










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.35 no.1 Santa Marta Jan./Dec. 2006
CRECIMIENTO DEL ABANICO DE MAR GORGONIA VENTALINA (LINNAEUS, 1758) (CNIDARIA: GORGONIIDAE) EN EL ÁREA DE SANTA MARTA, CARIBE COLOMBIANO
GROWTH OF SEA FAN GORGONIA VENTALINA (LINNAEUS, 1758) (CNIDARIA: GORGONIIDAE) IN THE SANTA MARTA AREA, COLOMBIAN CARIBBEAN
Nelson Manrique-Rodríguez1, Sonia Bejarano-Chavarro2 y Jaime Garzón-Ferreira3
1Laboratorio de Biología Molecular Marina BIOMMAR, Universidad de los Andes, Bogota, Colombia. E-mail: na.manrique47@uniandes.edu.co (NMR)
2Marine Spatial Ecology Lab MSEL. University of Exeter, UK. E-mail: sb277@exeter.ac.uk
3Instituto de Investigaciones Marinas y Costeras (INVEMAR), Cerro Punta Betín, Santa Marta, Colombia. E-mail: jgarzon@invemar.org.co
RESUMEN
Como contribución al conocimiento de la biología del abanico de mar Gorgonia ventalina y para aportar información que sirva de apoyo en el diseño de estrategias de manejo y conservación de esta especie amenazada, se desarrolló una investigación sobre su tasa de crecimiento en varias formaciones coralinas del área de Santa Marta (incluyendo el Parque Nacional Natural Tayrona) en el Caribe colombiano. Se analizaron mediciones manuales de 35 colonias y fotográficas en 11 de esas 35 colonias al inicio y al final del estudio al cabo de ocho meses. La tasa media anual con los dos métodos fue alta y osciló entre 7.6 y 8.1 cm/año de altura y 6.9 y 8.3 cm/año de ancho respectivamente y con un incremento en área de 246.1 cm2/año, esta elevada tasa sugiere que el área de estudio posee las características ideales para el óptimo desarrollo de la especie. Sin embargo, el crecimiento de estas poblaciones puede estar limitado por cambios ambientales, biológicos y antrópicos.
PALABRAS CLAVE: Crecimiento, Abanicos de mar, Gorgonia ventalina, Santa Marta, Caribe colombiano.
ABSTRACT
As a contribution to the biological knowledge about the sea fan Gorgonia ventalina and the use of this information as a tool to generate conservation and management strategies for this threatened species, a study about its annual growth rate, in different reef areas in Santa Marta and the National Natural Park Tayrona in the Colombian Caribbean took place. Manual measurements of 35 colonies were analyzed and images of 11 colonies were recorded in the beginning and in the end of the field stage. The annual growth rate varies among colonies and areas where measurements were taken. The average annual growth rate obtained with both of the methods was high and oscillated between 7.6 and 8.1 cm/year in height and between 6.9 and 8.3 cm/year wide. The total area of the colonies increased 246.1 cm2/year. These high rates suggest that the study area has the ideal features for the optimal development of the species. Eventhough, these features may be limited by environmental, biological and anthropic changes.
KEY WORDS: Growth, Sea fans, Gorgonia ventalina, Santa Marta, Colombian Caribbean.
INTRODUCCIÓN
Los gorgonáceos son habitantes conspicuos y de gran importancia ecológica en los arrecifes coralinos del mar Caribe (Bayer, 1961). Entre ellos, el abanico de mar Gorgonia ventalina (Linnaeus, 1758) era el más notorio e importante en las zonas someras (Cairns, 1977; Lasker y Coffroth, 1983; Botero, 1987) hasta finales de los años ochenta cuando sufrió una serie de mortandades masivas en diversas localidades (Smith et al., 1996) como Costa Rica, Panamá (Guzmán y Cortés, 1984), Trinidad (Laydoo, 1983) y Jamaica (Geiser et al., 1998). Se estima que las poblaciones del Caribe colombiano fueron afectadas entre 1988 y 1994, llegando a desaparecer casi por completo en varias de las principales áreas coralinas como las Islas del Rosario, Santa Marta y Parque Nacional Natural Tayrona (Garzón-Ferreira y Zea, 1992), San Andrés (Díaz et al.,1995) y Cayos de Albuquerque y Courtown (Sánchez et al., 1997). Tales fueron la magnitud y el alcance de estos eventos que actualmente Gorgonia ventalina es considerada especie amenazada a nivel regional y se ha incluido localmente entre las especies vulnerables a enfrentar un moderado riesgo de extinción o deterioro poblacional a mediano plazo (Ardila et al., 2002).
Aspectos básicos de la biología y ecología de G. ventalina han sido descritos por Wainwright y Dillon (1969), Grigg (1972), Alcolado et al. (1980) y Yoshioka y Yoshioka (1987). Se han estudiado otros aspectos como el origen, distribución y posibles causas de la mortandad masiva (Nagelkerken et al., 1997a y b) y la dinámica de contagio de la Aspergillosis (Harvell et al., 2004). Sin embargo, en pocos casos se ha registrado su tasa de crecimiento (i.e. Birkeland, 1974;Yoshioka y Yoshioka, 1991). Aunque en Colombia, se han llevado a cabo evaluaciones de la estructura de la comunidad de gorgonáceos en varias localidades (Botero, 1987; Sánchez, 1995 y 1999; Sánchez et al., 1997),no existe información reciente sobre el estado y el desarrollo de las poblaciones del abanico de mar 15 años después de la mortalidad masiva. Son pocas las estimaciones de la tasa de crecimiento a pesar de saberse que este es uno de los atributos claves en la persistencia de las especies y en el desarrollo de las poblaciones.
Teniendo en cuenta la problemática de los abanicos de mar G. ventalina en el Caribe y como un aporte al vacío de información descrito, se estimó la tasa de crecimiento anual de abanicos de mar de diferentes tallas. Este trabajo formó parte de la evaluación del estado actual de las poblaciones de Gorgonia ventalina en el área de Santa Marta y el Parque Nacional Natural Tayrona durante el 2003 dentro de las actividades del Sistema Nacional de Monitoreo de Arrecifes Coralinos en Colombia (SIMAC). El objetivo de esta evaluación fue proporcionar información científica útil para generar alternativas encaminadas al manejo y conservación de esta especie, por lo cuál esta evaluación se enmarcó dentro del programa de Biodiversidad y Ecosistemas Marinos BEM en la línea de Biología y Estrategias de Conservación del INVEMAR.
ÁREA DE ESTUDIO
El área de Santa Marta y el Parque Nacional Natural Tayrona están ubicados sobre la costa del departamento del Magdalena (Figura 1). Su topografía está determinada por la entrada directa al mar de las estribaciones de la Sierra Nevada de Santa Marta. Sus costas generalmente rocosas y diversos acantilados, bahías, ensenadas, puntas e islotes generan un alto grado de heterogeneidad ambiental (Garzón-Ferreira y Cano, 1991). Estas características geomorfológicas, el tipo de sustrato y el espesor del andamiaje coralino, han dado origen a dos tipos de formaciones coralinas básicas: aquellas formadas por colonización parcial de biota hermatípica sobre bloques de rocas y aquellas edificadas sobre un sustrato rocoso más estable, que han alcanzado una considerable cobertura (Díaz et al., 2000).
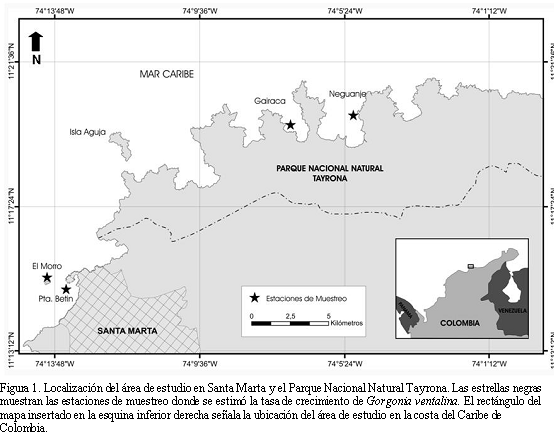
En las zonas más protegidas de las bahías como en Punta Betín y Neguanje las formaciones coralinas son más complejas y diversas. Inclusive, hacia las zonas en donde el relieve escarpado lo permite, se desarrollan en un sustrato muy inclinado arrecifes de parche que actúan como rompeolas y dan protección a la costa contra la erosión (Díaz et al., 2000). En el interior de las zonas expuestas de las bahías y ensenadas del área como en Gairaca y el Morro, el sustrato duro (rocoso y/o coralino) es menos abundante en los primeros 15 m de profundidad. En el islote Morro Grande y en Isla Aguja e islotes vecinos, el fondo rocoso-coralino se extiende hasta los 40-45 m de profundidad y la pendiente del fondo es más pronunciada que en el sublitoral de las bahías y ensenadas (Botero, 1990).
MATERIALES Y MÉTODOS
La tasa de crecimiento de 35 colonias sanas y distribuidas en diferentes rangos de tallas (R1= 18 colonias de 1 a 20 cm; R2= 9 colonias de 21 a 40 cm y R3= 8 colonias mayores de 41 cm) fue evaluada manualmente entre marzo y diciembre de 2003, en cuatro estaciones someras (2 a 15 m) ubicadas en los costados expuestos al oleaje del Morro y Gairaca y en las zonas protegidas de Punta Betín y Neguanje. Para tal efecto, cada colonia fue numerada con un rótulo de cinta plástica (Rotex 3M®) atado con alambre dulce a su parte basal. La altura y ancho máximos fueron medidos con una cinta métrica flexible (precisión, 0.1 cm) al principio y al final del periodo de estudio (Figura 2a y b). Inicialmente se marcaron en total 60 colonias de las cuales 14 se perdieron y 11 fueron afectadas por diferentes condiciones de deterioro durante los ocho meses, presentando tasas de crecimiento nulas (menores o iguales a cero). Estas colonias no fueron contempladas dentro de los análisis, para minimizar el error (Birkeland, 1974; Lasker et al., 2003).
Complementariamente, de las 35 colonias monitoreadas con el método manual, se seleccionaron 11 para hacerles un seguimiento fotográfico con una cámara digital (Sony® Cybershot 4.1). Todas las fotografías se tomaron utilizando una tabla acrílica cuadriculada (divisiones de 1 x 1 cm) como referencia de tamaño (Figura 2c y d), que permitió calcular el incremento total en longitud y área durante el monitoreo. Por otra parte, para cuantificar el crecimiento de las colonias que tuvieron seguimiento fotográfico, se editaron las imágenes en el programa Adobe Photoshop 7® ajustando su cuadricula a otra sobrepuesta a la imagen. Posteriormente se calculó el incremento en altura, ancho y área de cada colonia empleando el programa Scionimage® (Scion Corporation Frederick, MD) el cual estima la cantidad de píxeles contenidos en un segmento o área de referencia de la cuadricula en la fotografía (1 cm o 1 cm2) y, basándose en esta estimación, determina la longitud de un segmento mayor (e.g. altura o ancho) o el área de superficie de la colonia.

Un año después de la primera medición (marzo 2004) se realizó una tercera visita a las cuatro estaciones en la cual fue posible reubicar 12 de las colonias marcadas inicialmente en dos de las estaciones. No obstante, estos datos se compararon para ver alguna variación con las tasas anuales estimadas a partir de la información recopilada durante ocho meses. La tasa de crecimiento de cada colonia estimada con los dos métodos durante el periodo de monitoreo se calculó restando su longitud final de su longitud inicial y su área inicial de su área final. Suponiendo que el crecimiento es constante a través del tiempo, se estimaron a partir de estos valores la tasa media mensual y anual de cada colonia, y de cada estación. La tasa media de crecimiento estimada con los dos métodos en el área de estudio se determinó a partir de los promedios de sus cuatro estaciones, obtenidas a su vez a partir de los valores calculados para cada colonia. Además se determinó el valor promedio para las estaciones de costados expuestos y protegidos del oleaje con el objeto de evaluar diferencias en relación con la dinámica del agua, agrupando los valores de las estaciones correspondientes. Se estimó otro valor promedio para examinar la variabilidad de las tasas entre colonias de diferentes tallas, agrupadas en tres clases (R1= 18 colonias de 1 a 20 cm, R2= 9 colonias de 21 a 40 cm y R3= 8 colonias mayores de 41 cm).
A pesar de los problemas de replicabilidad entre los tres factores a estudiar (localidad, exposición al oleaje, y talla) generados por la pérdida de varias colonias, se ajustó el mejor modelo estadístico que generara el menor ruido en la información. Como los datos no se ajustaron a una distribución normal (prueba de Shapiro-Wilks) aún después de ser transformados (log10), para evaluar si existe variabilidad en la tasa anual de crecimiento entre las estaciones del área de estudio y entre tasas de las diferentes clases de talla establecidos, se llevó a cabo el análisis univariado de varianza de Kruskal-Wallis (Sokal y Rohlf, 1979). A su vez, para determinar si hubo diferencias en la tasa anual de crecimiento con respecto a la exposición al oleaje y los métodos de muestreo, se empleó la prueba pareada no paramétrica de Mann Whitney-Wilcoxon (Sokal y Rohlf, 1979). Estos análisis estadísticos se desarrollaron con el programa STATGRAPHIC Centurión V®.
RESULTADOS
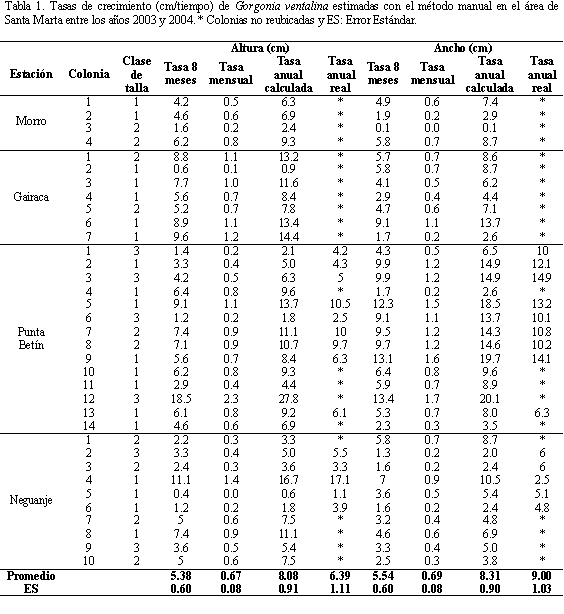
Las estimaciones con el método manual muestran que las colonias de Gorgonia ventalina crecen mensualmente como promedio 0.6±0.1 cm de altura y 0.7±0.1 cm de ancho. Por lo tanto, suponiendo que el crecimiento sea constante, las poblaciones pueden crecer 8.1±0.9 cm de altura y 8.3±0.9 cm de ancho en un año (Tabla 1). En la figura 3, se presenta la tasa media de crecimiento anual calculada en cada estación con el método manual. Nótese que el incremento en altura presentó valores máximos en Gairaca (9.9±1.7 y 9±1.7 cm/año, respectivamente) y valores mínimos en el Morro y Neguanje (6.2±1.5 y 6.2±1.6 cm/año respectivamente). La tasa de incremento calculada en ancho presentó valores máximos en Punta Betín (12.8±1.5 cm/año) y mínimos en el Morro (4.7± 1.9 cm/año). A su vez, al revisar los promedios estimados con la información recopilada durante ocho meses y la real registrada durante un año, se observa que los valores son similares. Sin embargo, esta comparación muestra una diferencia cercana a los dos centímetros que pudo ser generada por las colonias que no fueron reencontradas (14 colonias).
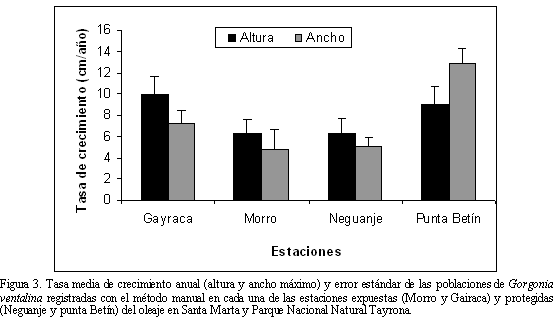
Por otro lado, a pesar de los problemas de replicabilidad en el estudio, la prueba de Kruskal-Wallis reveló que no se presentaron diferencias estadísticamente significativas en las tasas de crecimiento en altura entre las diferentes estaciones (P= 0.147). En contraste, se evidenciaron diferencias significativas en las tasas de crecimiento en ancho (P= 0.009), mostrando que en la estación de Punta Betín el incremento fue mayor que en las demás estaciones. Por otro lado, la prueba de Mann Whitney-Wilcoxon indicó que no existen diferencias entre la exposición al oleaje y la tasa de crecimiento en altura (P= 0.310) ni en ancho (P= 0.249). Así mismo, la variabilidad en las tasas de crecimiento entre las diferentes clases de talla con la prueba de Kruskal-Wallis no presentó diferencias significativas en altura (P= 0.426) ni en ancho (P= 0.391).
Las 11 colonias monitoreadas con el método fotográfico evidencian una tasa de crecimiento promedio mensual de 0.6±0.1 cm de altura y de 0.7±0.1 cm de ancho, así como un incremento anual de 7.6±1.0 cm de altura y de 6.9±1.2 cm de ancho. Con el método fotográfico, sobresalió la tasa de crecimiento más alta en la estación de Punta Betín (altura: 9.6±3.3 cm y ancho: 10.4±5.1 cm) y la tasa más baja se registró en Gairaca (altura: 6.4±2.3 cm y ancho: 2.7±2.0 cm) (Tabla 2). Finalmente, la variabilidad en las tasas de crecimiento entre los dos métodos, analizada con la prueba de Mann Whitney-Wilcoxon no reveló contrastes significativos en altura (P= 0.947) y ancho (P= 0.212). Por otro lado, con el método fotográfico se registró un incremento promedio anual en área de 246.1±80.6 cm2 siendo también la estación de Punta Betín la que presentó el mayor incremento (417.6±205.4 cm2) y la estación de Neguanje el incremento más bajo (131.6±53.8 cm2) (Tabla 2).
DISCUSIÓN Y CONCLUSIONES
Luego de monitorear la tasa de crecimiento de Gorgonia ventalina en cuatro formaciones coralinas del área de Santa Marta, se registró que ésta fue mayor (8.1 y 8.3 cm/año de altura y ancho, respectivamente) a la estimada por Yoshioka y Yoshioka (1991). Estos autores estimaron una tasa de 2.1 cm/año para G. ventalina empleando el método manual en dos arrecifes al sur oeste de Puerto Rico. Igualmente, esta tasa de crecimiento fue superior a la registrada por Birkeland (1974) en dos zonas expuestas al oleaje en Panamá. Este autor estimó una tasa de 4.1 cm/año con el método manual. La elevada tasa de crecimiento en el área de Santa Marta puede estar determinada por la disponibilidad de alimento en partículas suspendidas y microorganismos del plancton, proporcionados por la escorrentía y aguas residuales. Esto, sumado a la orientación de las corrientes que pueden dispersar toda esta materia orgánica en el área (Ramírez, 1990) y favorecer el acceso de los abanicos al alimento disponible. De esta manera se sugiere que las colonias que tienen en Santa Marta una mayor disponibilidad de alimento, un metabolismo más elevado y por ende un mayor crecimiento que en otras áreas, con una menor influencia de la escorrentía.
A pesar de los problemas de replicabilidad entre los tres factores, se especula que la variabilidad de las tasas de crecimiento entre las colonias puede estar relacionada con su estado de salud. Aunque se escogieron colonias sanas para el seguimiento, varias sufrieron diferentes condiciones de deterioro. Además, se infiere que estas colonias destinaron gran parte de su energía metabólica en la regeneración de tejido y por consiguiente su crecimiento fue limitado. Esta afirmación se sustenta con las tasas de crecimiento nulas y negativas en colonias afectadas por los agentes de deterioro en las estaciones de Neguanje y Gairaca. Por otro lado, a pesar de no evidenciar diferencias estadísticamente significativas en las tasas de crecimiento de altura entre las estaciones, los valores más altos se registraron en Gairaca, donde las colonias monitoreadas fueron predominantemente jóvenes y con una baja incidencia de condiciones de deterioro (Bejarano et al., 2005).
Las tasas de crecimiento en ancho, que fueron significativamente mayores en las colonias de Punta Betín pueden estar relacionadas con la influencia del río Manzanares que desemboca en la Bahía de Santa Marta, el emisario submarino que descarga sus aguas residuales en un punto muy cercano a la estación y las actividades de la zona portuaria, los cuales generan una alta turbidez (Ramírez, 1990). Se sugiere que allí las colonias zooxanteladas de Gorgonia ventalina tienden a crecer más hacia lo ancho para tener una mayor área de captura de rayos solares.
Fenómeno conocido como plasticidad fenotípica que ha sido registrado en varios organismos clónales (Willis, 1985; Ayre y Willis, 1988; Anthony, 1999). Por otro lado, tal como lo registró Birkeland (1974) en Panamá, en el presente estudio no se encontraron diferencias significativas en la tasa de crecimiento con respecto a la influencia del oleaje. Esto puede ser sustentado por la flexibilidad característica de los abanicos y la orientación en ángulo recto de las colonias frente al oleaje, lo que les permite mitigar la dinámica de este sin alterar su desarrollo, como lo han descrito Wainwright y Dillon (1969) y Alcolado et al. (1980). La razón de no evidenciar diferencias entre las tasas de crecimiento en las diferentes clases de tallas, radica en que el tamaño de las colonias es controlado por el balance entre la disponibilidad de alimento y la tasa metabólica (Rinkevich, 2000; Lasker et al., 2003) procesos que han sido modelados por Sebens (1987) y Kim y Lasker (1998).
El método fotográfico digital resultó ser una herramienta precisa, ya que además de estimar medidas lineales (altura y ancho), permite estimar el incremento en área, que es el indicador de crecimiento más riguroso (Juan Armando Sánchez. Universidad de los Andes. Bogotá, Colombia. 2004. Com. Pers.). A pesar de que las estimaciones con los dos métodos no presentaron diferencias significativas, las tasas de crecimiento en longitud calculadas con el método fotográfico fueron muy similares en comparación a las obtenidas mediante el seguimiento manual. La estación de Punta Betín mostró a través de este método un mayor incremento en ancho, mientras que las estaciones de Neguanje y Gairaca exhibieron mayor crecimiento en altura. Nunca antes había sido registrado el crecimiento en área para Gorgonia ventalina, lo que permite evidenciar un mayor crecimiento de las colonias en Punta Betín (417.6 cm2), que en las otras estaciones. La explicación de estos resultados puede estar dada por las mismas razones ambientales y ecológicas que se discutieron para el método manual.
En resumen se considera que el área de Santa Marta y el Parque Nacional Natural Tayrona presentan las características ambientales favorables para el buen crecimiento de los abanicos de mar. No obstante, estos atributos están siendo actualmente condicionados por cambios en la temperatura, turbidez, incidencia de condiciones de deterioro y la influencia antrópica en el área que pueden repercutir en su historia de vida.
RECOMENDACIONES
Se recomienda diseñar y ejecutar rigurosamente estrategias puntuales de manejo y conservación, que vigilen y protejan a los abanicos de mar Gorgonia ventalina de Santa Marta y el parque Tayrona, que han venido recuperándose paulatinamente especialmente en el sector de Gairaca (Manrique-Rodríguez, 2004). Igualmente es importante seguir investigando la biología de esta especie, con estudios acerca de su diversidad genética, reproducción, reclutamiento, regeneración, restauración, efectos de las principales condiciones de deterioro y realizar programas de monitoreo permanente en el área y en otras localidades del Caribe colombiano donde se vio seriamente amenazada esta especie.
AGRADECIMIENTOS
Los autores quieren expresar sus más sinceros agradecimientos al Instituto de Investigaciones Marinas y Costeras INVEMAR por el apoyo prestado a lo largo de ésta investigación bajo el marco del Sistema Nacional de Monitoreo de Arrecifes Coralinos en Colombia SIMAC y el programa Caribbean Coastal Marine Productivity CARICOMP. Igualmente a Juan Manuel Díaz, Camilo García, Sven Zea, Juan Armando Sánchez y Adolfo Sanjuán. Los estudiantes del INVEMAR: Mónica Sepúlveda, Edwin Gómez, Isabel C. Chacón G. y Katherine Bernal participaron activamente en la fase de campo. Esta publicación forma parte del trabajo de grado de Nelson Manrique para optar por el título de Biólogo Marino (Universidad Jorge Tadeo Lozano), bajo la dirección de Jaime Garzón-Ferreira y Sonia Bejarano.
BIBLIOGRAFÍA
1. Alcolado, P., A. Corvea y A. González. 1980. Variaciones morfológicas internas y externas de los abanicos de mar Gorgonia spp. y su valor adaptativo. Cienc. Biol., 5: 47-56. [ Links ]
2. Ayre D.J. y B.L. Willis. 1988. Population structure in the coral Pavona cactus clonal genotypes show little phenotypic plasticity. Mar. Biol., 99: 495-505. [ Links ]
3. Anthony, K.R.N. 1999. A tank system for studying benthic aquatic organisms at predictable levels of turbidity and sedimentation: Case study examining coral growth. Limnol. Oceanogr., 44: 1415-1422. [ Links ]
4. Ardila, N., G. Navas y J. Reyes. 2002. Libro rojo de los invertebrados marinos de Colombia. INVEMAR Santa Marta, 180p. Serie de Libros Rojos de Fauna amenazada de Colombia. Tomo 1. [ Links ]
5. Bayer, F. M. 1961.The shallow-water Octocorallia of the west Indian region. Martinicus Nijhoff, La Haya, 373p. [ Links ]
6. Bejarano-Chavarro S., N. Manrique-Rodríguez y J. Garzón-Ferreira. 2005. Recent partial mortality and other health conditions of the sea fans Gorgonia ventalina Linnaeus 1758 in the Santa Marta area, Colombian Caribbean, 15 years after a mass mortality event. Proc. 10th Int. Coral Reef Symp. Japón. p. 982-988. [ Links ]
7. Birkeland, C. 1974. The effect of wave action on the population dynamics of Gorgonia ventalina Linnaeus. Stud. Trop. Oceanogr. 12: 115-126. [ Links ]
8. Botero, L. 1987. Gorgonian octocoral communities of the Santa Marta area, Caribbean coast of Colombia: species composition, patterns of zonation and quantitative structure. PhD thesis, University of Delaware, United States. 136 p. [ Links ]
9. Botero, L. 1990. Quantitative structure of gorgonian communities in Tayrona National Park, Caribbean coast of Colombia. An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México, 17(1): 73-88. [ Links ]
10. Cairns, S. 1977. Guide to the commoner shallow-water gorgonians (sea whips, sea feathers and sea fans) of Florida, the Gulf of Mexico and Caribbean Region. Miami Univ. Sea. Grant. Field. Guide., 6: 1-74. [ Links ]
11. Díaz, J.M., J. Garzón-Ferreira y S. Zea. 1995. Los arrecifes coralinos de la Isla de San Andrés. Colombia: Estado actual y perspectivas para su conservación. Academia Colombiana de Ciencias Exactas Físicas y Naturales, Colección Jorge Álvarez Lleras 7, Santa fé de Bogotá, 150p. [ Links ]
12. Díaz, J. M., L.M. Barrios, M. Cendales, J. Garzón-Ferreira, J. Geister, M. López, G. Ospina, F. Parra-Velandia, J. Pinzón, B. Vargas, F. Zapata y S. Zea. 2000. Áreas coralinas de Colombia. INVEMAR, Santa Marta, Ser. Publ. Espec., 5: 176 p. [ Links ]
13. Garzón-Ferreira J. y M. Cano. 1991. Tipos, distribución, extensión y estado de conservación de los ecosistemas marinos costeros del Parque Nacional Natural Tayrona. Versión presentada al Séptimo Concurso Nacional de Ecología, "Enrique Pérez Arbelaez", Fondo para la Protección del Medio Ambiente - FEN COLOMBIA Bogotá, 82 p. [ Links ]
14. Garzón-Ferreira, J. y S. Zea. 1992. A mass mortality of Gorgonia ventalina (Cnidaria: Gorgoniidae) in the Santa Marta area, Caribbean coast of Colombia. Bull. Mar. Sci., 50: 522-526. [ Links ]
15. Geiser D.M., J.W. Taylor, K. B. Ritchie y G.W. Smith. 1998. Cause of sea fans death in the West Indies. Nature, 394: 137-138. [ Links ]
16. Grigg, R. 1972. Orientation and growth form of sea fans. Limnol. Oceanogr., 17(2): 185 - 192. [ Links ]
17. Guzmán, H. M. y J. Cortés. 1984. Mortandad de Gorgonia flabellum linnaeus (Octocorallia: Gorgonidae) en la costa Caribe de Costa Rica. Rev. Biol. Trop., 32(2): 305-308. [ Links ]
18. Harvell, D., J. Ward, N. Douglas, G. Vasta, J. Bruno y K. Kim. 2004. Enviromental modulators of sea fan inducible resistance to fungal disease. Corson Hall, Cornell University, Ithaca, New York, USA. Abst. 10th Int. Coral Reef Symp. 452 p. [ Links ]
19. Kim. K. y H.R. Lasker. 1998 Allometry of resource capture in colonial cnidarians and constraints on modular growth. Funct. Ecol., 12: 646-654. [ Links ]
20. Lasker, H. y M. Coffroth, 1983. Octocoral distributions at Carrie Bow Cay, Belize. Mar. Ecol. Prog. Ser., 13: 21-28. [ Links ]
21. Lasker H.R., M.L. Boller, J. Castanaro y J.A Sánchez. 2003. Determinate growth and modularity in a gorgonian octocoral. Biol. Bull., 205: 319-330. [ Links ]
22. Laydoo, R. 1983. Recent mass mortality of gorgonians in Trinidad. Unpub. Rep., Inst. Mar. Affairs, Trinidad, W. Indies. 12 p. [ Links ]
23. Manrique-Rodríguez, N. 2004. Estado actual y crecimiento de Gorgonia ventalina (Linnaeus, 1758) (Cnidaria:Gorgoniidae) en el área de Santa Marta, Caribe colombiano. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá, 85 p. [ Links ]
24. Nagelkerken, I., K. Buchan, G. W. Smith, K. Bonair, P. Bush, J. Garzón-Ferreira, L. Botero, P. Gayle, C. Hereber, C. Petrovic, L. Pors y P. Yoshioka. 1997a. Widespread disease in Caribbean sea fans: I. Spreading and general characteristics. Proc. 8th Int. Coral Reef Symp., Panamá, 1: 679-687. [ Links ]
25. Nagelkerken, I., K. Buchan, G. W. Smith, K. Bonair, P. Bush, J. Garzón-Ferreira, L. Botero, P. Gayle, C. D. Harvell, C. Hereber, K. Kim, C. Petrovic, L. Pors y P. Yoshioka. 1997b. Widespread disease in Caribbean sea fans: II. Patterns of infection and tissue loss. Mar. Ecol. Prog. Ser., 160: 255-263. [ Links ]
26. Ramírez, G. 1990. Distribución de nutrientes inorgánicos en las aguas costeras de la región de Santa Marta, Caribe colombiano. VII Seminario Nacional de Ciencias y Tecnologías del Mar. p. 244-254. [ Links ]
27. Rinkevich, B. 2000. Steps towards the evaluation of coral reef restoration by using small branch fragments. Mar Biol., 136: 807-812. [ Links ]
28. Sánchez, J. A. 1995. Benthic communities and geomorphology of the Tesoro Island reef, Colombian Caribbean. An. Inst. Invest. Mar. Punta Betín., 24: 55-77. [ Links ]
29. Sánchez, J. A., J. M. Díaz y S. Zea. 1997. Gorgonian communities in two contrasting environments on oceanic atolls of the southwestern Caribbean. Bull. Mar. Sci., 61(2): 453-465. [ Links ]
30. Sánchez, J. A. 1999. Black coral-octocoral distribution patterns on Imelda Bank, a deep-water reef, Colombia, Caribbean Sea. Bull. Mar. Sci., 65(1): 215 -225. [ Links ]
31. Sebens, K.P. 1987. The ecology of indeterminate growth in animals. Annu. Rev. Ecol. Syst., 18: 371-407. [ Links ]
32. Smith, G. W., L. D. Ives, I. Nagelkerken, y K. B. Ritchie. 1996. Caribbean sea fan mortalities. Nature, 383: 487-495. [ Links ]
33. Sokal, R. y F. J. Rohlf. 1979. Biometría: Principios y métodos estadísticos en la investigación biológica. Editorial Blume, Madrid 831 p. [ Links ]
34. Yoshioka, P. M. y B.B. Yoshioka. 1987. Variable effects of hurricane David on the shallow water gorgonians of Puerto Rico. Bull. Mar. Sci., 40(1): 132-144. [ Links ]
35. Yoshioka, P. M. y B.B. Yoshioka. 1991. A comparison of the survivorship and growth of shallow-water gorgonian species of Puerto Rico. Mar. Ecol. Prog. Ser., 69: 253-260. [ Links ]
36. Wainwrigth, S. A. y J. R. Dillon. 1969. Orientation of sea fans. Biol. Bull. Mar. Biol. Lab., Woods Hole. 136: 130-139. [ Links ]
37. Willis B.L. 1985. Phenotypic plasticity versus phenotypic stability in the reef corals Turbinaria mesenterina and Pavona cactus. Proc. 5th Int. Coral Reef Symp., Tahiti 4: 107-112. [ Links ]
FECHA DE RECEPCIÓN: 19/10/05 FECHA DE ACEPTACIÓN: 20/09/06
