










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.30 no.1 Santa Marta Jan./Dec. 2001
POSTLARVAS Y JUVENILES DE CAMARONES FARFANTEPENAEUS SPP Y XIPHOPENAEUS KROYERI EN LA BOCA DE LA BARRA (CIÉNAGA GRANDE DE SANTA MARTA), CARIBE COLOMBIANO
POSTLARVAE AND JUVENILES OF THE SHRIMPS FARTANTEPENAEUS SPP. AND XIPHOPENAEUS KROYERY AT BOCA DE LA BARRA (CIENAGA GRANDE DE SANTA MARTA), COLOMBIAN CARIBBEAN
Ángela C. López 1, María M. Criales 2 y Camilo B. García3
1 Instituto de Investigaciones Marinas y Costeras - INVEMAR, AA 1016, Santa Marta, Colombia. E-mail: alopez@invemar.org.co (ACL).
2 RosenstielSchool and Atmosperic of Marine Science -RSMAS- (Universidad de Miami), 4600 Rickenbacker Causeway Miami, Fl 33149,EE.UU. E-mail: mcriales@rsmas.miami.edu (MMC).
3 Universidad Nacional de Colombia (Departamento de Biología)/INVEMAR, AA 1016, Santa Marta, Colombia. E-mail: cgarcia@invemar.org.co (CBG).
RESUMEN
Postlarvas y juveniles de los camarones Farfantepenaeus spp. y Xiphopenaeus kroyeri se colectaron en la Boca de la Barra (Ciénaga Grande de Santa Marta), mediante redes de canal fijas que se orientan con la marea. Se realizaron muestreos quincenales nocturnos durante un año (febrero 1998 - febrero 1999), correspondiendo a los períodos de luna nueva y luna llena, intercaladamente. Para Farfantepenaeus spp. se observó que las postlarvas tienden a presentarse durante la época seca del año y preferiblemente en la fase de luna llena, siendo la diferencia entre fases lunares estadísticamente significativa (test de Mann-Whitney, P<0.05). Los juveniles presentaron un comportamiento opuesto: hubo diferencias significativas (test de Mann-Whitney, P< 0.05) entre épocas climáticas, mostrándose en mayor concentración en la época húmeda, pero no entre las fases lunares (test de Mann-Whitney, P>0.05). Para X. kroyeri en ningún caso las diferencias entre épocas climáticas o fases lunares fueron significativas (test de Mann-Whitney, P>0.05). Sin embargo, se pueden señalar tendencias: las postlarvas se presentaron en mayor proporción durante la luna nueva, pero sin tendencias respecto a la época climática mientras que en los juveniles las diferencias se observan por época climática (mayor proporción en la época húmeda), y no entre las fases lunares. Se discuten estos resultados en relación con los escasos antecedentes históricos. Se registran las relaciones LT-LC y LT-Peso y algunas características merísticas y morfométricas.
PALABRAS CLAVE: Farfantepenaeus spp. Xiphopenaeus kroyeri. Postlarvas. Juveniles. Distribución temporal.
ABSTRACT
Postlarvae and juveniles of Farfantepenaeus spp. and Xiphopenaeus kroyeri were collected at Boca de la Barra (Ciénaga Grande de Santa Marta) by means of a channel net that orientates with the tide. Sampling was carried out every two weeks for a year (February 1998-February 1999) so that each sampling date corresponded to full and new moon alternatively. Farfantepenaeus spp. postlarvae tended to appear in the dry season and with significantly greater abundance during full moon (Mann-Whitney test, P<0.05). Juveniles showed a different pattern: there were significant differences between seasons (Mann-Whitney test, P<0.05) with higher concentrations in the rainy season, but not so between moon phases (Mann-Whitney test, P>0.05). In the case of X. kroyeri in no instance significant were differences found between seasons or moon phases (Mann-Whitney test, P>0.05). However, some tendencies can be pointed out: postlarvae appeared in higher proportions during the new moon but no major difference was recorded between seasons whereas juveniles appeared in greater concentracions in the rainy season with almost no difference between moon phases. These results are disscused in terms of the scanty historical previous work. The relations TL-CL and CL-Weight and some morphometric and meristic features are reported.
KEY WORDS: Farfantepenaeus spp. Xiphopenaeus kroyeri. Postlarvae. Juveniles. Temporal distribution.
INTRODUCCIÓN
Los camarones Farfantepenaeus spp. y Xiphopenaeus kroyeri se cuentan entre los principales recursos que sustentan las pesquerías artesanales de la Ciénaga Grande de Santa Marta (CGSM) (INVEMAR, 1998). Estos crustáceos, desovan mar afuera, en donde permanecen durante sus primeras etapas de desarrollo (nauplio, mysis, zoeas y los primeros estadios de postlarvas (García y Le Reste, 1987). Una vez se convierten en postlarvas, migran hacia los estuarios, como la CGSM, en busca de alimento y protección (Allen et al., 1980; García y Le Reste, 1987). En el estuario las postlarvas culminan su desarrollo convirtiéndose en juveniles con un número definitivo de espinas en el rostrum y diferenciación de sus estructuras reproductivas aun cuando no madurez sexual, características que los asemejan al estado adulto (Chapa, 1975; García y Le Reste, 1987).
Conocimientos acerca de la biología y ciclos de vida de especies comerciales proveen información útil para comprender la dinámica poblacional y reproductiva de las especies y el manejo apropiado de las mismas. Por esto se hace necesario la realización de trabajos encaminados a describir estos ciclos y el comportamiento de los individuos en sus diferentes etapas, ya que esta información puede facilitar el entendimiento de las variaciones estaciónales de una población y, en el caso de los estados larvales y juveniles, aportar datos acerca de aspectos reproductivos de las especies.
En la CGSM trabajos en los que se incluyen estaciones en la Boca de la Barra (BB), (comunicación de la CGSM con el mar, sitio del presente estudio, ver abajo) han sido realizados por Buenaventura y Arango (1980) y Perea (1981) y en ellos se describen patrones migratorios, variaciones temporales en abundancia, biomasa y características merísticas y morfométricas de postlarvas y juveniles de camarones incluidos en el género Farfantepenaeus. Sánchez (1991) realizó estudios de este tipo en el área, pero sólo a nivel de juveniles. Información sobre Xiphopenaeus kroyeri se presenta en los trabajos de Cortés (1989) Cortés y Críales (1990) y Cortés y Newmark (1992), quienes trabajaron con juveniles y adultos de las especie, en el sector de Costa Verde, un área marina cercana a la BB.
En el presente estudio se determinaron la densidad y biomasa de postlarvas y juveniles de camarones Farfantepenaeus spp. y X. kroyeri en una zona de tránsito como es la BB, con el fin de establecer los cambios de estas variables respecto a épocas climáticas y fases lunares y de esta forma aportar al conocimiento de la dinámica en el tiempo de estas especies en un sitio clave para sus migraciones. Información sobre características merísticas como el número de espinas dorsales en el rostrum y morfométricas como la longitud total y longitud caparazón, también es incluida en este estudio, debido a su importancia en la diferenciación entre especies y estadios. Así mismo, la relación entre los valores de longitud total y longitud caparazón registrada en este trabajo podrá utilizarse en futuros estudios para calcular el valor de una de las dos variables sin necesidad de determinar la otra.
Cabe señalar que el registro de camarones en este estudio bajo el nombre de Farfantepenaeus spp. se realizó teniendo en cuenta el trabajo de Pérez-Farfante y Kensley (1997) y corresponde al género denominado como Penaeus spp. en trabajos anteriores. Por otra parte, en los trabajos de Buenaventura y Arango (1980), Perea (1981), Sánchez (1991) y Santos-Martínez (1998), las especies referidas como Penaeus aztecus subtilis y Penaeus duorarum notialis corresponden a Farfantepenaeus subtilis y Farfantepenaeus notialis, respectivamente (Pérez-Farfante, 1988).
AREA DE ESTUDIO
La CGSM se encuentra localizada entre los 10o40' - 10o59' N y 74o 26' - 74o 38' W, con un área de 450 km² (Figura 1). Esta Ciénaga se constituye como la más importante laguna costera de Colombia desde el punto de vista ecológico y socioeconómico, debido a que soporta una de las principales pesquerías artesanales en el país.
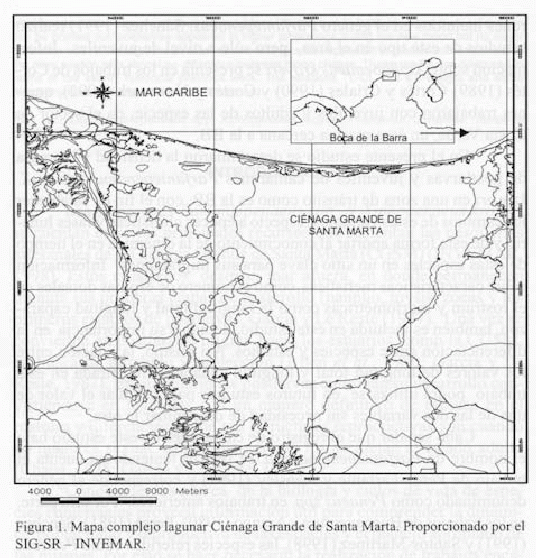
El área se caracteriza por presentar un clima seco a semihúmedo (IGAC, 1978) con dos épocas climáticas definidas: Una seca (diciembre a abril), donde predominan los vientos alisios del noreste y una húmeda (septiembre a noviembre) donde se presentan vientos de poca fuerza y dirección variable, con un notorio aumento de las precipitaciones locales y aportes de los ríos que desembocan en la CGSM (Blanco, 1988; CORPES, 1992). Sin embargo, eventos climáticos como "El Niño" pueden ocasionar variaciones en la temporalidad de estas épocas.
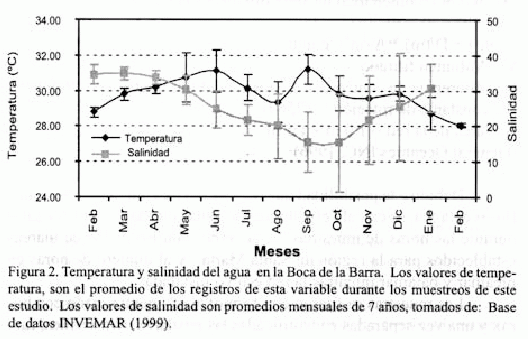
El punto de muestreo en este estudio se ubico en la BB, la cual presenta un ancho aproximado de 250 m (Botero, 1988) y una profundidad que oscila entre 2 y 8 m (Base de datos INVEMAR, 2000). En la figura 2, se representan valores mensuales de salinidad (Base de datos INVEMAR, 2000) y temperatura del agua (presente estudio) en la BB. Se puede observar que la primera oscila entre 15.4 y 34.7, presentándose los mayores valores en febrero, marzo y abril (época seca) y los menores en septiembre y octubre (época húmeda). La temperatura del agua se encuentra entre 28.06 oC y 31.19 oC y al contrario de la salinidad la tendencia es a presentarse los mayores valores durante la época húmeda y los menores en la seca.
La BB se encuentra sometida a la influencia directa de las mareas diarias, las cuales son irregulares con rango micromareal semidiurno y una amplitud que oscila entre 20 y 30 cm. La marea diurna tiene una amplitud de 15 cm con una altura máxima de 11.35 cm y mínima de 3.62 cm (Martínez et al., 1992). En esta área se destacan dos tipos de circulación del agua: en época seca el sentido del flujo de agua predominante es hacia adentro de la CGSM, presentándose movimientos giratorios hacia la izquierda desde la BB; en época húmeda, cuando son importantes los aportes de los ríos que desembocan en la CGSM, los
flujos de agua son preferencialmente hacia fuera (INDERENA, 1972; CORPAMAG, 1995). El aporte promedio de agua de la Ciénaga al mar a través de la Boca de la Barra, es de 560 m3/s (com. pers. B.F. Jönsson).
MATERIALES Y MÉTODOS
Entre febrero 1998 y febrero 1999 se colectaron postlarvas y juveniles de camarones penéidos del género Farfantepenaeus y del camarón titi X. kroyeri en la BB, mediante muestreos quincenales nocturnos durante las fases de luna nueva y luna llena (dos noches por fase). Se empleó para ello una red de canal (1.0 m x 1.5 m de boca y 5.5 m de largo) fija a un bloque de concreto, cuya boca se extiende a 1 m de profundidad. La red se mueve libremente con los cambios de marea, permitiendo capturar de esta manera tanto las postlarvas que entran a la Ciénaga en busca de protección y alimento como los juveniles que salen para culminar su desarrollo. El copo de la red se colocaba a las 19:00 horas y se retiraba a las 7:00 horas de la mañana siguiente.
La boca de la red se equipó con un flujómetro General Oceanics, el cual permitió calcular el volumen total de agua filtrado por la red, durante las 12 horas de muestreo cada noche. La formula utilizada para calcular el volumen total de agua filtrado en la red es:
V (m3) = D (m) * A (m2), donde,
V = Volumen filtrado
D = diferencia de conteos final e inicial del flujómetro * R / 999999
(R: constante de rotación = 57560)
A = área de la red = 1.0 m x 1.5 m
(General Oceanics INC, 1996)
Debido a la posibilidad que tiene la red de girar de acuerdo a los flujos de agua, se calculó el volumen de agua filtrada que entró y salió durante las horas de muestreo de acuerdo con los ciclos de mareas establecidos para la región de Santa Marta, y al número de horas en pleamar y bajamar mientras la red estuvo instalada.
Las muestras se fijaron con formol al 4% neutralizado con bórax y una vez separadas e identificadas las postlarvas y juveniles, fueron preservadas en alcohol etílico al 70%. Conociendo los volúmenes filtrados de entrada y salida y teniendo en cuenta que según el ciclo de vida de Farfantepenaeus spp. y X. kroyeri, las postlarvas entran al estuario y los juveniles salen (García y Le Reste, 1987), se calcularon las densidades de los estadios para ambas taxa y se expresaron finalmente como indv./1000m3. La biomasa de cada uno de los individuos colectados fue medida en una balanza analítica Sartorius y se expresó como peso húmedo (g/1000m3).
Adicionalmente, se determinó la longitud total (LT), longitud caparazón (LC) y el número de espinas dorsales del rostrum, consideradas características importantes para la diferenciación entre géneros y estadios larvales. En la tabla 1 se presentan las principales características a partir de las cuales se diferencian los géneros de camarones penéidos aquí estudiados y en cada uno de ellos entre postlarvas y juveniles. Se establecieron relaciones LT - biomasa y LT - LC, a partir de los valores obtenidos para los individuos de cada taxa.
La variación temporal de las densidades y biomasas de postlarvas y juveniles de ambas taxa (por épocas climáticas y fases lunares), se representó gráficamente. Se aplicó la prueba no paramétrica de Mann-Whitney, para probar si existen diferencias significativas en densidad o biomasa entre épocas climáticas y entre fases lunares (Zar, 1984).
Las épocas climáticas se determinaron teniendo en cuenta los valores totales de precipitación mensual durante el periodo de muestreo

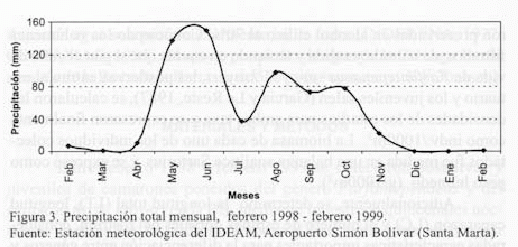
(datos tomados de: Estación Meteorológica IDEAM, Aeropuerto Simón Bolívar Santa Marta). Estos valores se representan en la figura 3, a partir de la cual se puede decir que la época seca corresponde a los meses de diciembre a abril y la época húmeda de mayo a noviembre, en el año de muestreo.
RESULTADOS
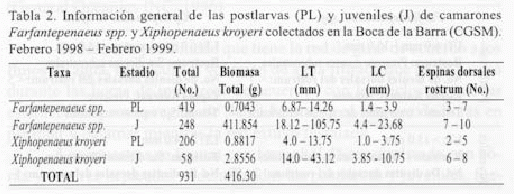
En la tabla 2 se presentan las estadísticas globales del estudio. Como se observa, de los 931 individuos capturados en total, la mayor parte corresponden a postlarvas y juveniles de Farfantepenaeus spp. Respecto a la biomasa, son los juveniles de Farfantepenaeus spp. los que más aportan a este valor. Además de las abundancias totales y biomasa, en la tabla 2 se registran los rangos de tallas (LT, LC) para ambas taxa y el rango de número de espinas dorsales del rostrum.
Distribución Temporal
En las figuras 4a y 4b se compara el comportamiento mensual de la densidad y biomasa de postlarvas y juveniles de Farfantepenaeus spp. Se encontró una concentración diferencial de postlarvas y juveniles, tendiendo los primeros a presentar mayores concentraciones en enero/99 (época seca), mientras que los juveniles aparecen en mayores concentraciones en noviembre/98 (época húmeda). Otro incremento de menor proporción en la densidad y biomasa de postlarvas, se observó en mayo/98 (época húmeda).
Al relacionar las épocas climáticas y las fases lunares para Farfantepenaeus spp. se encontró que para las postlarvas existe una asociación entre época seca y luna llena, expresada por valores más altos y mayor frecuencia de aparición en esta combinación (Figura 5a).
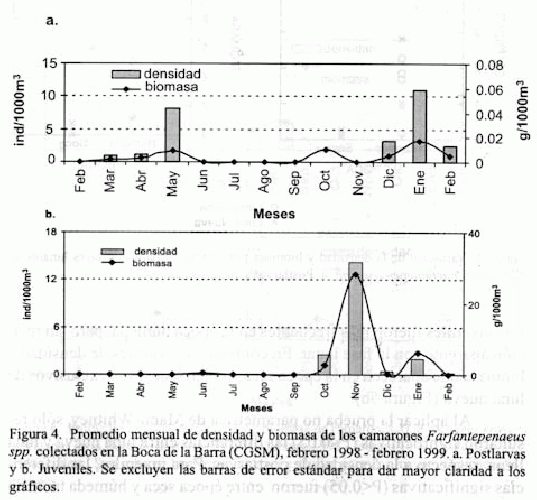
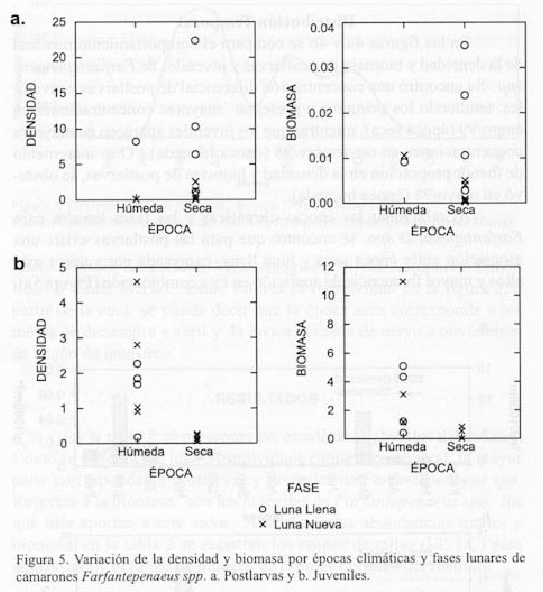
Los juveniles fueron más frecuentes en la época húmeda, pero sin relación aparente con la fase lunar. En contraste, los valores de densidad y biomasa que aparecen en la época seca, si bien bajos, son exclusivos de luna nueva (Figura 5b).
Al aplicar la prueba no paramétrica de Mann-Whitney, sólo resultaron significativas (P<0.05) las diferencias entre luna nueva y luna llena respecto a la densidad de postlarvas. Para juveniles, las diferencias significativas (P<0.05) fueron entre época seca y húmeda tanto en densidad como en biomasa.
Para X. kroyeri, en la figura 6a se observa que las densidades y biomasas de las postlarvas se dan en los primeros meses de 1998, correspondiendo el mayor valor a marzo y un segundo notoriamente diferente al primero en mayo. A diferencia del comportamiento presentado por Farfantepenaeus spp. no es tan clara la diferencia en los meses de aparición de las postlarvas y juveniles (Figura 6b), pues estos últimos aunque presentaron su valor mas alto en agosto/98, también se observaron en cantidades apreciables durante mayo/98.
La figura 7a, que relaciona épocas climáticas y fases lunares para postlarvas de X. kroyeri, sugiere, que hay una mayor incidencia de estas en luna nueva, independientemente de la época climática. Los juveniles parecen presentar un comportamiento inverso, con mayor
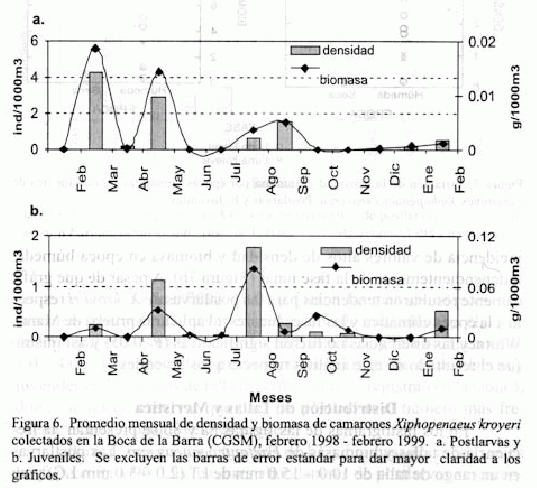
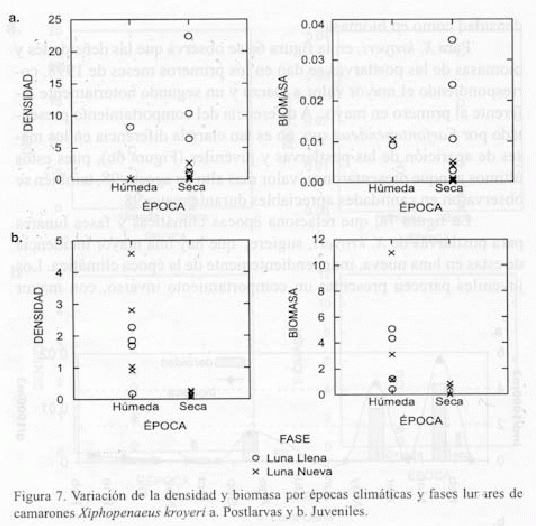
incidencia de valores altos de densidad y biomasa en época húmeda, independientemente de la fase lunar (Figura 7b). A pesar de que gráficamente resultaron tendencias para las postlarvas de X. kroyeri respecto a la época climática y las fases lunares, al aplicar la prueba de Mann-Whitney las diferencias no fueron significativas (P>0.05) y así mismo fue el resultado de este análisis respecto a los juveniles.
Distribución de Tallas y Merística
En los histogramas de las figuras 8a y 8b se presentan la frecuencia de tallas y biomasas de Farfantepenaeus spp. Las postlarvas en un rango de talla de 10.0 - 15.0 mm de LT (2.0 - 3.0 mm LC) y un
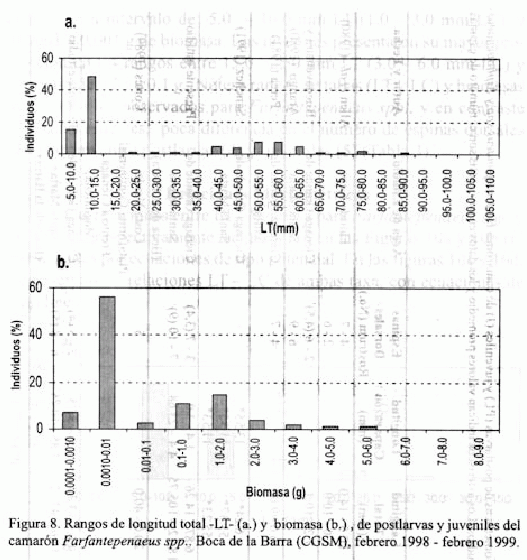
rango de biomasa de 0.0010 - 0.01 g fueron las más abundantes. Los juveniles mostraron la mayor concentración de individuos en el rango de LT entre 40.0 - 65.0 mm (9.0 - 15.0 mm LC) y una biomasa entre 1.0 - 4.0 g. El número de espinas dorsales del rostrum de postlarvas y juveniles en los rangos de talla descritos aquí, se registran en la tabla 3, donde se puede observar que para las postlarvas el número más frecuente es tres y para juveniles nueve.
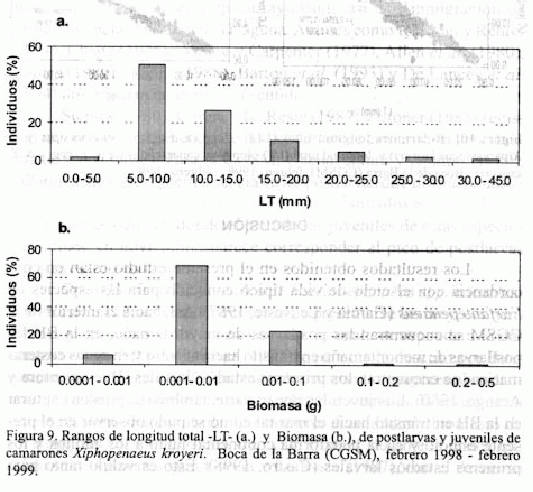
Para X. kroyeri los intervalos de LT y biomasa se muestran en las figuras 9a y 9b. Las mayores abundancias de postlarvas se encontraron
en el intervalo de 5.0 - 10.0 mm LT (1.0 - 3.0 mm LC), y 0.0001 - 0.001 g de biomasa. Los juveniles presentaron su mayor proporción en los rangos entre 15.0 -25.0 mm LT (3.0 - 6.0 mm LC) y biomasa de 0.01 - 0.1 g. Nótese rangos de tallas (LT y LC) y biomasas menores a los observados para Farfantepenaeus spp., y en contraste con estas especies, poca diferencia en el número de espinas dorsales del rostrum entre postlarvas (4) y juveniles (5) (Tabla 3).
Relaciones Morfométricas
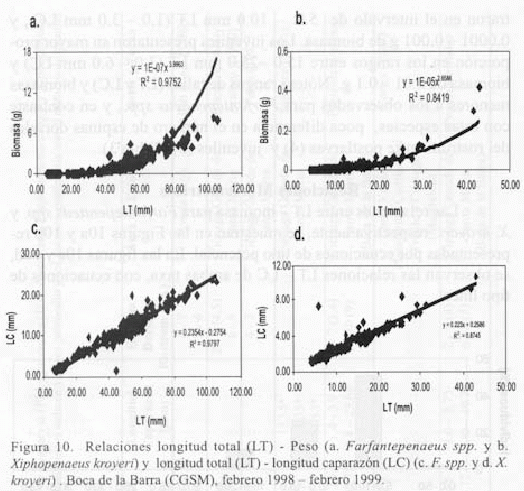
Las relaciones entre LT - biomasa para Farfantepenaeus spp. y X. kroyeri respectivamente, se muestran en las Figuras 10a y 10b representadas por ecuaciones de tipo potencial. En las figuras 10c y 10d, se observan las relaciones LT - LC de ambas taxa, con ecuaciones de tipo lineal.
DISCUSIÓN
Los resultados obtenidos en el presente estudio están en concordancia con el ciclo de vida típico conocido para las especies de Farfantepenaeus (García y Le Reste, 1987). Así, hacia el interior de la CGSM se encuentran las postlarvas de mayor tamaño, en la BB las postlarvas de menor tamaño en transito hacia adentro y en aguas costeras marinas se encuentran los primeros estadios larvales (Buenaventura y Arango, 1980). Los juveniles por su parte, también se pueden capturar en la BB en tránsito hacía el mar tal como se pudo observar en el presente estudio y en la plataforma continental junto a los adultos y los primeros estadios larvales (Castro, 1998). Esto es valido tanto para Farfantepenaeus spp. como para X. kroyeri (Buenaventura y Arango, 1980; Perea, 1981; Cortés y Newmark 1992; Sánchez, 1991).
Con relación a la estacionalidad en la aparición de postlarvas de Farfantepenaeus spp., la tendencia observada aquí (mayores densidades y biomasas durante la época seca del año) confirma los trabajos realizados por Buenaventura y Arango (1980) y Perea (1981), quienes encontraron las mayores abundancias de postlarvas en diciembre, enero, febrero y marzo, meses considerados como secos. Un mecanismo plausible para explicar esta observación es la relación entre la acción de las mareas y la descarga neta de los cuerpos de agua como la CGSM al mar. Así, el predominio de postlarvas de Farfantepenaeus spp. durante la época seca puede deberse a que la resistencia al ingreso a la CGSM por la BB es mucho menor, por ser menor la descarga de agua en comparación con la época de lluvias y a que los vientos del NE son mas intensos durante esta época, ayudando así a la inmigración de postlarvas hacia el interior de la laguna. Autores como Roessler y Rehrer (1971), Chapa (1975), Young y Carpenter (1977), Allen et al. (1980), Staples (1980), Stoner (1988), Barton et al. (1993) y De Lancey et al. (1994) argumentan en el mismo sentido.
Staples (1980), García y Le Reste (1987) y Stoner (1988) registran tiempos de permanencia de las postlarvas de camarón en los estuarios de 2 - 6 meses. Para la CGSM, Buenaventura y Arango (1980), hallaron un tiempo de permanencia de las postlarvas de Farfantepenaeus spp. de 4 - 6 meses, lo cual coincide con los resultados encontrados en el presente estudio, en donde el pico de los juveniles de estas especies encontrado en noviembre, parece corresponder al pico de postlarvas que ingresaron en mayo.
Con relación a los juveniles de Farfantepenaeus spp. encontrados en este estudio, la tendencia fue la de presentarse las mayores densidades y biomasas en la época húmeda, lo cual corresponde a lo encontrado en la BB por Sánchez (1991), pero no a lo descrito por Buenaventura y Arango (1980), quienes observaron las mayores abundancias de juveniles durante meses considerados como de la época seca (enero, febrero, marzo, abril). Las mayores densidades y biomasas de juveniles de Farfantepenaeus spp. y X. kroyeri, durante la época húmeda se relacionan según García y Le Reste (1987), con el descenso pronunciado de la salinidad por efecto de las lluvias, que hace que los juveniles migren hacia el mar en busca de salinidades mayores que favorezcan su desarrollo a adulto y posterior desove.
Con relación a las fases lunares, para las postlarvas de Farfantepenaeus spp. no se encontró similitud con lo registrado por Buenaventura y Arango (1980) en la BB, quienes no hallaron diferencias en las abundancias de postlarvas entre fases lunares. Tampoco se presentó similitud con lo hallado por Roessler y Rehrer (1971), Chapa (1975) y Allen et al. (1980), quienes registraron las mayores densidades de postlarvas de Farfantepenaeus spp. durante luna nueva, siendo esto último atribuido por estos autores a que las postlarvas aprovechan los periodos de oscuridad para transportarse protegiéndose de la predación. Si bien García y Le Reste (1987) confirman que el patrón general de las postlarvas de camarones penéidos es el de presentarse durante luna nueva, también señalan la existencia de estudios donde los mayores picos se presentan en fases diferentes a esta, tal como ocurrió en el presente estudio (mayores densidades en luna llena) y el presente es uno de esos casos. Las postlarvas de X. kroyeri presentaron mayores densidades y biomasas durante luna nueva, lo cual está de acuerdo con el patrón general descrito por García y Le Reste (1987) para las postlarvas de camarones peneidos.
El comportamiento respecto a las fases lunares mostrado por los juveniles de Farfantepenaeus spp. y X. kroyeri colectados en el presente estudio, no corresponde al patrón general señalado por Chapa (1975) y García y Le Reste (1987), quienes sugieren que juveniles y preadultos de camarones penéidos responden a estímulos luminosos y por tanto es de esperarse su preferencia en los períodos de luna llena. En el presente estudio los juveniles de Farfantepenaeus spp. se presentaron en mayores densidades durante el período de luna nueva y los juveniles de X. kroyeri, no mostraron diferencias significativas entre fases lunares. Con relación a los trabajos realizados anteriormente en el área tampoco se encuentra relación con lo descrito aquí; Sánchez (1991) no encontró diferencias significativas entre fases lunares para Farfantepenaeus spp. y Cortés y Newmark (1992) observaron que los juveniles de X. kroyeri son más abundantes en luna llena.
Una hipótesis que podría explicar el comportamiento migratorio tanto de postlarvas como juveniles en la CGSM, esta relacionada con la conexión existente entre las mareas, los ciclos lunares y el régimen hidrológico en el área. De esta manera, una mayor entrada de postlarvas de Farfantepenaeus spp., coincide con la fase de luna llena y la época seca, cuando las descargas de agua de la ciénaga al mar son menores y se puede notar con mayor claridad el efecto de las mareas. Caso contrario se observa para los juveniles, que se presentan con mayor abundancia durante la época húmeda cuando las descargas de agua de la ciénaga al mar son mayores por efecto de las lluvias, haciendo que la acción de las mareas sea menos notoria y coincidiendo también, con mayores abundancias en la fase de luna nueva.
Sin embargo, comparando los escasos datos históricos con los del presente estudio, no es evidente un patrón definido para el área respecto al comportamiento de larvas y juveniles con relación a las fases lunares.
En la tabla 3 donde se describen los rangos de tallas para postlarvas y juveniles de Farfantepenaeus spp. y X. kroyeri , se observa que el rango obtenido en este estudio está incluido en el rango de tallas (LT) descrito por Allen et al. (1980) para tres especies de Farfantepenaeus (F. aztecus, F. brasiliensis y F. duorarum) en el área dede Florida Keys. En la CGSM, las especies de Farfantepenaeus son F. subtilis y F. notialis (INVEMAR, 1998) y es probable que a estas especies pertenezcan las postlarvas y juveniles encontradas aquí.
Comparando resultados del presente trabajo con los de Buenaventura y Arango (1980) y Perea (1981), se encontraron diferencias respecto a los rangos de LT descritos aquí para Farfantepenaeus spp.; estos autores registran rangos menores a los del presente estudio. Estas diferencias pueden estar relacionadas con el tipo de red y muestreo utilizados. Tanto Buenaventura y Arango (1980) como Perea (1981) realizaron arrastres cubriendo solo los 50 cm superiores de la columna de agua, lo cual pudo impedir que se colectaran postlarvas de mayor tamaño y mayor número de espinas dorsales en el rostrum, que están en proceso de asentamiento. En el presente estudio, la red utilizada se extendió 1 m en la columna de agua lo cual posiblemente permitió capturar postlarvas en proceso de asentamiento y por tanto de mayor tamaño.
AGRADECIMIENTOS
La presente investigación se realizó gracias al apoyo logístico y financiero del Instituto de Investigaciones Marinas y Costeras (INVEMAR) y está enmarcada dentro del proyecto "Patrones de Reclutamiento y Mecanismos de Transporte Costero de Estadios Tempranos de Desarrollo de Peces y Crustáceos Decápodos en el Área de Santa Marta, Caribe Colombiano" (Código 2105-09-689-96), financiado por el Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología (COLCIENCIAS). El proyecto se ejecutó bajo el convenio INVEMAR - Universidad de Miami.
BIBLIOGRAFIA
1 Allen, D. M., J. H. Hudson y T. J. Costello. 1980. Postlarval shirmp (Penaeus) in the Florida Keys: species, size and seasonal abundance. Bull. Mar. Sci., 30 (4): 21-33. [ Links ]
2 Barton, D. R., R. F. Shaw, W. H. Herke y R. H. Blanchet. 1993. Recruitment of postlarval and juvenile brown shrimp (Penaeus aztecus Ives) from offshore to estuarine waters of the northwestern Gulf of Mexico. Est. Coast. Shelf Sci., 36, 377-394. [ Links ]
3 Blanco, J. 1988. Las variaciones ambientales estacionales en las aguas costeras y su importancia para la pesca en la región de Santa Marta, Caribe colombiano. Tesis M.Sc. Biol. Mar., Universidad Nacional de Colombia. Bogotá, 50 p. [ Links ]
4 Botero, L. 1988. Estudio ecológico integrado de la Ciénaga Grande de Santa Marta. Informe Proyecto, INVEMAR. Bogotá, 200 p. [ Links ]
5 Buenaventura, D. y S. Arango. 1980. Biología y dinámica de las poblaciones de los camarones del género Penaeus en el área de Santa Marta y Ciénaga Grande. Tesis Biol., Universidad del Valle. Cali, 150 p. [ Links ]
6 Castro, P. P. 1998. Contribución al conocimiento de las larvas de crustáceos decápodos del Golfo de Salamanca, Caribe colombiano. Tesis Biol. Mar. Universidad de Bogotá Jorge Tadeo Lozano. Bogotá, 45 p + anexos. [ Links ]
7 Chapa, H. 1975. Notas sobre el comportamiento de los camarones del género Penaeus. En: Mem. II Simp. Latinoamer. Ocean. Biol., Universidad de Oriente, Cumaná, Venezuela. Tomo I: 121-138. [ Links ]
8 Cook, H. L. 1964. A generic key to the protozoean, mysis, and postlarval stages of the littoral Penaeidae of the northwestern Gulf of México. U.S. Fish Wildl. Serv. Fish. Bull., 65: 437-447. [ Links ]
9 CORPAMAG. 1995. Campaña de mediciones hidrosedimentológicas y ambientales en la Ciénaga Grande de Santa Marta. Informe Técnico, Santa Marta, 47 p. [ Links ]
10 CORPES. 1992. El Caribe colombiano: realidad ambiental y desarrollo. Consejo regional de planificación de la costa Atlántica, Santafé de Bogotá, 350 p. [ Links ]
11 Cortés, M. L. 1989. Estudio de la biología y ecología del camarón tití Xiphopenaeus kroyeri (Heller), en Costa Verde, Ciénaga, Magdalena (Caribe colombiano), con notas sobre el camarón blanco Penaeus (Litopenaeus) schmitti Burkenroad. Tesis Biol., Pontificia Universidad Javeriana. Bogotá, 117 p. [ Links ]
12 Cortes, M.L. y M.M. Criales. 1990. Analysis of the stomach contents of the shrimp Xiphopenaeus kroyeri (Heller) (Crustacea: Decapoda: Penaeidae). An. Inst. Inv. Mar. Punta Betin, 19-20: 23-33 [ Links ]
13 Cortés, M. L. y R.F. Newmark. 1992. Distribución y abundancia del camarón tití Xiphopenaeus kroyeri en Costa Verde (Ciénaga), Caribe colombiano. Bol. Ecotrópica, 25: 15 - 27. [ Links ]
14 DeLancey, L. B., J. E. Jenkins and J. D. Whitaker. 1994. Results of long-term, seasonal sampling for Penaeus postlarvae at breach inlet, South Carolina. Fish. Bull., 92: 633-640. [ Links ]
15 García, S. y L. Le Reste. 1987. Ciclos vitales, dinámica, explotación y ordenación de las poblaciones de camarones penéidos costeros. FAO Doc. Tec. Pesca, 203: 180 p. [ Links ]
17 General Oceanics INC. 1996. General Oceanics digital flowmeter mechanical & electronic operators manual. Miami. [ Links ]
18 IGAC. 1978. Atlas Regional del Caribe. Subdirección de Investigación y Divulgación Geográfica. IGAC. Bogotá, 119 p. [ Links ]
19 INDERENA. 1972. Régimen de las aguas y la salinidad en la Ciénaga Grande de Santa Marta. Características generales de la circulación. Estudio Cualitativo. Laboratorio de ensayos de hidráulica. Bogotá, 59 p. [ Links ]
20 INVEMAR, 1998. Informe Final Proyecto Evaluación de los Principales Recursos Pesqueros de la Ciénaga Grande de Santa Marta, Costa Caribe colombiana. INVEMAR-COLCIENCIAS-GTZ-PROCIÉNAGA. Santa Marta, 10 p. + anexos. [ Links ]
21 INVEMAR. 2000. Base de datos de variables fisicoquímicas Ciénaga Grande de Santa Marta. Sin publicar. Instituto de Investigaciones Marinas y Costeras - INVEMAR -, Santa Marta. [ Links ]
22 Martínez, J.O., J. L. González, J. V. Franco, L. Marín y L. H. Molina y F. Pérez. 1992. Geomorfología y aspectos erosivos del litoral Caribe colombiano. Mem. VIII Sem. Nal. Cienc. Tecnol. Mar., CCO. Santa Marta, 1: 56-70. [ Links ]
23 Perea, F. L. 1981. Contribución al conocimiento de la ecología de dos especies de camarones penéidos, Penaeus schmitti (Burkenroad) y Penaeus aztecus subtilis (Pérez Farfante) en la región de Santa Marta (Magdalena). Tesis Biol., Universidad del Valle. Cali, 45 p. [ Links ]
24 Pérez-Farfante, I. 1970. Características diagnósticas de los juveniles de Penaeus aztecus subtilis, P. duorarum notialis y P. brasiliensis (Crustácea Decápoda Penaeidae). Mem. Soc. Cienc. Nat. La Salle, 30(87): 182p. [ Links ]
25 ___________. 1988. Illustrated key to penaeoid shrimps of commerce in the Americas. NOAA Technical Report NMFS, 64: 32p. [ Links ]
26 ___________ y B. Kensley. 1997. Penaoid and sergestoid shrimps and prawns of the world keys and diagnoses for the families and genera. Du Museums París. Washington. 233p. [ Links ]
27 Reyes, H. 1975. Posición taxonómica y descripciones de las postlarvas de Penaeus spp de la Bahía de Yavros y estuario de río Mayo Sonora, México. Mem. II Simp. Latinoamer. Ocean. Biol. Universidad de Oriente. Cumaná, Venezuela. Tomo I: 9 -18. [ Links ]
28 Roessler, M. A. y R. G. Rehrer. 1971. Relation of catches of postlarval pink shrimp in Everglades National Park, Florida, to the commercial catches on the Tortugas grounds. Bull. Mar. Sci., 21 (4): 790 - 805. [ Links ]
29 Sánchez, L.A. 1991. La Ciénaga Grande de Santa Marta como área de cría para diferentes especies de camarones juveniles del género Penaeus. Tesis Biol., Universidad Nacional de Colombia. Bogotá, 63p. [ Links ]
30 Staples, D.J. 1980. Ecology of juvenile and adolescent banana prawns, Penaeus merguiensis, in mangrove estuary and adjacent off-shore area of the Gulf of Carpentaria. II. Emigration, populatión structure and growth of juveniles. Aus. J. Mar. Freshwater Res., 31: 653-665. [ Links ]
31 Stoner, A. W. 1988. A nursery ground for four tropical Penaeus species: Laguna Joyuda, Puerto Rico. Mar. Ecol., 42: 133 - 141. [ Links ]
32 Young, P. C. and S. M. Carpenter. 1977. Recuitment of postlarval penaeid prawns to nursery areas in Moreton Bay, Queensland. Aus. J. Mar. Freshwater Res., 28: 745-773. [ Links ]
33 Zar, J. H. 1984. Biostatistical análisis. Prentice-Hall, New Jersey, 716 p. [ Links ]
FECHA DE RECEPCION: 08/11/1999 FECHA DE ACEPTACION: 12/09/2001
