










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.34 no.1 Santa Marta Dec. 2005
Silvia Martínez1 y Alberto Acosta2
1Pontificia Universidad Javeriana. Facultad de Ciencias. Departamento de Biología. Cra. 7 No. 43 – 82. Edificio 53, oficina 106 B. Bogotá, Colombia. Correo-e: s1512145@student.rug.nl
2Pontificia Universidad Javeriana. Facultad de Ciencias. Departamento de Biología. Cra. 7 No. 43 – 82. Edificio 53, oficina 106 B. Bogotá, Colombia. Correo-e:laacosta@javeriana.edu.co
RESUMEN
En la última década varias predicciones han sido planteadas, acerca de la magnitud y dirección de los cambios temporales en la estructura coralina, pero pocos estudios las han demostrado cuantitativamente. Con el fin de evidenciar tales cambios, se cuantificó la estructura de la comunidad coralina en cuatro arrecifes del Caribe colombiano, dos del área de Santa Marta: Punta Betín (PB) e isla Morro Grande (MO) y dos en el Parque Nacional Natural Tayrona (PNNT): ensenada de Granate (GR) y bahía Gayraca (GA). La composición y cobertura coralina fueron cuantificadas durante el 2002 y comparadas con estudios de línea base llevados a cabo en 1989 (PB, MO y GR) y 1992 (GA). Estas variables se cuantificaron mediante transectos lineales con cadena de 20 m, entre 1.5 m y 29 m de profundidad. Después de una década, todos los arrecifes perdieron, en promedio, 13.8 % de la cobertura coralina absoluta (respecto al total del fondo) y 42 % de la relativa (respecto al total de coral). La composición coralina cambió en el tiempo en los 4 arrecifes estudiados, disminuyendo el número de géneros y aumentando el número de especies. El número de géneros cambió entre 8 y 18 % y el número de especies entre 27 y 68 %, debido a la presencia/ausencia de especies en los muestreos. Sin embargo, ni la riqueza ni la diversidad mostraron diferencias significativas en cada uno de los arrecifes. Montastrea cavernosa y M. annularis siguen siendo las especies dominantes con coberturas relativas superiores al 20 %. La riqueza y la cobertura absoluta de coral vivo cuantificadas en el 2002 siguieron el gradiente de deterioro registrado en el pasado, siendo menores en los arrecifes del área de Santa Marta y mayores en los del PNNT. Sin embargo, el porcentaje absoluto de pérdida de coral siguió un patrón inverso en el cual el arrecife de PB fue el menos afectado (6 %), seguido por MO (12.6 %), GR (14.6 %) y GA el más afectado (22 %). El coral muerto en PB y MO fue reemplazado por esponjas cuya cobertura aumentó en el tiempo, mientras que en GR y GA el coral muerto no ha sido colonizado por otros grupos. La pérdida de cobertura de los corales escleractíneos sugiere un cambio temporal significativo en la estructura de la comunidad hacia un estado de mayor deterioro, que puede estar alterando la función del arrecife.
PALABRAS CLAVE: Arrecifes coralinos, Cambio temporal, Caribe colombiano, Degradación, Estructura de la comunidad coralina.
ABSTRACT
Several predictions have been made, about magnitude and direction of temporal changes in the coral community structure, during the last decade. However, few studies have demonstrated it quantitatively. In order to document such changes, the coral community structure was studied in four continental reefs in the Colombian Caribbean, two in the Santa Marta region: Punta Betín (PB) and Morro Grande Island (MO), and two in the Tayrona National Natural Park (PNNT): Granate cove (GR) and Gayraca bay (GA). Coral composition and cover were quantified in 2002 and compared to base line studies from 1989 (PB, MO & GR) and from 1992 (GA). These variables were quantified by 20 m chain linear transects between 1.5 m and 29 m depth. After a decade, all reefs lost in average 13.8 % (with respect to the total of the bottom) absolute coral cover and 42 % relative coral cover (with respect to the total of coral bottom). Composition changed through time in the four sampled reefs by decreasing the number of genera and increasing the number of species. The change in genera ranged between 8 and 18% and in species between 27 and 68 % due to the presence/absence of species in samples. Nevertheless, neither the richness nor the diversity showed significant differences for any of the reefs. Montastrea cavernosa y M. annularis are still the dominant species in the community with relative coral cover higher than 20 %. Quantified richness and absolute coral cover during 2002 followed the degradation gradient documented in the past, being the values smaller in the Santa Marta’s reefs and higher in the Tayrona´s. However, absolute percentage coral cover lost showed an inverse pattern, being PB (6 %) the least affected, followed by MO (12.6 %), GR (14.6 %) and being GA (22 %) the most affected. Dead coral in PB and MO was replaced by sponges which cover increased significantly over time; meanwhile in GR and GA, dead coral doesn’t seem to have been replaced by other groups. The scleractinian coral cover lost suggests a significant temporal change in the community structure toward a grater degradation state, which might be shifting reef’s function.
KEY WORDS: Colombian Caribbean, Coral community structure, Coral reefs, Degradation, Temporal change.
INTRODUCCIÓN
Debido a la presencia y al desarrollo de los arrecifes coralinos cerca a las zonas costeras, se ha observado un rápido proceso de degradación de estos ecosistemas por factores, principalmente de origen antropogénico, tales como la sobrepesca, el aumento en la sedimentación, la entrada de nutrientes, la extracción de corales y arena, y la destrucción de colonias coralinas por actividades como el buceo recreativo (Ramírez et al., 1986; Díaz et al., 1995; Connell, 1997; GCRMN, 2000). Tal deterioro ha sido evidenciado a nivel global con aumento en la mortalidad coralina (alcanzando valores mayores al 90 % en algunos arrecifes), cambio en la composición de especies, reducción en la tasa de calcificación, presencia de enfermedades (Porter et al., 2001) e incluso un reemplazo de comunidades dominadas por corales hacia otras dominadas por algas u otros organismos bentónicos no constructores de arrecifes (Hallock et al., 1996).
Las comunidades coralinas fósiles, cuando han sido estudiadas en una amplia escala temporal (miles de años), muestran una estructura determinada y dinámica persistente y predecible (Pandolfi, 1999; 2002), dado su elevado grado de orden en distribución de especies. Sin embargo, al ser estudiadas a escalas temporales reducidas (décadas), muestran un alto grado de variabilidad en la estructura, donde las frecuentes perturbaciones y fluctuaciones ambientales evitan que exista un equilibrio temporal (Connell, 1978; Pandolfi, 2002). Actualmente la mayoría de investigaciones concluyen que aunque las modificaciones ambientales a gran escala (cambio climático global) han deteriorado de forma importante la estructura y la función de las comunidades coralinas, perturbaciones inducidas por el hombre a menor escala durante las últimas décadas, han tenido efectos tan rápidos que no han permitido la adaptación de los corales (Ginsburg, 1994; Pandolfi, 2002).
Los arrecifes coralinos de la región de Santa Marta – PNNT son de tipo franjeante simple a sotavento (al oriente de las bahías), por lo general poco amplios, que se profundizan hasta los 25 m-30 m, presentando una estructura de bajo desarrollo y han sido descritos en diferentes estudios desde hace más de cuatro décadas por Antonius (1972), Erhardt y Werding (1975), Solano (1987), Acosta (1989), Werding y Sánchez (1989), Zea y Duque-Tobón (1989), Garzón-Ferreira y Cano (1991) y Garzón-Ferreira (2001), entre otros. Algunos de estos autores han planteado que el bajo crecimiento coralino y desarrollo de los arrecifes está limitado por condiciones subóptimas, como aguas de surgencia y la descarga continental de aguas dulces cargadas de sedimentos y nutrientes provenientes principalmente de la Ciénaga Grande de Santa Marta (que recibe aportes del río Magdalena) y de forma secundaria de los ríos Gaira, Manzanares y las descargas de aguas residuales del Boquerón (Antonius, 1972; Werding y Sánchez, 1988; 1989; Zea, 1993). Estas aguas generan un gradiente de perturbación que se mueve hacia el noreste afectando las formaciones coralinas del área de Santa Marta y del PNNT, disminuyendo la salinidad y aumentando la turbidez, la sedimentación, la temperatura y los niveles de nutrientes y de ciertos contaminantes en el mar (Erhardt y Werding, 1975; Acosta, 1989). Este gradiente, junto con otros factores como la pesca con dinamita, el turismo intensivo y las enfermedades han degradado paulatinamente los arrecifes del área durante las últimas décadas (Garzón-Ferreira y Cano, 1991). Por lo anterior, es urgente monitorear estas comunidades coralinas con el fin de establecer la magnitud del cambio y la influencia de los diferentes agentes de perturbación en el área.
En el área de Santa Marta – PNNT, únicamente en bahía Chengue se ha llevado a cabo un programa de monitoreo durante varios años por parte del CARICOMP (Caribbean Coastal Marine Productivity) y el SIMAC (Sistema Nacional de Monitoreo de Arrecifes Coralinos en Colombia), en un arrecife en buen estado de conservación con respecto a otros del área (ello, presumiblemente debido a su aislamiento y mayor distancia a las fuentes de perturbación). Los resultados de tal estudio no han indicado grandes cambios en la cobertura coralina durante los últimos años (Garzón-Ferreira, 2001). Sin embargo, los datos de un arrecife no necesariamente son representativos de los cambios que se pueden estar presentando en todos los arrecifes del área, ya que el deterioro coralino ha sido observado de forma cualitativa por varios investigadores en las últimas décadas. A pesar de ello, hasta ahora no existen monitoreos en otros arrecifes del área, expuestos a diferentes niveles de perturbación, que permitan demostrar cuantitativamente los cambios sufridos por las comunidades coralinas en el tiempo.
Acosta (1989) evaluó la estructura de las comunidades coralinas de tres arrecifes (Punta Betín, Isla Morro Grande y Ensenada de Granate) y de uno en 1992 (Bahía Gayraca), todos representativos del área de Santa Marta – PNNT. El autor encontró que el grado de desarrollo arrecifal y el estado de salud de la comunidad de corales estaban relacionados con su estructura y que ésta, a su vez, reflejaba un gradiente que coincidía con la calidad del agua. De esta forma, la estructura coralina resultó ser menos compleja en los arrecifes más cercanos a la Bahía de Santa Marta y más compleja en aquellos pertenecientes al PNNT (alejados de la fuente de perturbación). El objetivo del presente estudio fue evaluar las comunidades coralinas en estos mismos cuatro arrecifes después de una década, con el fin de establecer si su estructura presenta cambios en el tiempo, así como la magnitud y dirección de estos cambios.
ÁREA DE ESTUDIO
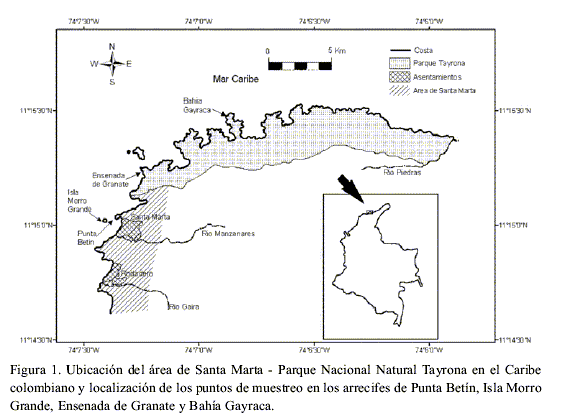
El área de estudio se ubica al noreste de Colombia, en el departamento del Magdalena, sobre la costa Caribe y comprende el área de Santa Marta y PNNT entre las coordenadas 11°15´- 11°22´ N y 73°57´- 74°12´ W (Figura1). El sector de Santa Marta incluye las bahías de Gaira, Santa Marta y Taganga (Díaz et al., 2000), en tanto que el del PNNT está entre Punta Minas y la desembocadura del río Piedras. El litoral costero es de carácter rocoso y se caracteriza por bahías y ensenadas alternadas con playas arenosas y cabos e islotes rocosos, que abarcan 85 km de costa, estando expuesto a fuertes vientos y corrientes (Brattström, 1980; Díaz et al., 2000; Brüggenmann et al., 2002).
El clima es tropical, con una temperatura que oscila entre 25 °C y 30°C y una precipitación promedio anual de 500 mm; la temperatura del agua oscila entre 21°C y 29°C y la salinidad entre 24.8 y 37 (Acosta, 1989; De Jongh, 1990). A lo largo del año se pueden distinguir cuatro estaciones; una seca principal, una lluviosa menor, una seca menor y una lluviosa mayor. Durante la época seca mayor (diciembre a febrero), la corriente principal se dirige hacia el suroeste debido a los vientos alisios del noreste, los cuales adicionalmente causan surgencia que disminuye la temperatura del agua hasta 21°C y aumenta la salinidad hasta 37. Por el contrario, durante la época lluviosa mayor (septiembre a noviembre), los vientos del suroeste causan una contracorriente en dirección al noreste, que transporta aguas continentales de efluentes como el río Magdalena, la Ciénaga Grande de Santa Marta y ríos de menor caudal (Gaira y Manzanares), causando en el área de estudio los máximos valores de temperatura del agua (31°C) y de turbidez (10 cm de visibilidad de disco sechi), y los mínimos de salinidad (24.8; Prahl y Erhardt, 1985; Acosta, 1989; De Jongh, 1990).
Estaciones de muestreo
El área de estudio comprende dos formaciones arrecifales pertenecientes al sector de la bahía de Santa Marta (Punta Betín y Morro Grande), y dos pertenecientes al sector del Parque Nacional Tayrona (Granate y Gayraca; Figura 1).
Punta Betín es una península rocosa de pendientes escarpadas, ubicada al norte de la bahía de Santa Marta, por lo que se ve directamente afectada por la descarga de aguas dulces (río Manzanares y emisario submarino) y el puerto (granos y polvillo de carbón). El arrecife es, de los estudiados, el que presenta la menor cobertura coralina, siendo dominado por Montastrea cavernosa que se distribuye ampliamente desde 8 m hasta 23 m de profundidad, y otras especies como Agaricia agaricites, Stephanocoenia intercepta, Madracis decactis y Millepora alcicornis. Los transectos cuantificados se localizaron al sur de la península, en la parte protegida (Acosta, 1989).
La isla Morro Grande se encuentra ubicada aproximadamente a 1.5 km de la costa, al oeste de Punta Betín, por lo que presenta mayor influencia de aguas oceánicas con respecto a los otros tres arrecifes estudiados. El arrecife se divide en una terraza arrecifal hasta los 20 m con grandes colonias de coral y un talud hasta los 30 m con corales foliáceos; es dominado por Diploria strigosa y M. cavernosa (Zea, 1993). Los transectos fueron localizados al suroeste de la isla, donde se encuentran las mejores formaciones coralinas (Acosta, 1989).
La ensenada de Granate es una bahía ubicada entre Cabo de la Aguja y Punta Granate, a 6.5 km al noreste de Santa Marta. Está relativamente bien protegida de los vientos alisios y del oleaje. El arrecife posee un desarrollo coralino desde los 5 hasta los 24 m de profundidad y se caracteriza por una extensa llanura arrecifal de poca pendiente, siendo M. cavernosa, M. annularis y C. natans las especies dominantes (Garzón-Ferreira y Cano, 1991). Los transectos se ubicaron al noreste de la bahía sobre un fondo poco pendiente (Acosta, 1989).
La Bahía de Gayraca se ubica entre las bahías de Chengue y Neguange. La mayor parte de la bahía se encuentra protegida de los vientos alisios. El arrecife posee un talud de fuerte pendiente con corales escleractíneos hasta 25 m de profundidad, donde se destacan las especies M. annularis, M. cavernosa y C. natans (Garzón-Ferreira y Cano, 1991). Los transectos se ubicaron en la parte protegida de la bahía cerca al extremo noreste (Acosta y Zea, 1997).
MATERIALES Y MÉTODOS
La fase de campo se llevó a cabo durante la época seca menor (finales de junio y principios de julio) en el 2002. La estructura de la comunidad coralina se cuantificó por medio de transectos lineales con cadena, siguiendo a Loya (1972), por ser la misma metodología utilizada por Acosta (1989) en Punta de Betín, Isla Morro Grande, Ensenada de Granate y por Acosta y Zea (1997) para el muestreo de 1992 en Bahía Gayraca. El número de transectos por arrecife y las profundidades de los mismos fueron iguales a los realizados en el pasado y dependieron de las discontinuidades en la estructura, de la composición de la comunidad coralina y del fondo arenoso como límite vertical para la formación coralina. A su vez, los transectos se ubicaron en los mismos sitios en cada arrecife, para lo cual se contó con perfiles batimétricos y cuerdas guía dejadas en el primer estudio de línea base (Acosta, 1989). Así, se cuantificaron un total de 11 transectos en Punta Betín (1.5, 4, 9.5, 8, 10.5, 14.5, 16.5, 18, 19.5, 21 y 22.5 m), 11 en Isla Morro Grande (4.5, 9, 10.5, 12, 14, 15, 15.5, 19.5, 22, 24 y 29 m), 16 en Ensenada de Granate (5, 6, 7.5, 8, 9, 11, 12, 14, 15, 18, 18.5, 19, 19.5, 20.5, 22 y 24 m) y 10 en Bahía Gayraca (6, 8, 10, 13, 15, 16, 18, 21, 23, 25 m), para un total de 48 transectos en cada período de tiempo.
Cada transecto se realizó tendiendo una cadena de 20 m, siguiendo el contorno del fondo, en posición paralela a la costa y perpendicular a una línea guía tendida sobre el sustrato desde la orilla hasta la parte profunda. El número y la longitud de los transectos constituyen una muestra significativa del número de especies que posee la comunidad en estos arrecifes, basándose en curvas de colector realizadas previamente por Solano (1987) y Acosta (1989). Para corroborar la suficiencia del muestreo en el presente se utilizó la prueba de la curva T-S propuesta por Ugland et al. (2003), la cual indicó que el número de transectos por arrecife fueron suficientes para muestrear, en el 2002, el 96, 95, 93 y 96 % de la riqueza de especies en los arrecifes de Punta Betín, Morro Grande, Granate y Gayraca respectivamente. En cada transecto se contó el número de eslabones ocupados por cada colonia de las diferentes especies de corales hermatípicos (identificados in situ y a la misma resolución taxonómica del pasado) y de otras categorías como esponjas, macroalgas, octocorales, arena, coral muerto y roca. Esta técnica es similar a la estandarizada actualmente por SIMAC y CARICOMP, lo que permite que la información sea comparada con otros arrecifes del Caribe.
Para el análisis temporal de los datos de estructura de la comunidad se compararon las matrices obtenidas en este estudio con las suministradas por Acosta (1989) para Punta Betín, Morro y Granate, y en 1992 para Gayraca.
Composición
Para analizar el cambio en la composición de especies en el tiempo, se compararon tanto las especies como los géneros presentes/ausentes para cada uno de los arrecifes entre los dos tiempos (pasado vs. presente). El porcentaje de cambio tuvo en cuenta el número de especies y géneros que se mantuvieron en cada arrecife en el tiempo vs. número de especies y géneros que entraron y salieron en una década. El análisis a nivel de género se llevó a cabo con el fin de obtener mayor confiabilidad en la determinación del cambio en la composición coralina y para evitar posibles problemas taxonómicos en la identificación de especies (ej. Agaricia spp., Madracis spp. y Scolymia spp.).
Cobertura
El porcentaje de cobertura absoluta (con respecto al total del fondo) de cada categoría (coral vivo, esponjas, bases de octocorales, macroalgas, coral muerto y roca, arena) se comparó entre años para cada uno de los arrecifes. Dentro de la categoría de coral muerto se incluye todo el sustrato duro no colonizado por otras categorías ya mencionadas, aunque en su mayoría se refiere a coral muerto colonizado por cualquier tipo de algas diferentes a macroalgas (i.e. algas costrosas y céspedes algales). La comparación de la cobertura de coral en el tiempo se llevó a cabo tanto para el arrecife como un todo, como entre intervalos de profundidad (somero: <9 m, medio: 9-18 m y profundo: >18 m). Así mismo se determinó cuáles especies de corales hermatípicos presentaron cambios significativos de cobertura en el tiempo (pasado vs. presente), para lo cual se tomaron los promedios de cobertura de cada especie en cada arrecife. Para estos análisis se realizaron pruebas T-student pareadas entre los valores de cobertura entre los dos años estudiados, siendo la cobertura por transecto la unidad de muestra y normalizando la variable por medio de la transformación arcoseno raíz de p (siendo p la proporción de la categoría).
Riqueza
De igual forma, se tomó el número de especies encontradas por transecto en cada arrecife, comparándolas en el tiempo por medio de una prueba T-student.
Diversidad
Los valores de diversidad de cada uno de los arrecifes se calcularon utilizando cuatro índices diferentes y complementarios: Fisher Alpha, Dominancia de Simpson, diversidad y equitatividad de Shannon–Wienner. Para observar si las diferencias en el tiempo fueron estadísticamente significativas en cada arrecife, se aplicó la prueba T para varianzas al índice de diversidad de Shannon (Magurran, 1998). Adicionalmente, para comparar la estructura de los arrecifes en el pasado y en el presente, y simultáneamente los ocho arrecifes (4 en el pasado y 4 en el presente), se realizó un análisis exploratorio de clasificación normal mediante un cluster de similaridad de Bray-Curtis, utilizando la técnica de ligamiento de promedio de grupos.
RESULTADOS
Composición
La composición de la comunidad coralina cambió a nivel de cada arrecife (de 31 % a 46 % de especies y de 17 % a 22 % de géneros) y para toda el área de estudio (10 % para géneros y 34 % para especies, al agrupar los cuatro arrecifes, Tabla 1). En los cuatro arrecifes aumentó el número de especies pero disminuyó el de géneros (Tabla 1).
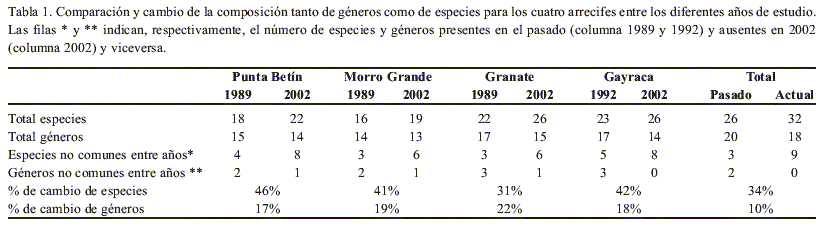
La comunidad de Punta Betín fue la que más cambió en el tiempo a nivel de especies (46 %) pero la que menos cambió a nivel de género (17 %); mientras que la comunidad de Granate fue la que menos cambió a nivel de especie (31 %) y más a nivel de género (22 %, Tabla 1).
Cobertura
La cobertura relativa de coral disminuyó entre 30 y 45 % en la última década (10 años para el arrecife de Gayraca y 13 años para los otros tres arrecifes, Tabla 2). Por su parte, la pérdida de cobertura coralina en términos absolutos (respecto al total del fondo) fue de 13.8 % en promedio para los cuatro arrecifes (Tabla 2). En términos absolutos, Gayraca fue el arrecife en el que se redujo más la cobertura coralina (22 %) y Punta Betín fue el arrecife con menos disminución (6 %). En términos relativos la cobertura coralina se redujo más en Morro Grande (45.1 %) y Gayraca (44.9 %), y en menor porcentaje en el arrecife de Punta Betín (29.7 %, Tabla 2).

Sin embargo, en la actualidad la cobertura total de coral vivo sigue siendo mayor en Gayraca, seguida por Granate, luego por Morro Grande y por último Punta Betín, tal y como se encontró en el pasado (Tabla 2).
Punta Betín
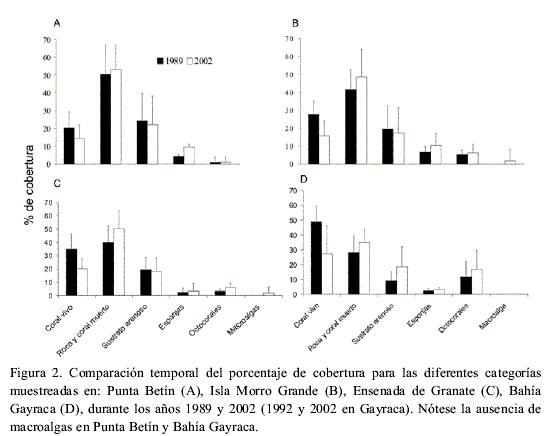
Aunque en Punta Betín se redujo la cobertura total de coral vivo en un 6 % entre 1989 y 2002 (Figura 2A), esta diferencia no fue estadísticamente significativa en el tiempo (N=11, P=0.098). Por el contrario, la cobertura de esponjas aumentó más del doble de manera significativa (N=11, P < 0.001). La cobertura de las categorías de coral muerto y roca, arena y octocorales no cambiaron en el tiempo de forma significativa (N=11, P>0.05, Figura 2A).
Al comparar el promedio de cobertura de coral vivo por intervalos de profundidad entre 1989 y 2002, se observó que solamente la disminución en la parte media del arrecife resultó significativa y fue de casi un 14 % (N=5, P=0.033). El porcentaje de cobertura de coral vivo se redujo en 8 de 11 transectos, mientras que en los transectos someros de 1.5 m y 4 m la cobertura coralina aumentó (Figura 3A y B). En 1989 la cobertura de coral vivo fue menor en los transectos someros (1.5 m y 4 m) y profundos (21 m y 22.5 m) (Figura 3 A), mientras que en 2002 la mayor cobertura se observó a 14.5 m y la menor en los transectos profundos (Figura 3 B). A 9.5, 10.5 y 16.5 m de profundidad la reducción en la cobertura de coral vivo fue mayor al 20 %. Los datos anteriores indican que el patrón de cobertura de coral con respecto a la profundidad cambió en el tiempo, ya que en 1989 se observaba una mayor cobertura en la zona media del arrecife (9.5 a 18 m), mientras que en 2002 no se presentó un patrón claro entre la cobertura de coral vivo y la profundidad.
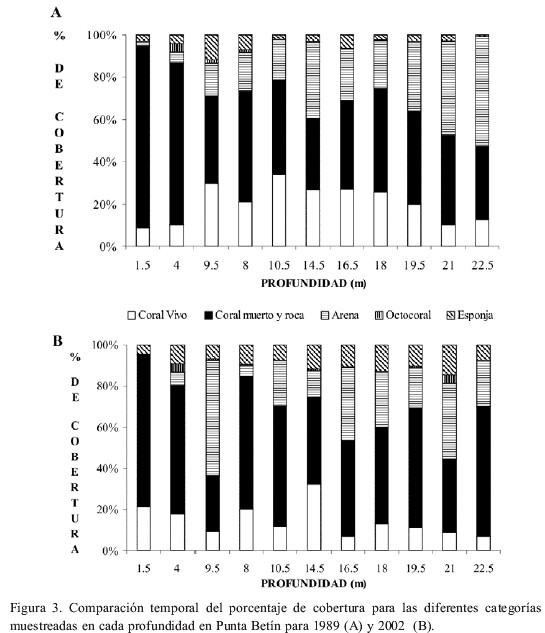
La cobertura de esponjas aumentó en 10 de 11 transectos entre 1989 y 2002, siendo el transecto ubicado a 9.5 m, el único en el que se observó disminución para esta categoría (Figura 3 A y B). Los octocorales no presentaron cambios entre años, ni un patrón claro entre su cobertura y la profundidad.
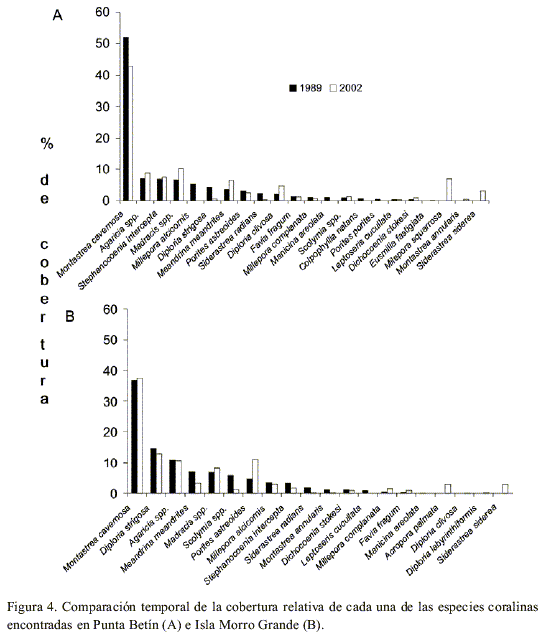
< p> En Punta Betín solamente Siderastrea radians presentó una disminución de cobertura relativa (2.1%) estadísticamente significativa en trece años (N=11, P=0.044). De las otras 22 especies, solamente 4 (M. cavernosa, D. strigosa, P. astreoides y Millepora complanata) disminuyeron su cobertura relativa en el 2002 (Figura 4A), aunque tal cambio no fue estadísticamente significativo (P>0.05). M. cavernosa, si bien sigue siendo la especie dominante, perdió importancia relativa con respecto a las otras especies (Figura 4A).
Isla Morro Grande
En el arrecife de Morro, la cobertura absoluta de coral vivo disminuyó de forma significativa en un 16 % entre 1989 y 2002 (n=11, P=0.002; Figura 2B), mientras que la de esponjas aumentó significativamente (n=11, P=0.027). Las categorías de coral muerto y roca, sustrato arenoso y octocorales no cambiaron significativamente en el tiempo (n=11, P>0.05). Las macroalgas, aunque con poco porcentaje (1.7 %), aparecen por primera vez en el arrecife en 2002, a 12 m de profundidad (Figura 2B).
La cobertura promedio de coral vivo se redujo de forma significativa con la profundidad, tanto en la parte media (casi 20 %, N=5, P=0.012) como en la profunda (2 %, N=3, P=0.032). La cobertura de coral vivo se redujo en los 11 transectos cuantificados, siendo mayor la pérdida a 10.5 m y 15 m, alcanzando valores superiores al 20 %, y menor a 14 m de profundidad (0.9%; Figura 5 A y B). En 1989 se observó una mayor cobertura en los transectos de profundidades intermedias, patrón que no se observó en 2002 (Figura 5 A y B).
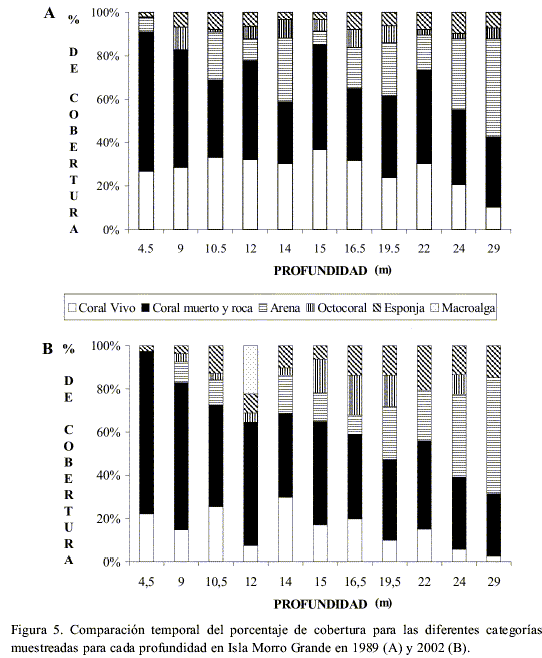
En el caso de las esponjas, la cobertura aumentó en 10 de 11 transectos entre 1989 y 2002, al igual que en Punta Betín; siendo 9 m la única profundidad en que esta categoría disminuyó su cobertura (Figura 5 A y B). La cobertura de octocorales, arena y coral muerto y roca tampoco presentó cambios en el tiempo, relacionados con la profundidad.
Siderastrea radians fue el único coral cuya cobertura presentó diferencias significativas en el tiempo, reduciéndose en 1.7 % (n=11, P=0.020). Por otra parte, siete corales escleractíneos disminuyeron su cobertura relativa (D. strigosa, M. meandrites, Scolymia spp., Millepora alcicornis, Stephanocoenia intercepta, M. annularis, D. stokesi) y 5 la aumentaron (M. cavernosa, Madracis spp., P. astreoides, M. complanata, F. fragum, Figura 4B), aunque de forma no significativa (P>0.05).
Ensenada de Granate
Entre 1989 y 2002, la cobertura de coral vivo se redujo significativamente en un 14.6 % (n=16, P<0.001, Figura 2C), mientras que la de coral muerto y roca aumentó (n=16, P=0.045). El resto de categorías no presentaron diferencias significativas (Figura 2C). Las macroalgas aparecieron en el arrecife en el 2002 con una cobertura de 1.9 %.
La cobertura absoluta de coral vivo, al ser analizada por intervalos de profundidad dentro del arrecife, mostró una reducción de 12 %, 15 % y 16 % en las zonas somera, media y profunda respectivamente, siendo la zona media la única en la que tal reducción fue significativa (n=6, P=0.002). La cobertura absoluta de coral vivo se redujo en 13 de los 16 transectos cuantificados, y en siete de ellos la reducción fue mayor a un 20 % (Figura 6 A y B). La mayor reducción se presentó a 12 y 20.5 m. Por el contrario, a 6, 8 y 19.5 m la cobertura de coral vivo aumentó ligeramente (<2%), entre 1989 y 2002. En 1989 la mayor cobertura coralina se encontraba a profundidades intermedias (Figura 6 A), patrón que cambió en el 2002 (Figura 6 B).
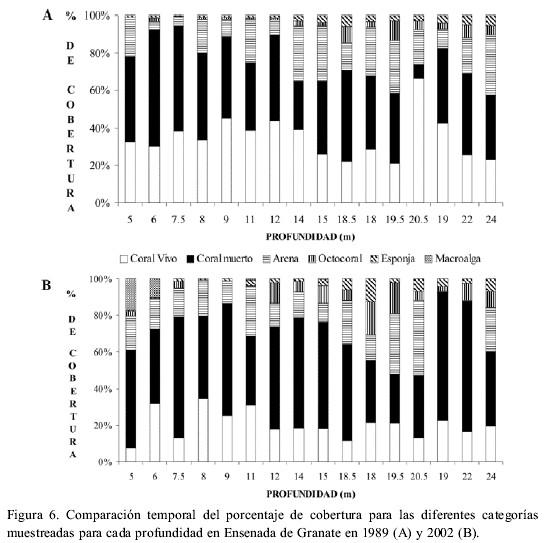
La cobertura de coral muerto y roca aumentó en doce de los 16 transectos cuantificados entre 1989 y el 2002; tanto octocorales como esponjas incrementaron su cobertura en 9 y 10 transectos respectivamente (Figura 6 A y B).
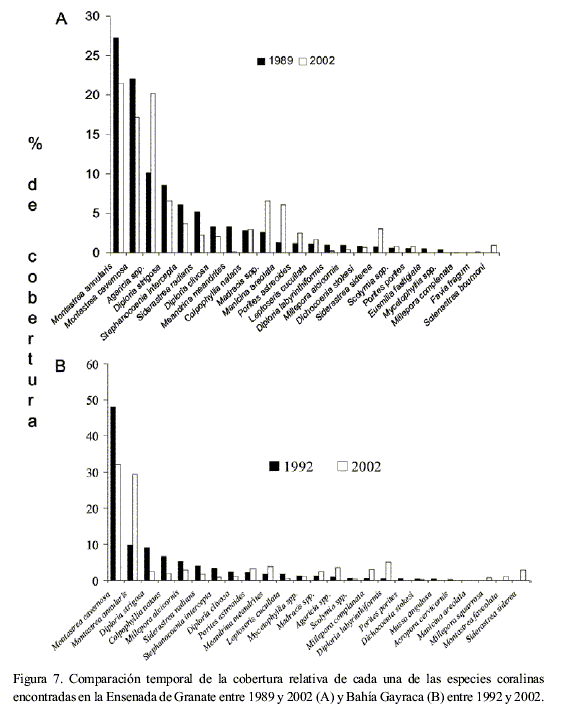
De las 22 especies coralinas cuantificadas, tres redujeron su cobertura relativa en el tiempo de forma significativa: M. cavernosa (n=16, P<0.001), M. alcicornis (n=16, P=0.026) y S. intercepta (n=16, P=0.004). Adicionalmente, 10 especies aumentaron su cobertura relativa en 2002, y siete más la disminuyeron, aunque tal cambio no fue significativo estadísticamente. Las especies dominantes en el pasado, M. annularis y M. cavernosa, fueron las que exhibieron mayor reducción en su cobertura relativa, especialmente la segunda. Por su parte, el género Agaricia dobló su porcentaje de cobertura relativa en el tiempo (Figura 7A).
Bahía Gayraca
La cobertura absoluta de coral vivo se redujo casi a la mitad después de una década (Figura 2D), siendo de todas las categorías cuantificadas, la única que cambió de forma significativa (n=10, P=0.004). Esta pérdida en cobertura de coral vivo se compensó con un aumento en el porcentaje de las otras categorías como coral muerto y roca, sustrato arenoso y octocorales, y en menor proporción de esponjas (Figura 2D), aunque los aumentos para estos cuatro grupos no fueron estadísticamente significativos en el tiempo (n=10, P>0.05).
La cobertura absoluta de coral vivo se redujo en casi un 35 % en la parte profunda del arrecife, mientras que en las partes somera y media fue de 21 % y 15 % respectivamente (Figura 8 A y B). Sin embargo, tal reducción sólo fue significativa en las partes somera y profunda (somera: n=3, P=0.036; profunda: n= 3, P = 0.001). Una reducción en la cobertura de coral vivo se notó en los diez transectos cuantificados, siendo mayor al 30 % a 21, 23 y 25 m de profundidad. A diferencia de los otros arrecifes, el patrón de mayor cobertura coralina a profundidades intermedias se mantuvo en el tiempo, a pesar de la pérdida de cobertura en 2002.
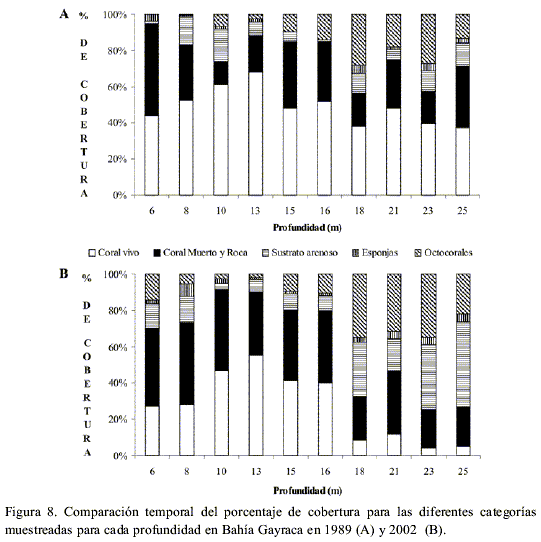
La cobertura de arena aumentó en siete de los diez transectos cuantificados, y disminuyó tan solo en los transectos profundos. Los octocorales aumentaron su cobertura en 6 transectos, pertenecientes a las zonas someras y profundas del arrecife (Figura 8 A y B).
La reducción de cobertura fue estadísticamente significativa en el tiempo para M. cavernosa (n=10, P<0.001), Colpophylia natans (n=10, P=0.049), D. strigosa (n=10, P=0.026) y Leptoseris cucullata (n=10, P=0.040, Figura 7B). De las especies dominantes, M. cavernosa disminuyó su cobertura relativa en casi un 20 %, mientras que M. annularis la aumentó casi en la misma proporción (Figura 7B).
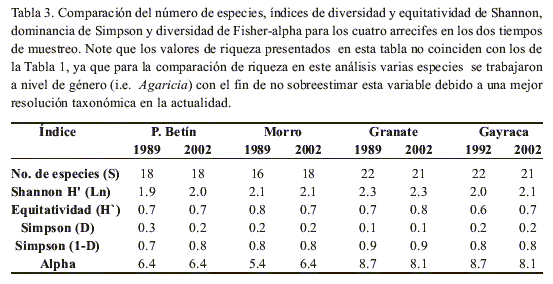
Riqueza y diversidad
Ningún arrecife presentó cambios en el tiempo al comparar los valores de riqueza, diversidad y dominancia (Tabla 3). La riqueza y la diversidad de Shannon no presentaron diferencias significativas al ser comparadas en el tiempo para ninguno de los arrecifes (P>0.05). Sin embargo, los arrecifes de Granate y Gayraca evidencian una mayor equitatividad en la actualidad, debido a la alta reducción en la cobertura relativa de especies dominantes. El índice de diversidad de Alpha indicó que Morro Grande presentó el mayor cambio relativo en el tiempo (aumento en diversidad), seguido por Granate y Gayraca (disminución de diversidad), y Punta Betín permaneciendo constante.
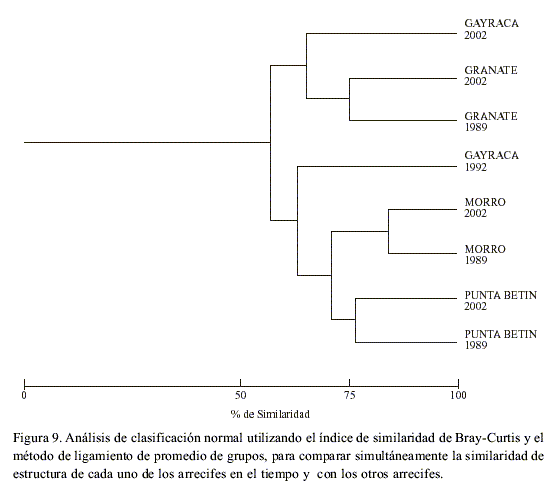
Clasificación
Al comparar la similaridad en estructura coralina de cada arrecife en el tiempo y con los otros arrecifes después de una década (Figura 9), se observó que aunque en general la estructura de las comunidades para todos y cada uno de los arrecifes mantuvo una similaridad superior al 50 %, éstas cambiaron en el tiempo en un alto porcentaje (disimilaridad >20 %). Según este análisis, la estructura de la comunidad coralina en el arrecife de Morro Grande fue la que menos cambió en el tiempo, seguida por las comunidades de Punta Betín y Granate, siendo en todos los casos la estructura de la comunidad más similar dentro de cada arrecife en el tiempo que entre arrecifes (Figura 9). Por el contrario, la estructura de la comunidad coralina en Gayraca fue la que presentó un mayor cambio en el tiempo (>35 %), ya que no se agruparon los arrecifes de 1992 y 2002 entre sí, sino que la estructura de la comunidad coralina de Gayraca en 1992 resultó más similar a Punta Betín y Morro; y en el 2002 a Granate (Figura 9).
DISCUSIÓN
Es evidente que la comunidad arrecifal de la región de Santa Marta – PNNT se ha deteriorado en la última década. Este deterioro se evidenció en el cambio en la estructura de la comunidad, expresado principalmente en las variables de composición y cobertura coralina, las cuales mostraron diferencias significativas en el tiempo. Esto comprueba lo propuesto por Pandolfi (1999; 2002) y Pandolfi y Jackson (1997), quienes argumentan gran variabilidad de las comunidades a pequeñas escalas temporales (décadas, como es el caso de este estudio).
El cambio en el tiempo de la cobertura coralina en un arrecife se considera especialmente importante, ya que de acuerdo con Hughes y Tanner (2000) la cobertura es el atributo más usado para describir el estado de un arrecife, por lo que su pérdida es considerada como crítica en el mantenimiento y función de la comunidad coralina y del arrecife como un todo. La reducción en cobertura de coral vivo ha sido uno de los principales indicadores del deterioro coralino en el mundo (Ginsburg, 1994). En general, para los cuatro arrecifes muestreados en este estudio, se registra una reducción relativa en la cobertura coralina del 42 % durante la última década (equivalente a una pérdida anual de 2.3 % en PB, 3.5 % en MO, 3.2 % en GR y 4.5 % en GA), cifras similares a las encontradas en las últimas décadas en otros arrecifes del Caribe. Algunos ejemplos son los registrados por Hughes (1994), quien documentó la pérdida de tejido coralino vivo de un 70 % a valores menores al 10 % en un periodo aproximado de una década en arrecifes de Jamaica; Hughes y Tanner (2000) indicaron una reducción en la cobertura coralina en 16 años de 52 % a 3 % en la Florida, y Porter y Porter (2002) para los cayos de la Florida cuantificaron una reducción del 38 % de cobertura coralina entre 1996 y 1999, con una tasa promedio de pérdida de cobertura de 13 % al año. Por su parte, Caro-Zambrano (2003) cuantificó en la parte media de Punta Betín una reducción en la cobertura coralina de 2.1 % entre 1998 y 2002, pero una cobertura constante en la parte media de Morro durante los mismos años. Sin embargo, Rodríguez-Ramírez y Garzón-Ferreira (2003) monitorearon la comunidad coralina de Bahía Chengue durante un período de 7 años, cuantificando una cobertura coralina inicial de 34.5 %, la cual permaneció constante entre 1993 y 1999. Este valor no solamente es mayor a los cuatro arrecifes de la zona, sino también a los otros arrecifes monitoreados en el Caribe (Caro-Zambrano, 2003). Según los autores, el arrecife en Bahía Chengue se encuentra en un mejor estado de conservación con respecto a otros arrecifes de la zona. No obstante, este monitoreo (Rodríguez-Ramírez y Garzón-Ferreira, 2003) solamente cubrió una longitud de 50 m lineales y un intervalo de profundidad de 9 m a 12 m, a lo que se podría atribuir la estabilidad de los datos, ya que para detectar cambios en poco tiempo se requeriría un tamaño muestreal mayor al utilizado por CARICOMP (Green y Smith, 1997), como el adoptado en este estudio (mínimo de 200 m en cada arrecife, en un intervalo de profundidad entre 1.5 m y 29 m y una mayor escala de tiempo).
Al analizar el cambio en la cobertura coralina por especies en cada arrecife, resulta extraño que pocas especies hayan presentado diferencias estadísticamente significativas al comparar sus coberturas en el tiempo, ya que muchas especies exhiben valores diferentes entre pasado y presente, tanto en términos absolutos como relativos. Esto se puede deber a que cada una de las especies pierde muy poca cobertura en cada transecto, y por lo tanto el cambio no es detectado estadísticamente en cada una de las especies, o simplemente el análisis utilizado no es el más apropiado para detectar las diferencias en los datos.
S. radians fue la única especie que redujo su cobertura en el tiempo de forma significativa, tanto en Punta Betín como en Morro, lo cual resulta extraño, ya que esta especie es considerada una de las más tolerantes al estrés en el Caribe, al ser encontrada en lugares con alto grado de perturbación. Según Lirman et al. (2002) no sólo resiste baja salinidad y altas tasas de sedimentación, sino que es altamente resiliente, debido a que las colonias, luego de perder una gran cantidad de tejido, lo recuperan rápidamente. Por su parte, Montastrea cavernosa presentó alta reducción de cobertura relativa en el 2002 respecto al pasado, concordando con lo encontrado por Caro-Zambrano (2003), quien la registró como la especie con mayor pérdida de cobertura en la Bahía de Santa Marta entre 1998 y 2002. Sin embargo, M. cavernosa sigue siendo la especie dominante junto con M. annularis (con coberturas mayores al 20 %) con respecto a la mayoría de especies que se consideran intermedias y raras en el arrecife por sus menores coberturas. Estos resultados concuerdan con los obtenidos por varios autores tales como Solano (1987), quien para bahía Chengue encontró que los escleractíneos más abundantes de cada estación contribuían con más del 70 % de la cobertura total de la misma, y alrededor del 60 % de las especies no superaban el 1 % de cobertura viva, aunque representaban aproximadamente el 30 % del número total de especies. M. annularis, por ejemplo, ha sido registrada como dominante en la Florida (Jaap et al.,1989) y en las Islas Vírgenes (Edmunds, 2002), con coberturas hasta de 40 %.
La marcada dominancia de unas pocas especies, tanto en el pasado como en el presente, se refleja en los valores de diversidad, dominancia y equitatividad, los cuales, a diferencia de los de composición y cobertura, no cambiaron de manera significativa después de una década en ninguno de los arrecifes. Lo anterior indica que aunque la reducción en cobertura ha sido significativa, la pérdida de cobertura ha mantenido las proporciones entre las especies y por lo tanto la uniformidad en los índices. Así se podría decir que los índices de riqueza y diversidad, ampliamente usados en estudios ecológicos, no siempre son buenos indicadores de cambio en las comunidades en el tiempo, por lo menos en arrecifes coralinos, ya que, por ejemplo la riqueza, se puede mantener constante aunque haya cambios en composición, debido a que nuevas especies reemplazan aquellas que se extinguen localmente (McClanahan et al., 2002).
Sin embargo, la mayor reducción de cobertura en las especies dominantes se reflejó, de forma leve y no significante, con el incremento en los valores de diversidad y equitatividad en el tiempo en algunos arrecifes y a la reducción de la dominancia. De esta forma, se incrementa matemáticamente la cobertura relativa de especies intermedias y raras. Lo anterior puede estar apoyando la hipótesis de disturbio intermedio (Connell, 1978), ya que pareciera que las perturbaciones en la última década han afectado principalmente a las especies dominantes. No obstante, el hecho de una mayor equitatividad no se puede interpretar como “positivo” para el arrecife, ya que la pérdida real de cobertura coralina, sin importar la especie, hace a la comunidad menos estructurada aunque se mantenga el número de especies.
La composición coralina cambió en la última década, siendo mayor el porcentaje de cambio dentro de cada arrecife que para toda el área de estudio (total de los cuatro arrecifes), y mayor para especies que para géneros. Los cambios en cada arrecife podrían explicarse a partir de la hipótesis de partición de nichos planteada por Knowlton y Jackson (1994), según la cual muchas especies coralinas tienen alta especificidad de hábitat y nichos, lo cual limita su ámbito de distribución espacial, por lo que su dinámica poblacional, basada en densidad o cobertura, podría responder a pequeños cambios ambientales locales como calidad de agua, haciendo que algunas especies disminuyan su cobertura y otras se extingan localmente. Sin embargo, en este punto hay que tener en cuenta que la ausencia o presencia de varias especies en el muestreo actual con respecto al pasado, podría indicar una reducción en sus coberturas o densidades en el arrecife y no necesariamente su extinción o aparición en el mismo.
El cambio en la estructura coralina, en este caso en términos de cobertura y composición, podría representar, por ende, un cambio en la función como lo han registrado varios autores. Por ejemplo, se ha demostrado que la pérdida de biodiversidad, en términos de número de especies, afecta la estabilidad o persistencia de la comunidad en el tiempo (Stiling, 1999). Adicionalmente, se considera que existe relación directa entre el número de especies y la eficiencia en el uso de nutrientes, productividad (Naeem et al., 1994) y resistencia a perturbaciones por parte del sistema (Tilman et al., 1996). Los arrecifes coralinos se han considerado ecosistemas muy persistentes, pero poco resilientes a las constantes y fuertes fluctuaciones ambientales (McClanahan et al., 2002). Por lo tanto, perturbaciones antropogénicas (directas o indirectas) que causan presiones en grupos funcionales (cambio en composición y abundancia) y en la organización ecológica (como la profundidad máxima de distribución coralina), posiblemente disminuyen la habilidad de la comunidad coralina, de recuperarse y mantenerse en el tiempo (Connell, 1978; McClanahan et al., 2002). Aunque algunas comunidades coralinas aparentemente se hayan recuperado luego de estar sometidas a perturbación y evidencien altas tasas de recolonización, se sabe que es muy difícil que vuelvan a su estado original, ya que pocas veces hay reclutamiento y reemplazamiento de colonias muertas por las mismas especies (evidenciado en Punta de Betín, Santa Marta; J. Rodríguez com. per.), presentándose también cambios en los patrones de zonación, como ocurrió en las Islas del Rosario en Colombia (Cendales et al., 2002). Las poblaciones también manifiestan efectos colaterales, como una menor densidad, disminuyendo las interacciones en el sistema, como ocurrió en los arrecifes de Kaneohe Bay en Hawaii (Castro y Huber, 2000; McClanahan et al., 2002).
Evidentemente, la comunidad coralina del área de Santa Marta – PNNT ha estado sometida a fuerte perturbación, determinada por factores de estrés bióticos, abióticos y antropogénicos (Díaz et al., 2000). El incremento en las últimas décadas de la tasa de sedimentación-resuspensión y de la nutrificación de las aguas por variaciones en el régimen hidrológico del río Magdalena a través de la Ciénaga Grande de Santa Marta, más el aporte de aguas negras en la bahía de Santa Marta, han sido considerados los principales factores de degradación de los arrecifes a partir de los años setenta (Werding y Sánchez, 1988; 1989; Acosta, 1989; Garzón-Ferreira y Cano, 1991).
La alta tasa de mortalidad coralina en el PNN Tayrona, debido a la sedimentación, ha sido evidenciada por Acosta (1989), quien determinó que en invierno la influencia de la sedimentación (con valores de hasta 30.8 mg/cm2/día) superaba la tolerancia de los corales causando en respuesta blanqueamiento localizado. A su vez, dicho autor registró a Punta Betín como el arrecife con menor estructura y mayor deterioro con respecto a los otros arrecifes del área por estar ubicado más cerca de la fuente de perturbación (promedio anual de 3.74 mg/cm2/día en la tasa de resuspensión - sedimentación). Garzón-Ferreira y Cano (1991) propusieron la sedimentación como causante de deterioro, ya que en áreas expuestas y medianamente expuestas al oleaje, los arrecifes en general se han mantenido en buena condición (ej. Bahía Cinto). En otras regiones del Caribe colombiano la pérdida coralina a partir de la década de los ochenta también se ha atribuido principalmente al aumento en la sedimentación local por efectos antropogénicos (Ramírez et al., 1986; 1994; Garzón-Ferreira y Kielman, 1993).
Por su parte, la nutrificación también parece estar influyendo de manera importante en las comunidades coralinas, especialmente en los arrecifes de Punta Betín y Morro Grande, que por ser los más cercanos al emisario submarino del Boquerón, el cual vierte al mar las aguas negras de Santa Marta y el Rodadero, están sometidos al aporte de M.O.P y M.O.D al sistema. Esta influencia de materia orgánica se evidencia con el aumento significativo en la cobertura de esponjas en ambos arrecifes, lo cual seguramente se atribuye a su alta capacidad de utilizar M.O.P y M.O.D como fuente de alimento y a sobrevivir en ambientes con bajos niveles de luz por alta turbidez (Ruppert y Barnes, 1995). Adicionalmente, según Zea (1994), en el área de Santa Marta los arrecifes están cambiando de arrecifes coralinos a arrecifes de esponjas. La aparición de macroalgas en el muestreo de los arrecifes de Morro y Granate podría considerarse otra evidencia del aporte de nutrientes al sistema (McClanahan et al., 2002). Sin embargo, en este punto es importante tener en cuenta la época del año en la que se muestreó (seca menor, caracterizada por una baja temperatura del agua) ya que de acuerdo con Bula-Meyer (1990) las algas en el parque Tayrona son fuertemente estacionales y su abundancia puede deberse a fluctuaciones naturales o a otros factores como la ausencia de herbívoros y no necesariamente a nutrificación del agua (Hughes et al., 1999). Por otra parte, aunque las algas aparecieron en el muestreo en el 2002 después de no haber sido cuantificadas durante 1989 y 1992, su abundancia sigue siendo muy baja con respecto a otros arrecifes del Caribe donde llegan a ser dominantes en los arrecifes (McClanahan et al., 1999; Lirman y Biber, 2000; Chiappone et al., 2001), lo que se puede deber, como se explicó anteriormente, a las bajas temperaturas del agua por fenómenos de surgencia durante épocas secas (Bula-Meyer, 1990; Díaz-Pulido y Garzón-Ferreira, 2002).
Aunque los valores actuales de cobertura coralina (menor cobertura en Punta Betín y mayor en Gayraca) siguen relacionándose con el gradiente de sedimentación y nutrificación registrado en el pasado (Acosta, 1989), la pérdida absoluta de coral vivo durante la última década siguió un patrón inverso (menor en Punta Betín y mayor en Gayraca) para el 2002. Si se tiene en cuenta que Gayraca es, de los cuatro arrecifes estudiados, el más alejado de las fuentes de contaminación ambiental y por lo tanto el que se ha desarrollado bajo unas condiciones de calidad de agua aparentemente mejores, es difícil explicar los factores que han determinado su alta reducción de cobertura coralina. Sin embargo, existen varias posibles explicaciones para este patrón tanto en Gayraca como en los otros tres arrecifes:
1. La menor pérdida de cobertura coralina en los arrecifes del área de Santa Marta se podría atribuir a selección natural, donde las diversas presiones de selección posiblemente han eliminado, en la última década, las especies o genotipos vulnerables y han mantenido aquellas más resistentes a las perturbaciones (i.e. M. cavernosa, Siderastrea siderea; J. Rodríguez com. per.). Contrariamente, en los arrecifes más lejanos a la fuente de perturbación (los del PNNT), hasta ahora se estaría generando presión de selección a los diferentes genotipos, lo que se reflejó en este estudio, en una mayor pérdida relativa de cobertura para las especies menos resistentes, ya que la intensidad de las perturbaciones se ha incrementado en el tiempo (J.M. Díaz com. per.).
2. Parte de la pérdida de cobertura coralina, en la última década, se podría explicar por buceo intensivo, especialmente en el arrecife de Granate, el cual sufrió comparativamente los mayores cambios en la estructura con respecto a lo registrado por Acosta (1989). El buceo es una práctica frecuente en Granate (visitada anualmente por unos 100.000 turistas; Díaz et al., 2000) y ha aumentado considerablemente en la última década, debido a una concesión otorgada por la unidad de Parques Nacionales Naturales para que los operadores de buceo locales concentren sus actividades casi exclusivamente en este arrecife (J.M. Díaz com. per.). Aunque no se ha cuantificado el impacto del buceo sobre los arrecifes en el área de estudio, se sabe que causan disminución en la cobertura coralina en otros arrecifes, como lo calcularon Hawkins y Roberts (1997). Estos autores proponen un umbral de 4000-6000 buzos anuales en un arrecife, donde superada esta cifra, pequeños aumentos en la intensidad de buceo causan un aumento exponencial (de hasta 20 %) en el porcentaje de pérdida de cobertura de coral vivo.
3. La pesca con dinamita es una práctica que se usa desde 1979 de forma extensiva en todas las bahías, aunque al parecer en la última década ha sido un poco controlada. El efecto de la dinamita se ejerce directa y negativamente sobre la cobertura coralina, principalmente de grandes colonias, más propensas a sufrir mortalidad y daño mecánico, como se pudo observar en Granate, dónde había colonias fragmentadas de los géneros Montastrea y Colpophyllia. La bahía de Gayraca también presenta una fuerte influencia pesquera, y Garzón-Ferreira y Cano (1991) detectaron la presencia, casi constante de pescadores con dinamita. Además la bahía posee, desde hace algunos años, carretera de acceso, lo cual puede haber aumentado tanto la pesca como el turismo.
4. El efecto de enfermedades coralinas y la bioerosión al parecer también está disminuyendo la cobertura de las diferentes poblaciones coralinas. Las enfermedades fueron cuantificadas para el Caribe colombiano por Barrios (2000), quien encontró un total de siete que afectan los corales y, aunque éstas son de las condiciones de deterioro las menos frecuentes, son las que presentan mayor mortalidad asociada. Adicionalmente, la autora reportó al PNNT como la región de todo el Caribe colombiano más afectada por mortalidad coralina actual asociada con enfermedades. La bioerosión, por su parte, reduce la cobertura coralina, como por ejemplo M. cavernosa, que fue registrada por Kielman y Hofman (1988) como una de las especies más afectadas por esponjas excavadoras, especialmente las del género Aka, en el arrecife de Morro.
5. Otra opción podría ser el efecto de huracanes (i.e. Mitch 1998 y Lenny 1999). Posiblemente estos contribuyeron a un mayor deterioro en unos arrecifes con respecto a otros, ya que su efecto podría depender de forma directa de la abundancia o cobertura coralina o del grado de exposición de la bahía (S. Zea, com. per.).
El efecto sinérgico de todos los factores de deterioro, tanto naturales (interacciones) como antropogénicos, son los que posiblemente han determinado el estado de las comunidades coralinas en el área de Santa Marta-PNNT, aunque sin seguimiento y experimentación es muy difícil determinar cuáles causas han influido con mayor intensidad y en qué momento. Nuevos estudios deben enfocarse en la posibilidad de recuperación de este ecosistema de forma natural a través del reclutamiento coralino, y nuevas medidas de manejo deben tomarse en el área con el fin de mitigar los factores de perturbación ya conocidos, ya que el deterioro de las comunidades coralinas en estructura y posiblemente en función implica pérdida de bienes y servicios para las comunidades costeras.
Como conclusión, podemos indicar que las comunidades coralinas de los arrecifes de Punta Betín, Morro Grande, Granate y Gayraca en la región de Santa Marta–PNNT han cambiado significativamente en la última década hacia un estado de mayor deterioro. Este cambio se evidencia, principalmente, en la composición de especies y en la pérdida de cobertura coralina que han sufrido todos los arrecifes. Los datos obtenidos en este estudio así como los registros de otros arrecifes en el Caribe, confirman la dirección y la magnitud de cambio, indicando que estas tasas de deterioro son altas e insostenibles en escalas ecológicas. Por lo tanto, de continuar esta tasa de pérdida coralina, en pocas décadas o siglos hablaremos no sólo de extinción local de especies, sino de arrecifes. Por lo tanto, las fuentes de perturbación identificadas en los diferentes estudios deben mitigarse (hasta donde sea posible) a través de planes de manejo a corto, mediano y largo plazo; y de esta forma permitir a los arrecifes recuperarse y mantenerse en el tiempo.
AGRADECIMIENTOS
Al INVEMAR por el apoyo económico y logístico prestado para realizar este proyecto.
BIBLIOGRAFÍA
1. Acosta, A. 1989. Composición y estructura de la comunidad de corales hermatípicos en tres zonas representativas de la región de Santa Marta (mar Caribe colombiano). Tesis de grado, Departamento de Biología, Universidad Nacional de Colombia, Bogotá, 242p. [ Links ]
2. Acosta, A. y S. Zea. 1997. Sexual reproduction of the coral Montastrea cavernosa (Scleractinia, Faviidae) in the Santa Marta area, Caribbean coast of Colombia. Marine Biology, 128:141-147. [ Links ]
3. Antonius, A. 1972. Occurrence and distribution of stony corals (Anthozoa and Hydrozoa) in the vicinity of Santa Marta, Colombia. Mitt. Inst. Colombo-Alemán Invest. Cient., 6:89-103. [ Links ]
4. Barrios, L.M. 2000. Evaluación de enfermedades en corales pétreos del Caribe colombiano. Tesis de Maestría, Universidad Nacional de Colombia, Bogotá, 155p. [ Links ]
5. Brattström, H. 1980. Rocky-shore zonation in the Santa Marta area, Colombia. Sarcia, 65:163-226. [ Links ]
6. Brüggemann, J., M. Hernández, E. Rodríguez, J. Soler y R. Tapper. 2002. Biodiversity and tourism in the framework on the Convention of Biological Diversity: The case of Tayrona National Park, Colombia. Report of the Workshop at Cañaveral, Tayrona National Park, 25 – 30 November 2001, German Federal Agency for Nature Conservation, 41p. [ Links ]
7. Bula-Meyer, G. 1990. Altas temperaturas estacionales del agua como condición disturbadora de las macroalgas del Parque Nacional Natural Tayrona, Caribe Colombiano: una hipótesis. An. Inst. Invest. Mar. Punta de Betín, 19-20:9-21. [ Links ]
8. Caro-Zambrano, C. 2003. Patrones de variación temporal y espacial de los arrecifes coralinos colombianos monitoreados en el período 1998-2001. Tesis de grado, Universidad Jorge Tadeo Lozano, Bogotá, 182p. [ Links ]
9. Castro, P. y M.E. Huber. 2000. Marine Biology. 3rd Edition. McGraw Hill Higher Education, USA, 444p. [ Links ]
10. Cendales, M.H., S. Zea y J.M., Díaz. 2002. Geomorfología y unidades ecológicas del complejo de arrecifes de las Islas del Rosario y Barú (Mar Caribe, Colombia). Rev. Acad. Colomb. Cienc., 26(101)497-510. [ Links ]
11. Chiappone, M., K. Sullivan, G. Bustamante y J. Tschirky. 2001. A rapid assessment of coral reef community structure and diversity patterns at naval station Guantánamo Bay, Cuba. Bull. Mar. Sci., 69:373-394. [ Links ]
12. Connell, J. 1978. Diversity in tropical rain forest and coral reefs. Science, 119:1302-1310. [ Links ]
13. _____. 1997. Disturbance and recovery of coral assemblages. Proceedings of the 8th International Coral Reef Symposium. Panamá, junio 24-29, 1: 9-22. [ Links ]
14. De Jongh, B.O. 1990. A comparison between quantitative taxonomic and physiognomic survey methods of hard substrate communities of the Santa Marta area, NE Colombia. Tesis de Grado, Institute of Taxonomic Zoology University of Amsterdam, 5-30pp. [ Links ]
15. Díaz, J.M., J. Garzón-Ferreira y S. Zea. 1995. Los arrecifes coralinos de la Isla de San Andrés, Colombia: estado actual y perspectivas para su conservación. Academia Colombiana de Ciencias Exactas, Físicas y Naturales, Colección Jorge Álvarez Lleras, Bogotá, 7:1-150. [ Links ]
16. Díaz, J.M., L.M. Barrios, M.H. Cendales, J. Garzón-Ferreira, J. Geister, M. López-Victoria, G. H. Ospina, F. Parra-Velandia, J. Pinzón, B. Vargas-Angel, F.A. Zapata y S. Zea. 2000. Áreas Coralinas de Colombia. Instituto de Investigaciones Marinas y Costeras “José Benito Vives de Andreis” INVEMAR, Santa Marta, Serie de Publicaciones especiales No. 5., 17-28, 130-136, 155-167pp. [ Links ]
17. Díaz-Pulido, G. y J. Garzón-Ferreira. 2002. Seasonality in algal assemblages on upwelling-influenced coral reefs in the Colombian Caribbean. Bot. Mar., 45:284-292. [ Links ]
18. Edmunds, P. 2002. Long-term dynamics of coral reefs in St. John, US Virgin Islands. Coral Reefs, 21: 357-367. [ Links ]
19. Erhardt, H. y B. Werding. 1975. Los corales (Anthozoa e Hidrozoa) de la Bahía de Santa Marta. Boletín Museo del Mar, 7: 3-50. [ Links ]
20. Garzón-Ferreira, J. 2001. Informe del estado de los ambientes marinos y costeros en Colombia: 2000. Serie documentos generales No. 3, Santa Marta, Colombia. 25-36pp. [ Links ]
21. Garzón-Ferreira, J. y M. Cano. 1991. Tipos, distribución, extensión y estado de conservación de los ecosistemas marinos costeros del Parque Nacional Natural Tayrona. Manuscr. VII Concur. Nal. Ecol. FEN/INVEMAR, Bogotá, Santa Marta. 82p. [ Links ]
22. Garzón Ferreira, J. y M. Kielman. 1993. Extensive mortality of corals in the Colombian Caribbean during the last two decades. Proc. Colloquium on Global Aspects of Coral Reefs: Health, Hazards and History. RSMAS, Univ. Miami, Miami, 247-253pp. [ Links ]
23. Ginsburg, R. (Com). 1994. Proc. Colloquium on Global Aspects of Coral Reefs: Health, Hazards and History. RSMAS, Univ. Miami, Miami, 420p. [ Links ]
24. Global Coral Reef Monitoring Network (GCRMN). 2000. Status of Coral Reefs of the World: 2000. C. Wilkinson (Ed). Australian Institute of Marine Science, Cape Ferguson, Queensland and Dampier, Western Australia, 363p. [ Links ]
25. Green, R.H. y S.R. Smith. 1997. Sample program design and environmental impact assessment on coral reefs. Proc. 8th Int. Coral Reef Symp. Panamá., 2:1459-1464. [ Links ]
26. Hallock, P., Müller-Karger, F.E. y J.C. Halas. 1996. Oceanography. K. Gordon Pirie (Ed). Contemporary readings in Ocean Sciences. Third Edition, Oxford Univ. Press., 233-249pp. [ Links ]
27. Hawkins, J.P. y C.M. Roberts. 1997. Estimating the carrying capacity of coral reefs for SCUBA diving. Proc. 8th Int. Coral Reef Symp. Panamá., 2: 1923-1926. [ Links ]
28. Hughes, T. 1994. Catastrophes, phase shifts, and large scale degradations of a Caribbean coral reef. Science, 265:1547-1551. [ Links ]
29. Hughes, T.P., A.M. Szmant, R.S. Steneck, R.C. Carpenter y S. Miller. 1999. Algal blooms on coral reefs: what are the causes? Limnol. Oceanogr., 44:1583-1586. [ Links ]
30. Hughes, T. y J. Tanner. 2000. Recruitment failure, life histories, and long-term decline of Caribbean corals. Ecology, 81: 2250-2263. [ Links ]
31. Jaap, W.C., W.G. Lyons, P.H. Dustan y J.C. Halas. 1989. Stony Coral (Scleractinia and Milleporina) community structure at Bird Key Reef, Ft. National Monument, Dry Tortugas, Florida. Florida Marine Research Publications, No. 46, 1-23 pp. [ Links ]
32. Kielman, M. y C. Hofman. 1988. A quantitative study on the excavating sponges of the Santa Marta area, Colombia. Institute of Taxonomic Zoology. Tesis de Grado. University of Amsterdam. Amsterdam, 20-39pp. [ Links ]
33. Knowlton, N. y J.B.C. Jackson. 1994. New taxonomy and niche portioning on coral reefs: jack of all trades or master of some? Trends Ecol. Evol., 9:7-9. [ Links ]
34. Lirman, D. y P. Biber. 2000. Seasonal dynamics of macroalgal communities of the northern Florida Reef Tract. Bot. Mar., 43:305-314. [ Links ]
35. Lirman, D., D. Manzello y S. Maciá. 2002. Back from the dead: The resilience of Siderastrea radians to severe stress. Coral Reefs, 21:291-292. [ Links ]
36. Loya, Y. 1972. Community structure and species diversity of hermatipic corals at Elliat, Red Sea. Mar. Biol., 23:100-123. [ Links ]
37. McClanahan, T., R.B. Aronson, W.F. Precht y N.A. Muthiga. 1999. Fleshy algae dominate remote coral reefs of Belize. Coral Reefs, 18:61-62. [ Links ]
38. McClanahan, T., N, Polunin y T. Done. 2002. Ecological states and resilience of the coral reefs. Conserv. Ecol., 6: 18. [ Links ]
39. Magurran, A. 1998. Ecological Diversity and its Measurement. Princenton Univ. Press, Princenton, NJ, USA, 179 pp. [ Links ]
40. Naeem, S., L.J. Thompson, S.P. Lawler, J.H. Lowton y R.M. Woodfin. 1994. Declining biodiversity can alter the performance of ecosystems. Nature, 368:734-737. [ Links ]
41. Pandolfi, J.M. 1999. Response of Pleistocene coral reefs to environmental change over long temporal scales. Amer. Zool., 39:113-130. [ Links ]
42. _____. 2002. Coral community dynamics at multiple scales. Coral Reefs, 21:13-23. [ Links ]
43. Pandolfi, J.M. y J.B.C. Jackson. 1997. The maintenance of diversity on coral reefs: Examples from the fossil record. Proc. 8th Int. Coral Reef Symp. Panamá, 1:397-404. [ Links ]
44. Porter, J.W. y K.G. Porter. 2002. Detection of coral reef change by the Florida Keys coral reef monitoring project. The Everglades, Florida Bay and coral reefs of the Florida Keys. An ecosystems sourcebook. CRC Press, Miami, 755-769 pp. [ Links ]
45. Porter, J.W., P.H. Dustan, W.C. Jaap, K.L. Patterson, V. Kosmynin, O.W. Meier, M.E. Patterson y M. Parsons. 2001. Patterns of spread of coral disease in the Florida Keys. Hydrobiologia, 460: 1-24. [ Links ]
46. Prahl, H. y H. Erhardt. 1985. Colombia: corales y arrecifes coralinos. Fondo para la protección del medio ambiente “José Celestino Mutis” – FEN- Bogotá, Colombia, 231-245 pp. [ Links ]
47. Ramírez, A., I. De Ramírez y J. Correal. 1986. Ecología descriptiva de las llanuras madreporarias del Parque Nacional Submarino Los Corales del Rosario (Mar Caribe), Colombia. Fundación Universidad de Bogotá Jorge Tadeo Lozano. Bogotá, Colombia, 11-23 pp. [ Links ]
48. Ramírez, A., D. Miranda y G. Viña. 1994. Estructura arrecifal del archipiélago de San Bernardo (Mar Caribe, Colombia). Estudio de línea base. Trianea, 5:189-219. [ Links ]
49. Rodríguez-Ramírez, A. y J. Garzón-Ferreira. 2003. Monitoreo de arrecifes coralinos, pastos marinos y manglares en la Bahía de Chengue (Caribe colombiano):1993-1999. INVEMAR, Serie de Publicaciones especiales No. 8, Santa Marta, 170pp. [ Links ]
50. Ruppert, E.E. y R.D. Barnes. 1995. Zoología de los invertebrados. 6a Edición. McGraw Hill, Interamericana, Mexico, 1114 pp. [ Links ]
51. Solano, O.D. 1987. Estructura y diversidad de la comunidad de corales hermatípicos en la Bahía de Chengue. Tesis de grado. Universidad Nacional. Bogotá, 111pp. [ Links ]
52. Stiling, P.D. 1999. Ecology. Theories and Applications. 3rd Edition. Prentice Hall, Upper Saddle River, New Jersey. USA. 638pp. [ Links ]
53. Tilman, D., D.Wedin y J. Knops. 1996. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature, 379:718-720. [ Links ]
54. Ugland, K.I., J.S. Gray y K.E. Ellingsen. 2003. The species-accumulation curve and estimation of species richness. J. Animal Ecology, 72: 888-897. [ Links ]
55. Werding, B. y H. Sánchez. 1988. Deterioro observado en las formaciones coralinas de la Bahía de Santa Marta, Colombia. An. Inst. Inv. Mar. Punta de Betín, 18: 9-16. [ Links ]
56. Werding, B. y H. Sánchez. 1989. The coral formations and their distributional pattern along a wave exposure gradient in the area of Santa Marta, Colombia. Medio Ambiente, 10(2):61-68. [ Links ]
57. Zea, S. y F. Duque-Tobón. 1989. Bleaching of reef organisms in the Santa Marta Region, Colombia: 1989 Caribbean-wide event. Trianea, 3:37-51. [ Links ]
58. Zea, S. 1993. Cover of sponges and other sessile organisms in rocky and coral reef habitats of Santa Marta, Colombian Caribbean Sea. Carib. J. Science, 29:75-88. [ Links ]
59. _____. 1994. Patterns of coral and sponge abundance in stressed coral reefs at Santa Marta, Colombian Caribbean. En: R.W.M. van Soest, T.M.G. van Kempen y J.-C. Braekman (eds.), Sponges in time and space: Biology, Chemistry, Paleontology. Balkema, Rotterdam, 257-264pp. [ Links ]
