (Figura 1) entre los 10°43' - 11°00' N y 74°16' - 74°35' O (IGACJ973), intercambiando materia y energía con los ríos Sevilla, Aracataca, Fundación, Magdalena y el Mar Caribe. Esta ubicación permite que allí se realicen importantes procesos ecológicos, los cuales generan una alta productividad que ha sustentado el rendimiento alcanzado por la pesquería. Sus aguas presentan una profundidad promedio de 1.6 m (rango de 0.7 a 3.5 m), una temperatura promedio de 30°C (rango de 29 a 32.5 °C) y una salinidad promedio de 26 (rango de 15.3 a 30.8), la cual varía como respuesta a los cambios climáticos que ocurren en dos períodos predominantes al año: uno seco de diciembre a agosto (interrumpido por un corto período lluvioso de mayo a junio) y otro lluvioso de septiembre a noviembre (Botero y Mancera, 1996).
En CGSM y CP, cerca de 20000 personas dependen directamente de la actividad pesquera artesanal, la cual realizan 3215 pescadores con artes y métodos de pesca como atarrayas, redes de enmalle (fijas y bolicheras), palangres, buceo (para ostra y caracol), redes camaroneras y líneas de anzuelos (Mancera y Botero, 1993). Durante 1994 la captura total estimada de peces fue 5335 t, de la cual el 80% estuvo representada por las especies Mugil incilis (32%), Cathorops spixii (25%) y Eugerres plumieri (23%) (Santos-Martínez, 1998), demostrándose así la importancia de la especie en estudio.
C. spixii (Agassiz, 1829), denominada chivo mapalé en CGSM y CP, es una de las especies de peces más importantes, debido a su abundancia y demanda en el mercado local. Se distribuye a lo largo de toda la costa Atlántica de Centro y Sur América desde Belice hasta Brasil; habita en aguas costeras marinas poco profundas, estuarios, lagunas y bocas de ríos, siempre sobre sustrato blando, principalmente fangoso (Fischer, 1978). En algunas zonas como la región nororiental de Venezuela y el delta del Orinoco es sumamente abundante (Cervigón, et al., 1992); es así como Mendoza (1993) reporta parámetros de crecimiento para C. spixii en las costas de este país. En la CGSM, Galvis (1983) realizó algunos estudios sobre taxonomía, reproducción, crecimiento y pesquería, por otro lado Estrada (1987) y Campos (1988) determinaron su importancia como indicadora de contaminación por organoclorados y metales pesados, respectivamente.
C. spixii que veinte años atrás se extraía como fauna acompañante en las capturas de Centropomus undecimalis, Tarpon atlanticus y Mugil liza, actualmente reemplaza a las especies antes mencionadas que ahora son poco frecuentes en las capturas comerciales y sólo se encuentran en estado juvenil (PRO-CIÉNAGA, 1995) como consecuencia del deterioro ambiental del ecosistema y la posible sobrepesca. Teniendo en cuenta que C. spixii es una de las tres especies que sostiene la actividad pesquera de la CGSM y CP, y que existe poca información actual sobre su dinámica poblacional, el objetivo de este estudio es analizar aspectos reproductivos, crecimiento, mortalidad y rendimiento y biomasa por recluta; con el fin de aportar medidas para el manejo sostenible de la pesquería de este recurso.
MATERIALES Y MÉTODOS
Fuentes de información
Para el desarrollo de esta investigación fueron utilizados datos de prospecciones pesqueras y estadísticas de capturas comerciales, recopilados en el marco del "Proyecto Evaluación de los Principales Recursos Pesqueros de la Ciénaga Grande de Santa Marta" realizado entre 1993 y 1998 (Santos-Martínez, 1998).
De cuatro prospecciones pesqueras realizadas en los meses de noviembre de 1993, marzo, junio y agosto de 1994 (Santos-Martínez et al, 1995), se analizaron los datos correspondientes a estados de madurez gonadal de C. spixii y las variables salinidad y temperatura de la superficie del agua. Estas prospecciones correspondieron a cuatro muestreos intensivos diseñados para la estimación de biomasa íctica en la CGSM usando el método de área barrida. Cada una de ellas fue realizada intentando cubrir las diferentes épocas climáticas predominantes y de transición, las cuales han sido identificadas por cambios de la salinidad, que a su vez afectan la fisiología reproductiva de las especies.
Por otra parte, de las estadísticas de capturas comerciales (Santos-Martínez, 1998) tomadas diariamente en los sitios de desembarco (Figura 1), se analizaron los datos de estimaciones de capturas y frecuencias de longitud total de C. spixii entre enero y diciembre de 1994.
Aspectos reproductivos
Un total de 951 individuos de C. spixii fueron capturados en los muestreos de prospección pesquera. El sexo y la madurez gonadal de cinco individuos por clase de talla igual a 0.5 cm, se determinó empleando la escala de Holden y Raitt (1975) modificada por Santos-Martínez (1989), la cual consta de los estados juveniles, indiferenciados, inmaduros, madurando, maduros y desovados. Esta información fue utilizada para determinar la variación temporal en la proporción de los diferentes estados de madurez por sexo, y su relación con la salinidad y temperatura del agua durante el periodo de estudio, con el fin de tener un conocimiento aproximado del ciclo reproductivo de C. spixii. Además se estimó la talla media de madurez sexual por medio de la frecuencia acumulada relativa de individuos en estados maduros y desovados.
Crecimiento
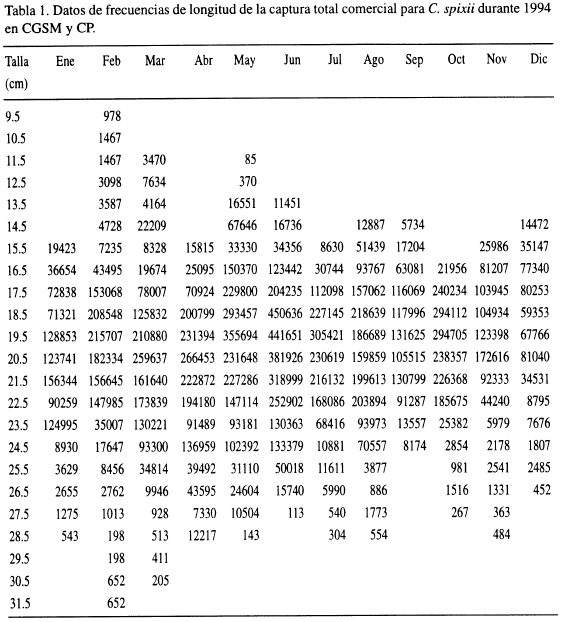
La composición de la captura total por tallas de C. spixii, estimada a partir de muestras de frecuencias de longitud de 9066 individuos (Tabla 1), fue usada para determinar los parámetros de crecimiento, empleando el paquete computacional FiSAT (Herramientas para la Evaluación de Stocks de FAO-ICLARM) (Gayanilo et al., 1993). El crecimiento se asumió al descrito por la función de von Bertalanffy, la cual expresa la talla en función de la edad:
L(t) = L∞ ( 1 - e-k(t-to))
donde Lt es la longitud a la edad t, L∞ es la talla asintótica, K es la constante de crecimiento y, to es la edad hipotética a longitud cero. La rutina ELEFAN I (Pauly, 1987) fue usada para ajustar dos curvas de crecimiento a los datos de frecuencia de longitud reestructurados, utilizando intervalos de tallas de 1 y 2 cm, con el fin de determinar cuál de ellas obtenía el mejor ajuste.
Teniendo en cuenta la correlación existente entre K y L∞, se estimó el índice de desempeño del crecimiento φ' de Pauly y Munro (1984), con el fin de comparar los parámetros de crecimiento de C. spixii con los de otras poblaciones, usando la siguiente expresión:
φ' = log10 K + 21og10 L∞
Estado de explotación y manejo de la pesquería
La tasa de mortalidad total (Z) se estimó usando los datos de frecuencias de longitud de la captura total y los parámetros de crecimiento, mediante el método de la curva de captura linealizada convertida a tallas (Pauly, 1984):
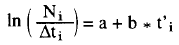
donde Ni es el número de individuos capturados pertenecientes a la clase de longitud i, t'i es la edad relativa de la clase de longitud i y Δti es el tiempo medio requerido por individuo para su crecimiento a través de la clase i.
La tasa de mortalidad natural (M), se estimó usando la ecuación empírica de Pauly (1984):
log10M = 0.0152-0.279*log10 L∞ + 0.6543*log10 K + 0.4634*log10 T
donde L∞ y K son parámetros de la función de von Bertalanffy y T es la temperatura media anual del agua (30 °C para este estudio).
La mortalidad por pesca (F) y la tasa de explotación (E) fueron calculadas por las siguientes ecuaciones respectivamente:
F = Z-M
E = F/Z
La talla media de captura fue estimada a partir de la curva de selección realizada con base en el lado izquierdo de la curva de captura y en un estimado de M con la rutina para estimación de mortalidades de FiSAT.
Se aplicó el modelo de rendimiento y biomasa relativa por recluta de Beverton y Holt (1957) usando la curva de selección logística. Además se determinó una estrategia de manejo buscando el punto óptimo del rendimiento de la pesquería, probando distintas combinaciones de la tasa de explotación y la talla media de captura en las isopletas de rendimiento de FiSAT.
RESULTADOS
Aspectos reproductivos
En las Figuras 2a y 2b se observa la distribución de estados gonadales de C. spixii para machos y hembras, respectivamente; mientras la Figura 2c muestra la variación de temperatura y salinidad en cada una de las cuatro prospecciones pesqueras realizadas.
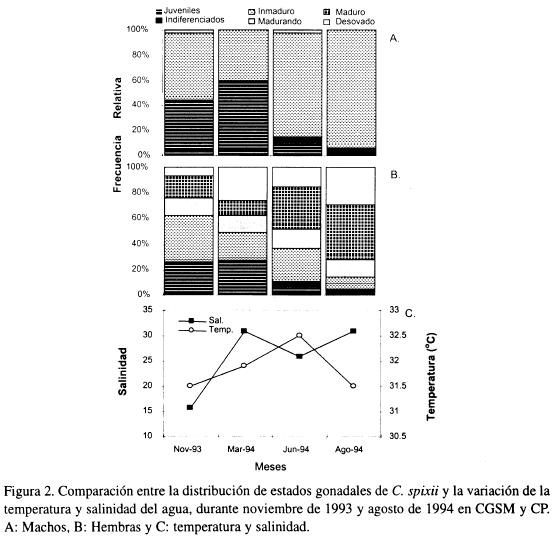
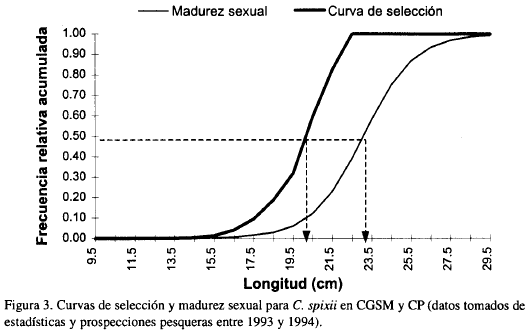
Se destaca que machos maduros y desovados estuvieron ausentes durante todo el periodo de muestreo (Figura 2a), contrario a lo ocurrido con las hembras (Figura 2b) en donde hubo representatividad de todos los estadios. En junio y agosto, cuando la salinidad estuvo por encima de 25, se observó la mayor proporción de hembras maduras y desovadas; mientras en noviembre y marzo, cuando la salinidad cambió de 16 a 31, se presentó la mayor frecuencia de juveniles en ambos sexos. Por otro lado se determinó que la talla a la cual el 50% de la población se encuentra sexualmente madura (talla media de madurez sexual) fue 23 cm (Figura 3).
Crecimiento
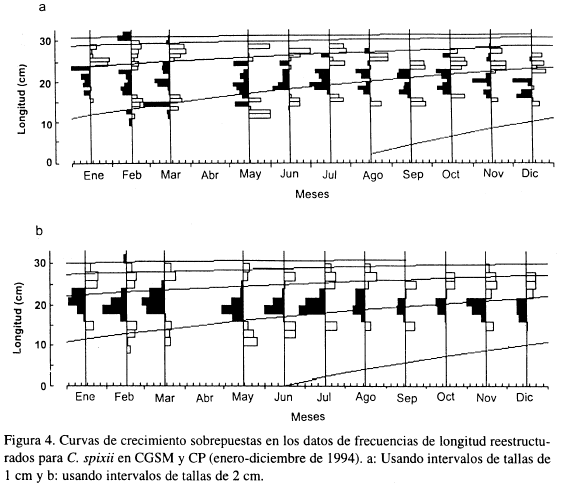
Las Figuras 4a y 4b muestran las correspondientes curvas de crecimiento sobrepuestas a los datos de frecuencias de longitud reestructurados, con intervalos de tallas de 1 y 2 cm respectivamente, provenientes de la composición por tallas de la captura total comercial estimada para 1994.
La Tabla 2 presenta los valores estimados de K, L∞ y φ' obtenidos de la literatura y para cada una de las curvas de crecimiento ajustadas. No existe una progresión modal clara de las frecuencias de longitud, por tanto, la medida de la bondad de ajuste es baja en ambos casos (Rn < 0.2).
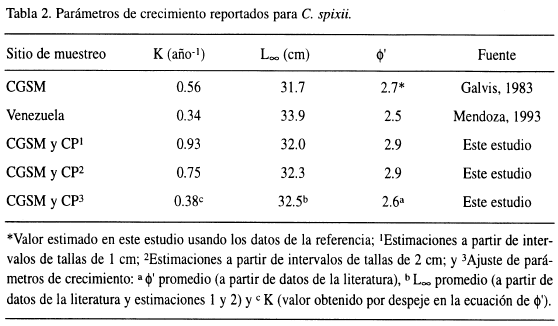
Con el fin de obtener mejores estimados de K y L∞, se recurrió al uso de φ' para promediar los parámetros de crecimiento y luego despejar de la ecuación de φ' el valor de K (K = antilog [(φ' - 21og10 L∞]). De esta manera los valores estimados de K, L∞ y φ' fueron 0.38 año-1, 32.5 cm y 2.6, respectivamente.
Estado de explotación y manejo de la pesquería
De acuerdo a lo observado en la Figura 3 donde se sobreponen las curvas de madurez sexual y de selección de C. spixii, se destaca que la segunda curva está ubicada muy a la izquierda de la primera; además 100% de la selección ocurre a partir de los 22 cm de longitud, talla a la cual sólo un 35% de la población se encuentra sexualmente madura. Tanto la talla media de captura estimada (20 cm), como la talla media de madurez (23 cm), difieren en un rango apreciable.
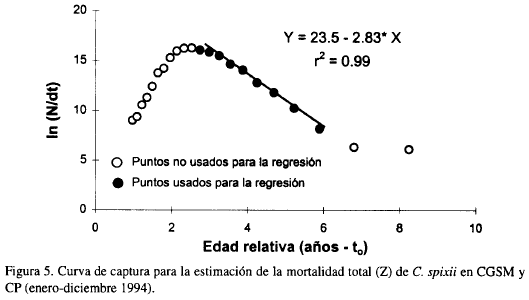
La tasa de mortalidad total obtenida mediante regresión lineal de los puntos ubicados en la fase explotada de la curva de captura fue Z = 2.83 año-1 (Figura 5). La mortalidad natural M se estimó en 0.96 año-1, en consecuencia se obtiene que la mortalidad por pesca fue F = 1.87 año-1 y la tasa de explotación E = 0.66.
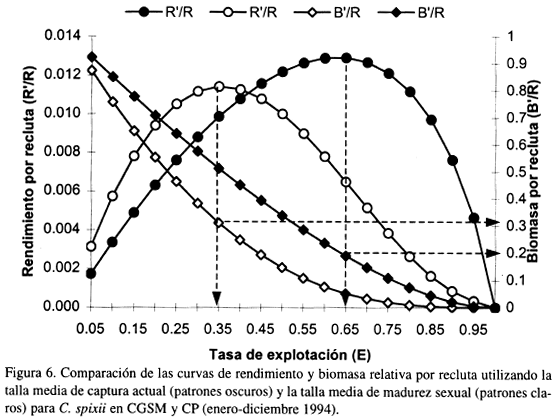
La Figura 6 muestra las curvas de rendimiento y biomasa por recluta del recurso C. spixii bajo dos esquemas diferentes de manejo del recurso, teniendo en cuenta variaciones en la talla media de captura y la tasa de explotación. Las curvas de patrones oscuros, muestran el comportamiento predictivo de la pesquería con base en la actual talla media de captura (20 cm). Para esta talla se obtuvo que la tasa de explotación que permite alcanzar el máximo rendimiento de la pesquería es 0.65, la cual produce un excedente de biomasa del 20%. Por otra parte las curvas de patrones claros, muestran el comportamiento predictivo de la pesquería tomando la talla media de captura igual a la talla media de madurez (23 cm). En este caso, la tasa de explotación que permite alcanzar el máximo rendimiento de la pesquería es 0.35, mientras el excedente de biomasa fue del 32%.
DISCUSIÓN
Aspectos reproductivos
Debido a la presencia de hembras maduras y desovadas en cada muestreo (Figuras 2a y 2b) se puede concluir que C. spixii se reproduce durante todo el año; esto concuerda con lo descrito por Galvis (1983) quien observó material gonadal de la misma especie también en la CGSM. Lo anterior guarda relación con la presencia de juveniles de ambos sexos igualmente en cada muestreo.
La mayor proporción de hembras maduras y desovadas encontradas en junio y agosto durante la época de transición climática lluviosa a seca, podría indicar la época de maduración y desove masivo. Estos eventos reproductivos están relacionados con altos valores de salinidad y no parecen ser afectados por la variable temperatura. De otra parte, la mayor frecuencia de juveniles encontrados en noviembre y marzo, durante la época de transición climática seca a lluviosa, podrían sugerir la ocurrencia de un pico de reclutamiento. Observaciones de juveniles en el borde oriental de la Boca de la Barra en la CGSM durante esta época, realizadas por Gal vis (1983), respaldan lo anterior.
Lo encontrado en este estudio confirma que en los ambientes laguno-estuarinos la salinidad es una de las variables que más incide en los eventos de maduración y desove, igualmente afecta la distribución espacial y temporal de las comunidades ícticas debido a las exigencias fisiológicas que establece el sistema para que pueda ser tolerada (Margalef, 1983; Balogun, 1987; Amadi, 1990).
No obstante, los resultados obtenidos en este estudio aunque basados en un tamaño de muestra suficiente y un rango de tallas representativo de la población, guardan cierto grado de incertidumbre debido a que sólo fue posible tomar cuatro muestras puntuales. Es sabido que para conocer el ciclo reproductivo de una especie, se debe contar con observaciones mensuales de estados de madurez en series de tiempo que cubran un ciclo anual; por tal razón, en este estudio se presenta un conocimiento aproximado del ciclo reproductivo de C. spixii, bajo el supuesto de que cambios en el comportamiento reproductivo de la especie son presentados en cada una de las épocas climáticas en que se realizaron las prospecciones pesqueras.
La talla media de madurez estimada (23.3 cm) es inferior a la reportada por Mancera (1994) de 26 cm, la cual fue calculada a partir de datos obtenidos en 1981 (Galvis, 1983). Esto puede explicarse en el hecho que trece años atrás el ecosistema presentaba condiciones distintas y C. spixii no era sometido a la actual presión de pesca, la cual además de reducir la talla media de captura, pudo ocasionar que la talla media de madurez descendiera en 2.7 cm.
Crecimiento
Como se observa en la Tabla 2, los valores de K estimados en este estudio (0.93 y 0.75) son mayores a los reportados por otros autores (0.56 y 0.34; Galvis, 1983 y Mendoza, 1993, respectivamente), mientras los valores de L∞ (32 y 32.3 cm) estuvieron dentro del rango estimado en la misma literatura (31.7 y 33.9 cm), en tanto el valor de φ' estimado (2.9) estuvo por encima de los calculados en la mencionada literatura (2.7 y 2.5). Una explicación al hecho de que las curvas de crecimiento no hallan presentado una progresión modal clara de las frecuencias de tallas en el tiempo, es que C. spixii es una especie tropical que parece reclutar a la pesquería durante todo el año, lo cual se desprende de las observaciones de juveniles en cada muestreo realizado.
Los parámetros obtenidos usando la relación de φ' (k = 0.38 año-1, L∞ = 32.5 cm y φ' = 2.6) estuvieron dentro de los rangos de la literatura mencionados en el párrafo anterior. Estos parámetros ajustados proporcionan una mayor confiabilidad para conocer el estado de explotación y recomendar medidas de manejo de la pesquería de C. spixii.
Estado de explotación y manejo de la pesquería
Debido a la fuerte pendiente de la curva de selección (Fig. 3) y a que la talla media de captura (20 cm) se encuentra muy por debajo de la talla media de madurez (23 cm), el stock desovante de C. spixii podría estar siendo afectado negativamente ya que se está impidiendo que al menos el 50% de la población madure. Esta situación es consecuencia del empleo de pequeños tamaños de malla en los diferentes artes de pesca usados en la pesquería de la CGSM (Rueda, 1995).
Tomando en cuenta inicialmente la premisa de que el rendimiento óptimo de la pesquería se alcanza cuando F = M, es decir E = 0.5 (Gulland, 1971); se tiene que la población de C. spixii podría encontrarse sobreexplotada, debido a que la tasa de explotación actual es de 0.66.
No obstante, la premisa anterior suele ser poco confiable a la hora de evaluar el estado del recurso, su conclusión coincide con la determinación de diferentes niveles óptimos de explotación a partir del modelo de rendimiento y biomasa relativa por recluta.
En primer lugar si tomamos el esquema de manejo basado en fijar la talla media de captura en 20 cm (patrones oscuros, Fig. 6), se obtuvo que la tasa de explotación óptima (0.65) fue ligeramente menor al valor actual (0.66), lo que quiere decir que actualmente el recurso es sobreexplotado. De aumentar un poco el esfuerzo, la sobrepesca llegará a niveles críticos que afecten el rendimiento, debido a la fuerte pendiente decreciente mostrada después del nivel óptimo, lo que implica un gran riesgo. Además, el nivel excedente de biomasa desovante (20%) es muy poco para garantizar la producción del stock.
Ahora si tomamos el segundo esquema de manejo (patrones claros, Fig. 6), basado en fijar la talla media de captura en 23 cm (talla media de madurez sexual), se determinó que la tasa de explotación óptima es 0.35, la cual corresponde a una pendiente de la curva de rendimiento equivalente a la mitad de la pendiente al origen. Esto confirma que el recurso se encuentra actualmente sobreexplotado y muy posiblemente se presenta una sobrepesca por crecimiento, debido a que los peces están siendo capturados desde los 15 cm, mientras la talla de primera madurez se encuentra alrededor de los 16.5 cm.
De acuerdo al anterior análisis es necesario regular la pesca, aumentando la talla media de captura a 23 cm y disminuyendo la tasa de explotación a 0.35, no implicando esto una disminución drástica del rendimiento. Además, el excedente de biomasa desovante producido (32%), podría garantizar la producción del stock. Para lograr esto es necesario fijar la tasa de mortalidad por pesca en 1.0 año-1 que corresponde a una cuota de captura anual de 735 t con base a las 1375 t de C. spixii capturadas durante el periodo de estudio (Santos-Martínez, 1998). Así mismo, se recomienda usar un tamaño de malla mínimo de 5.7 cm en atarrayas, boliches y redes de enmalle fijas para poder capturar individuos de 23 cm (Rueda y Santos-Martínez, 1997).
En consecuencia el mejor esquema de manejo es el segundo el cual es adverso al riesgo de sobrepesca, ya que la curva de rendimiento posee una pendiente más suave hacia el lado derecho.
AGRADECIMIENTOS
Los autores desean agradecer al Instituto de Investigaciones Marinas y Costeras (INVEMAR), al Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología (COLCIENCIAS) y al Proyecto Colombo-Alemán de Rehabilitación de la Ciénaga Grande de Santa Marta (PRO-CIÉNAGA), por el apoyo financiero y logístico suministrado para la realización del presente trabajo. Igualmente al Dr. Jaime Mendo, por la asesoría suministrada en el enfoque del análisis de la información; a la Bióloga Claudia Sánchez, al Ing. Pesquero Efraín Viloria por sus comentarios y sugerencias, y al técnico Juan Carlos Narváez.
BIBLIOGRAFÍA1 Amadi, A. A. 1990. Comparative ecology of estuaries in Nigeria, Hydrobiologia, 208(1-2): 27-38. [ Links ]
2 Beverton, J. H. and S. J. Holt, 1957. On the dynamics of exploited fish populations. Fish. Invest. Ser., 2(19): 1-533. [ Links ]
3 Balogun, J. K. 1987. Seacional fluctuations of salinity and fish occurrence in and around Epe Lagoon, Nigeria, African Journal of. Ecology, 25(1): 55-61. [ Links ]
4 Botero, L. y J. E. Mancera. 1996. Síntesis de los cambios de origen antrópico ocurridos en los últimos 40 años en la Ciénaga Grande de Santa Marta (Colombia). Rev. Acad. Colomb. Cienc, Vol. XX, 78: 465-474. [ Links ]
5 Campos, N. 1988. Contenido de metales pesados en el chivo mapalé Cathorops spixiióe la Ciénaga Grande de Santa Marta. Resumen VI Semin. Nal. Cien. Tecnol. Mar, CCO, Bogotá: 9-16. [ Links ]
6 Cervigón, R; R. Cipriani; W. Fischer; L. Garibaldi; M. Hendrick; A. J. Lemus; R. Márquez; J. M. Poutiers; G. Robaina y B. Rodríguez. 1992. Fichas FAO de identificación de especies para los fines de la pesca. Guía de campo de las especies comerciales marinas y de aguas salobres de la costa septentrional de Sur América. Roma, FAO, 513 p. [ Links ]
7 Estrada, M. 1987. Determinación de plaguicidas organoclorados en peces (Mugil incilisy Cathorops spixii) y ostras (Crassostrea rhizophoraé) de la Ciénaga Grande de Santa Marta. Informe final Proyecto Estudio ecológico de la Ciénaga Grande de Santa Marta -1a. etapa-. COLCIENCIAS-INVEMAR, Santa Marta, 58 p. [ Links ]
8 Fischer, W. 1978. FAO Species Identification Sheets for Fishery Purposes. Western Central Atlantic (Fishing Area 31). FAO. Vol. I-V, Roma. [ Links ]
9 Galvis, O. 1983. Los áridos de la Ciénaga Grande de Santa Marta. Tesis M. Sc. Biol. Mar., Univ. Nacional de Colombia, Bogotá, 163 p. [ Links ]
10 Gayanilo, F. O; P. Sparre and D. Pauly. 1993. the FiSAT Users Guide. FAO-ICLARM Stock Assessment Tools. FAO Comput. Inf. Ser. Fish. 99, Roma. [ Links ]
11 Gulland, J. A. 1971. Fish Resources of the Ocean. Fishing New Books, London, 255 p. [ Links ]
12 Holden, M. J. y D. F. S Raitt. 1975. Manual de ciencia pesquera. Parte II: Métodos para investigar los recursos pesqueros y su aplicación. FAO. Doc. Tec. de Pesca, Roma, 115: 1-211. [ Links ]
13 IGAC. 1973. Monografía del Departamento del Magdalena. Min. Hacienda y Crédito Público. Instituto Geográfico Agustín Codazzi, Bogotá. 162 p. [ Links ]
14 Mancera, J. E. y L. Botero, 1993. Estudio ecológico de la Ciénaga Grande de Santa Marta, delta exterior del Río Magdalena, 3a. etapa. Informe de avance. Proy. COLCIENCIAS-INVEMAR. 128 p. [ Links ]
15 Mancera, J. E. 1994. Hacia un ordenamiento pesquero de la Ciénaga Grande de Santa Marta. Informe Programa Lagunas Costeras. INVEMAR, Santa Marta, 19 p. [ Links ]
16 Margalef, R. 1983. Limnología. Ed. Omega S. A., Barcelona, 1010 p. [ Links ]
17 Mendoza, J. 1993. A preliminary budget for the northeastern Venezuela shelf ecosystem. In: Trophic models of aquatic ecosystem, Manila. ICLARM -DANIDA, 1: 256-297. [ Links ]
18 Pauly, D. 1984. Fish population dynamics in tropical waters: a manual for use for programmable calculators. ICLARM stud. Rev, 8: 325 p. [ Links ]
19 Pauly, D. and J. L. Munro. 1984. Once more on the comparison of growth in fish and invertebrates, Fishbyte, 2(1): 21. [ Links ]
20 Pauly, D. 1987. A review of the ELEFAN system for analysis of length-frequency data in fish and aquatic invertebrates, p. 7-34. In: D. Pauly and G. R. Morgan (Editors), Length-based methods in Fisheries Research. ICLARM, Conf. Proc. 13, 468 p. [ Links ]
21 PRO-CIÉNAGA, 1995. Plan de manejo ambiental de la subregión Ciénaga Grande de Santa Marta 1995-1998. CORPAMAG - INVEMAR, Santa Marta. [ Links ]
22 Rueda, M. 1995. Evaluación de la eficiencia y selectividad de las redes de enmalle en la Ciénaga Grande de Santa Marta, Caribe Colombiano. Tesis Ing. Pesquera, Univ. del Magdalena. Santa Marta, 107 p. [ Links ]
23 Rueda, M. y A. Santos-Martínez. 1997. Evaluación de la eficiencia y selectividad de la red bolichera en la pesquería de la Ciénaga Grande de Santa Marta, Caribe Colombiano. Bol. Invest. Mar. Cost., 26: 17-34. [ Links ]
24 Santos-Martínez, A. 1989. Estudio biológico y ecológico de la ictiofauna de la Ciénaga Grande de Santa Marta, Caribe colombiano. Tesis M.Sc. Univ. Nacional de Colombia, Bogotá, 178 p. [ Links ]
25 Santos-Martínez, A.; J. L. Mendo; J. E. Mancera; R. Giraldo; L. Botero; E. Viloria y M. Rueda. 1995. Biomasa íctica total y su distribución espacial en la Ciénaga Grande de Santa Marta, Caribe colombiano. Resúmenes VI Congreso Latinoamericano de Ciencias del Mar COLACMAR. Mar del Plata, Argentina. 345 p. [ Links ]
26 Santos-Martínez, A. 1998. Informe final Proyecto Evaluación de los Principales Recursos Pesqueros de la Ciénaga Grande de Santa Marta, Costa Caribe Colombiana. COLCIENCIAS. [ Links ]
27 Sparre, P. y S.C. Venema. 1997. Introducción a la evaluación de recursos pesqueros tropicales. Parte 1. Manual. FAO Documento Técnico de Pesca, 306. 1(2): 420 p. [ Links ]
FECHA DE RECEPCIÓN: 27/02/0000 FECHA DE ACEPTACIÓN: 18/04/1998
