










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.33 no.1 Santa Marta Jan./Dec. 2004
ANÁLISIS DEL CRECIMIENTO Y LA MORTALIDAD DE ANISOTREMUS INTERRUPTUS (GILL) (PERCIFORMES: HAEMULIDAE) EN LA COSTA DE COLIMA, MÉXICO
Growth and mortality of Anisotremus interruptus (Gill) (Perciformes: Haemulidae) on the coast of Colima, México.
Elaine Espino-Barr, Manuel Gallardo-Cabello, Fernando González-Orozco y Arturo Garcia-Boa
RESUMEN
En el presente trabajo se realiza un análisis sobre el crecimiento y la mortalidad total y natural del bacoco Anisotremus interruptus en las costas de Colima, México. Los parámetros de crecimiento estimado son los siguientes: L∞= 50.59 cm; W∞= 5051.04 g; k = 0.147 año-1; t0 = -0.916 años; A0.95 = 19.46 años. El periodo de mayor crecimiento lineal ocurre durante el primer año de vida, cuando el bacoco crece 12.52 cm, el segundo año crece 4.95 cm y el tercero, 4.60 cm. Los mayores valores del factor de condición se presentan durante febrero y septiembre. El coeficiente de mortalidad total (Z) es de 0.53 año-1. Estos valores forman parte de la información necesaria para sentar las bases para la administración de la pesquería de esta especie.
PALABRAS CLAVE: Anisotremus interruptus, Haemulidae, Crecimiento, Mortalidad
ABSTRACT
This paper deals with the growth and mortality analysis of the burrito grunt Anisotremus interruptus on the coast of Colima, México. The estimated growth parameters are: L∞ = 50.59 cm; W∞ = 5,051.04 g; k = 0.147 years-1; to = -0.916 years; A0.95 = 19.46 years. Most of the growth occurred during the first year of life, when the grunt grows 12.52 cm, the second year it grows 4.95 cm and the third, 4.60 cm. The highest value of the condition index took place between February and September. The total mortality rate (Z) was calculated as 0.53 years-1. These values are basic for the plan of administration of the fishery of this species.
KEY WORDS: Anisotremus interruptus, Haemulidae, Growth, Mortality.
INTRODUCCIÓN
El bacoco Anisotremus interruptus (Gill, 1862) es una especie de importancia comercial de la pesca ribereña del estado de Colima, México, habiendo ocupado algunos años (1984 a 1990) entre el segundo y quinto lugar, por su volumen de captura, dentro de su familia (Cruz-Romero et al., 1995 a). Esta especie ha sido poco estudiada; no obstante, se conoce su taxonomía y un poco de su biología y pesquería (Castro-Aguirre, 1978; van der Heiden, 1985; Ramírez-Rodríguez, 1987a y 1987b; Cruz-Romero et al., 1989; Ramírez-Rodríguez y Rodríguez, 1990; Chávez-Ramos et al., 1994; Saucedo y Ramírez, 1994; Aguilar-Palomino et al., 1996). En la zona de estudio, se ha determinado la edad y el crecimiento de esta especie por medio del análisis indirecto de frecuencia de tallas (Cruz-Romero et al., 1993 y 1995b) y la relación entre parámetros poblacionales y ambientales (Espino-Barr, 2000). Sin embargo, no existen trabajos que analicen el crecimiento de esta especie por medio de datos basados en la determinación de la edad por métodos directos, los cuales corroborarían las estimaciones indirectas, y establecer relaciones talla-edad.
De acuerdo con lo anterior, en el presente estudio se analiza por primera vez el crecimiento en longitud y en peso de A. interruptus, empleando datos de edad determinados por medio de la lectura de escamas. Asimismo, se aportan resultados sobre el factor de condición, la longevidad, la mortalidad total, la sobrevivencia y la mortalidad natural, los cuales pueden aplicarse en algunos modelos de rendimiento para conocer los niveles de captura permisibles y determinar puntos de referencia biológicos para el manejo del recurso.
MATERIALES Y MÉTODOS
Durante 1986 se muestrearon 300 individuos de bacoco de la captura comercial en el litoral de Colima (Figura 1). La captura se lleva a cabo con línea de mano y redes agalleras de 7.5 a 15.2 cm de ojo de malla. Las tallas obtenidas fluctúan entre 8 y 44 cm de longitud estándar. Con base en el análisis del tamaño de muestra (Zar, 1996) se determinó que la muestra es confiable.

Los datos obtenidos para cada ejemplar fueron: longitud estándar en centímetros (Ls) y peso total en gramos (W). También se colectaron alrededor de 15 escamas de cada individuo, tomadas del área detrás de la aleta pectoral y por debajo de la línea lateral. Las escamas se limpiaron con agua jabonosa, se montaron entre dos portaobjetos y se observaron con luz transmitida, para contar las líneas de crecimiento. El análisis de estas estructuras permitió la identificación de nueve grupos de edad (edad 1- 12.52 cm, 2 – 17.47, 3 – 22.07, 4 – 26.35, 5 – 29.43, 6 – 32.03, 7 – 34.93, 8 – 36.96 y 9 – 38.76), y demostró que el crecimiento de las escamas es proporcional al del pez y que el tiempo de formación de los anillos de crecimiento es de un año (Gallardo-Cabello et al., 2003). Estas longitudes promedio fueron empleadas en la obtención de las constantes de la ecuación de von Bertalanffy (1938), por medio de los métodos de Ford (1933), Walford (1946), Gulland (1964) y Prager (1987). El mejor método se determinó a través del valor mínimo de la suma de las diferencias entre los valores observados y los estimados al cuadrado (∑ ei 2).
La función W = a Lb fue aplicada para obtener la relación peso-longitud, y la “a” (ordenada al origen) fue tomada como el factor de condición (Safran, 1992). El análisis de la significancia de los índices alométricos se llevó a cabo aplicando la prueba de t Student (Mendenhall, 1987; Daniel, 1991). Los datos sobre crecimiento en longitud y la relación peso-longitud fueron empleados para obtener el crecimiento en peso substituyendo Lt y L∞ por Wt y W∞, respectivamente.
La ecuación de Taylor (1959) se utilizó en la determinación de la edad límite o longevidad y la mortalidad natural (M). El valor de la mortalidad total (Z) se obtuvo por medio del método de la curva de captura (Sparre y Venema, 1995). La sobrevivencia (S) se calculó por medio de la aplicación de la función de S = e –Z (Ricker, 1975).
RESULTADOS Y DISCUSIÓN
Crecimiento en longitud
Los valores de los parámetros de la ecuación de crecimiento de von Bertalanffy, se muestran en la tabla 1, mientras que en la tabla 2 se observan los valores de la longitud para cada edad. Aunque los métodos presentaron un buen ajuste a los datos observados con respecto a los calculados, el método de Prager (1987) fue el que presentó el mejor ajuste en términos de menor sesgo. La curva de von Bertalanffy se observa en la figura 2.
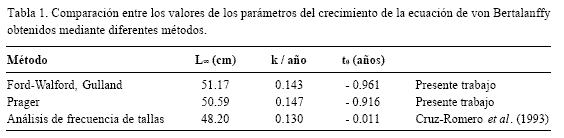
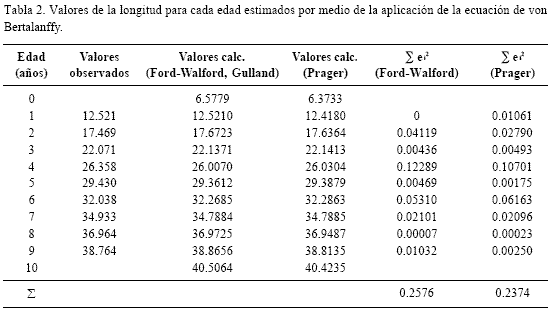
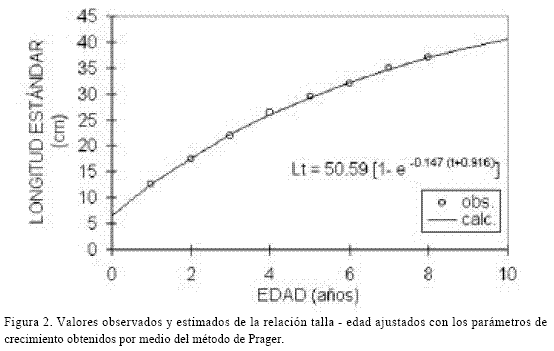
Los valores de los parámetros de la ecuación de von Bertalanffy para esta especie muestran un crecimiento lento (k = 0.147/año) en el cual el 95 % de la longitud asintótica se alcanza hasta los 22 años de edad aproximadamente. El mayor incremento del crecimiento en longitud se lleva a cabo durante el primer año de vida, en el que los ejemplares alcanzan una talla promedio de 12.42 cm. Posteriormente la tasa de crecimiento instantánea va disminuyendo gradualmente en relación con la edad, hasta alcanzar la longitud asintótica. La captura comercial está constituida en un 66% por preadultos, siendo los porcentajes de organismos en relación a la edad como sigue: 1 = 23.0, 2 = 22.1, 3 = 21.0, 4 = 16.2, 5 = 10.9, 6 = 4.6, 7 = 1.6, 8 = 0.5 y 9 = 0.1, lo que implica que la mayoría de la población no alcanza la edad de reproducción y puede, muy fácilmente, sobrepescarse.
No se observaron grandes diferencias entre los valores de los parámetros de la ecuación de von Bertalanffy obtenidos en el presente estudio (Tabla 2) y los expuestos por Cruz-Romero et al. (1993), aunque en este trabajo se reconoció el grupo de edad cero a diferencia del análisis mencionado que asigna el primer valor de la curva polimodal al grupo de edad 1. La diferencia entre los totales residuales entre los valores observados y los estimados es de apenas dos centésimas.
Crecimiento en peso
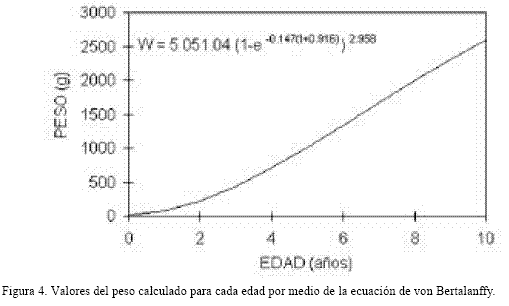
La relación peso longitud es: W = 0.046 x Ls 2.958 y la gráfica obtenida puede observarse en la figura 3. El índice alométrico b = 2.958 no mostró diferencia significativa a tres (t = 3.14, p > 0.05), lo que confirma que la relación entre el peso y la longitud es isométrica. La ecuación del crecimiento teórico en peso es útil para su conversión, en la aplicación de modelos de biomasa. En este caso se calculó como sigue: Wt = 5051.04(1 – e-0.147(t0+ 0.916))2.958

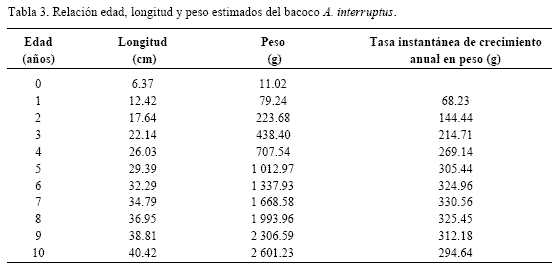
En la tabla 3 se observa la relación entre la longitud y el peso para cada edad y en la figura 4 se muestra la curva de crecimiento obtenida, donde se corrobora que la tasa instantánea aumenta conforme el pez incrementa su edad.
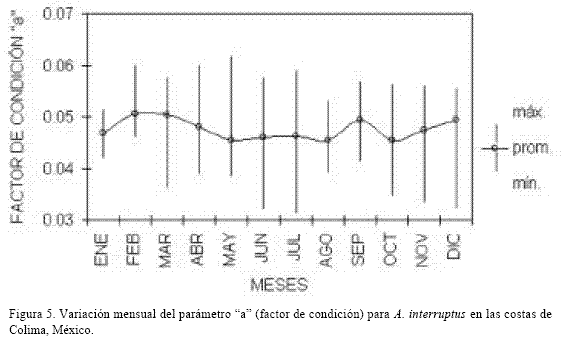
Factor de condición.
La tendencia de los valores del factor de condición obtenidos mensualmente y su variabilidad se observan en la figura 5. Los valores en el peso de los organismos muestran la tendencia a incrementarse durante los meses de enero a marzo y de octubre a diciembre, disminuyendo de abril a agosto. Este descenso puede representar períodos posteriores al desove en que el desgaste corporal del pez es grande (Rodríguez-Gutiérrez, 1992).
Longevidad.
La edad límite o longevidad es de 21.29 años, lo que significa que el pez alcanza el 95% de su longitud máxima teórica en este tiempo. Esta especie es longeva, lo que comúnmente sugiere que su crecimiento es lento (Sparre y Venema, 1995).
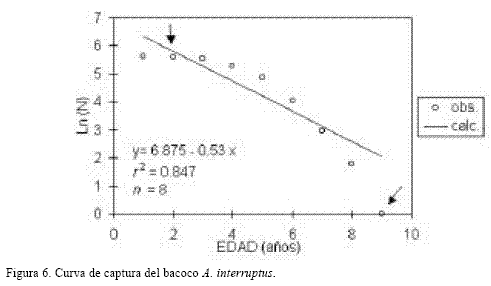
Mortalidad Total (Z).
En la figura 6 se observa la curva de captura obtenida, de la cual se estimó Z en 0.53/año, que no corresponde con los modelos que se observan tradicionalmente, constituidos por una rama ascendente y otra descendente a partir de la cual se obtiene el valor de la pendiente negativa. Ello es debido a que en el caso del bacoco los individuos se incorporan a la pesquería desde el primer año de edad, es decir no se protege a los individuos de menor talla que muy posiblemente no se han reproducido la primera vez. Lo anterior podría llevar a este recurso a un nivel de sobreexplotación, lo que no ha sucedido aún debido posiblemente a que en la pesca artesanal ribereña las capturas efectivas de esta especie no son lo suficientemente grandes como para provocar el agotamiento del recurso. O bien, en algunas épocas del año o inclusive años, disminuye la disponibilidad de los organismos a ser capturados, debido a los movimientos migratorios realizados por las poblaciones de esta especie hacia los fondos blandos (Aguilar-Palomino et al., 1996). También, cabe señalar que las mayores fluctuaciones de los recursos pesqueros ribereños en la costa de Colima, México son producidas por los factores ambientales y no por la explotación pesquera artesanal (Espino-Barr, 2000). El valor de la sobrevivencia (S) es de 0.589, lo que significa que en principio de cada 100 individuos, 41 mueren por mortalidad natural y por pesca.
La aplicación del uso de artes de pesca más selectivos que capturen organismos mayores a los tres años (edad de primera madurez) (Cruz-Romero, com. pers.), permitiría incrementar tanto la biomasa individual como la poblacional, a través del aumento de la longitud y del peso de los individuos, como lograr la protección de los organismos más jóvenes.
BIBLIOGRAFÍA
Aguilar-Palomino, B.; J. Mariscal-Romero; G. González-Sansón y L.E. Rodríguez-Ibarra. 1996. Ictiofauna demersal de fondos blandos de la plataforma continental de Jalisco y Colima, México, en la primavera de 1995. Ciencias Marinas, 22(4): 469-481. [ Links ]
Castro-Aguirre, J. L. 1978. Catálogo sistemático de los peces marinos que penetran a las aguas continentales de México, con aspectos zoogeográficos y ecológicos. PESCA Serie Científica, 19, 298 p. [ Links ]
Chávez-Ramos, H.; F. Galván-Magaña; L.A, Abitia-Cárdenas; J. de la Cruz-Agüero y J. Rodríguez-Romero. 1994. La ictiofauna marina de Baja California Sur, México, desde la perspectiva de un trabajo museológico. Inv. Mar. CICIMAR, 9(1): 43-49 [ Links ]
Cruz-Romero, M.; E. Espino-Barr y A. Garcia-Boa. 1989. Lista de peces del litoral colimense. INP/SEPESCA. Serie Doc de Trab., (9):21 p. [ Links ]
______.1993. Aspectos poblacionales de cinco especies de la familia Haemulidae (Pisces) en la costa de Colima, México. Cienc. Pesq., 10:43-54 [ Links ]
______.1995a. Pesquerías ribereñas del Pacífico mexicano. 28 p. En: Atlas Pesquero de México y Pesquerías Relevantes. SEPESCA/INP, CD.-ROM [ Links ]
______.1995b. La pesca ribereña en el estado de Colima. Estudios Jaliscienses, 20:14-26 [ Links ]
Daniel, W.W. 1991. Bioestadística. Base para el análisis de las ciencias de la salud. Ed. Noriega-Limusa, México, 667 p. [ Links ]
Espino-Barr, E. 2000. Criterios biológicos para la administración de la pesca multiespecífica artesanal en la costa de Colima, México. Tesis Doctoral, Univ. de Colima, Tecomán, Colima, México, 154 p. [ Links ]
Ford, E. 1933. An account of the herring investigations conducted at Plymouth during the years from 1924 to 1933. J. Mar. Biol. Assoc. U. K., 19:305-384. [ Links ]
Gallardo-Cabello, M.; E. Espino-Barr; F. González-Orozco y A. Garcia-Boa. 2003. Age determination of Anisotremus interruptus (Gill, 1863) (Perciformes: Haemulidae) by reading scales, in the coast of Colima, Mexico. Rev. Biol. Trop., 51(2):519-528. [ Links ]
Gulland, J.A. 1964. Manual of methods of fish population analysis. FAO Fish. Tech. Paper., 40, 60 p. [ Links ]
Mendenhall, W. 1987. Introducción a la probabilidad y la estadística. Ed. Grupo Editorial Iberoamérica, México, 628 p. [ Links ]
Prager, M.H. 1987. Nonlinear parameter estimation for fisheries (FISH-PARM), version 2.1S. Elsevier Scientific Publ. Co., software. Norfolk, VA, USA. [ Links ]
Ramírez-Rodríguez, E. M. 1987 a. Análisis preliminares de las pesquerías artesanales del área de Bahía Magdalena, B. C. S., durante 1982 y 1983. Mem. V Simp. Biol. Mar. La Paz Baja Calif. Sur, 149- 154 [ Links ]
______.1987 b. Abundancia relativa de peces demersales en el Golfo de California durante 1979. Inv. Mar. CICIMAR, 3(2): 31-42 [ Links ]
Ramírez-Rodríguez., M. y C. Rodríguez-M. 1990. Composición específica de la captura artesanal de peces en Isla Cerralvo, B. C. S., México. Inv. Mar. CICIMAR, 5(2): 137-141 [ Links ]
Ricker, W.E. 1975. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Can., (191): 382 p. [ Links ]
Safran, P. 1992. Theoretical analysis of the weight-length relationship in fish juveniles. Mar. Biol., 112: 545-551 [ Links ]
Saucedo, J. C. y M. Ramírez 1994. Peces de importancia comercial en el sur del estado de Sinaloa, México (pesca artesanal). Inv. Mar. CICIMAR, 9(1): 51-54. [ Links ]
Sparre, P. y S. C. Venema 1995. Introducción a la evaluación de recursos pesqueros tropicales. Parte 1 - Manual. FAO Doc. Tec. de Pesca, 306/1, Roma, 420 p. [ Links ]
Taylor, C.C. 1959. Temperature and growth. The Pacific razor clam. J. Conseil, 25(1):93-101 [ Links ]
Van der Heiden, A. M. 1985. Taxonomía, biología y evaluación de la ictiofauna demersal del Golfo de California. p. 149-200. In: A. Yáñez-Arancibia, (Ed.) Rec. Pesq. Pot. de México. La pesca acompañante del camarón. Pub. Inst. Cienc. del Mar Limnol., UNAM, INP/SEPESCA, México [ Links ]
von Bertalanffy, L. 1938. A quantitative theory of organic growth (inquiries on growth laws. II). Human Biol., 10(2):181-213. [ Links ]
Walford, L.A. 1946. A new graphic method of describing the growth of animals. Biol. Bull., 90(2):141-147. [ Links ]
Zar, J.H. 1996. Biostatistical analysis. 3rd ed. Prentice Hall. New Jersey, USA., 662p. [ Links ]
FECHA DE RECEPCIÓN: 08/09/04 FECHA DE ACEPTACIÓN: 04/06/04
DIRECCIÓN DE LOS AUTORES
CRIP-Manzanillo, INP, Playa Ventanas s/n, Manzanillo, Colima, 28200, México, Tel: (01) 33 32 37 50; Fax: (01) 33 32 37 51; E-mail: elespino@bay.net.mx (E.E.B.); escama@webtelmex.net.mx (F.G.O. y A.G.B.). Instituto de Ciencias del Mar y Limnología, UNAM, Apartado Postal 70-305, C.P. 09340, México, D.F., E-mail: gallardo@mar.icmyl.unam.mx (M.G.C).
