










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.35 no.1 Santa Marta Jan./Dec. 2006
MODEL-BASED GEOMORPHOLOGY OF MALPELO ISLAND AND SPATIAL DISTRIBUTION OF BREEDING SEABIRDS
MODELACIÓN GEOMORFOLÓGICA DE LA ISLA MALPELO Y DISTRIBUCIÓN ESPACIAL DE AVES MARINAS ANIDANTES
Mateo López-Victoria1 and Daniel M. Rozo2
1Department of Animal Ecology and Special Zoology, University of Giessen (JLU), Germany. E-mail: Mateo.Lopez-Victoria@bio.uni-giessen.de
2Instituto de Investigaciones Marinas y Costeras-INVEMAR (Marine and Coastal Research Institute), Cerro Punta Betín, Santa Marta, Colombia. E-mail: drozo@invemar.org.co
ABSTRACT
The spatial distribution of seabird nests present on Malpelo Island, the main characteristics of the island's topography, and the types of substratum available for nesting were addressed using a Digital Elevation Model (DEM). The model is based on data from remote sensors (satellite images, aerial photography and panoramic photography), and from field data processed through a geographical information system. The nests of Sula granti prevail on the surfaces of Malpelo Island and neighboring islets, while those of the remaining species (S. sula, Creagrus furcatus, Anous stolidus, A. minutus and Gygis alba) are spatially restricted to the cliff margins and to crevices or caverns at different heights. The presence of terrestrial predators and predatory birds (Fregata spp.) seems to mainly explain the marginal distribution of other breeding species. The cartographic data presented here differ significantly from past and current representations, especially those concerning the island's emerged surface and other geomorphologic features. This new cartographical information will support future studies on the terrestrial fauna and flora of Malpelo Island and its islets.
KEY WORDS: Seabirds, Nest distribution, Digital Elevation Model, Malpelo Island, Colombia.
RESUMEN
La distribución espacial de nidos de aves marinas en la Isla Malpelo, las principales características del relieve del suelo de la Isla y los tipos de sustratos disponibles para anidación, fueron estudiados utilizando un modelo digital de elevación (MDE). El modelo está basado en datos de sensores remotos (imágenes de satélite, fotografías aéreas y fotografías panorámicas), y en datos de campo procesados mediante un sistema de información geográfico. Los nidos de Sula granti prevalecen en las superficies de Malpelo e islotes vecinos, mientras que aquellos de las restantes especies (S. sula, Creagrus furcatus, Anous stolidus, A. minutus and Gygis alba) están espacialmente restringidos a los márgenes de los acantilados y a grietas o cavernas a distintas alturas. La presencia de depredadores terrestres y aves depredadoras (Fregata spp.) parecen ser las principales explicaciones de la distribución marginal de esas otras especies anidantes. Los datos cartográficos aquí presentados difieren significativamente de pasadas y actuales representaciones, especialmente en lo que concierne al área emergida de la Isla y a otros aspectos geomorfológicos. Esta nueva información cartográfica pretende apoyar futuros estudios sobre la fauna y flora terrestres de Malpelo y sus islotes.
PALABRAS CLAVE: Aves marinas, Distribución de nidos, Modelo digital de elevación, Isla Malpelo, Colombia.
INTRODUCTION
Malpelo Island is the top of a submerged mountain chain (Stead, 1975) and its topography relief is uneven and rough, giving the impression of an inaccessible and inhospitable naked rock (Townsend, 1895; Graham, 1975). Due to its remote location from the Pacific coast of Colombia and difficult access across its uneven and rough surfaces, research work conducted on the island has been relatively scarce, and in consequence, the cartographical information remains incomplete and poor. Up to date, only some bi-dimensional representations, with contour lines, have been made, including cartography regarding the island geomorphology and geology. These representations, on which all existing estimations of Malpelo's surface and its inhabiting fauna are based, were made more than three decades ago by the Smithsonian Institute (Kiester and Hoffman, 1975).
For the specific case of Malpelo, the traditional two-dimensional mapping methods provide incomplete information on the location, basic area, and contour lines of the island. The lack of information on the surface inhabited by different organisms, and their influence areas, difficult the estimation of their density, total population and biomass. For instance, research on the resident birds of Malpelo demands a reliable cartographical base to support this and any biological study, because the direct estimation of the total number of individuals is not possible due to the uneven topography of the island (López-Victoria and Estela, 2006).
Malpelo is home of the greatest breeding colony of the Nazca Booby (Sula granti) in the world and harbors populations of at least other six breeding bird species, the Masked Booby (Sula dactylatra), the Red-footed Booby (Sula sula), the Swallow-tailed Gull (Creagrus furcatus), the Brown Noddy (Anous stolidus), the Black Noddy (Anous minutus) and the White Tern (Gygis alba) (Pitman et al., 1995; Pitman and Jehl, 1998; Álvarez-Rebolledo, 2000; López-Victoria and Estela, unpublished). The ecology of these seven seabird species, as well as of other 50 bird species, including further seabirds and small migratory birds, have been subject of study since the end of the XIX century (Townsend, 1895), and especially during the last 20 years (Pitman et al., 1995; Álvarez-Rebolledo, 2000; López-Victoria and Estela, 2006). However, the housing requirements of resident birds have been poorly studied in terms of nesting space and location, due to the difficulties to conduct field work on the island.
The use of remote sensors and geographical information systems has proved to be a practical tool used in different ornithological studies during the last decades (Gottschalk et al., 2005). Adequate graphical representations can be obtained through a Digital Elevation Model (DEM), defined as a numerical structure of data, representing the spatial distribution of the elevation of the ground surface (Felicísimo-Pérez, 1994). This technique combines the information obtained from remote sensors, the data collected in the field and the existing cartography.
The purpose of this study was to develop a DEM for Malpelo Island and neighboring islets, based on field data and information from remote sensors. An approximated cartographical data base was generated in order to: 1) estimate the resident seabird requirements in terms of nesting spatial distribution and 2) describe the main characteristics of the island's topography and substratum, as a function of the area occupied by nests. This new cartographical data base is also intended to support future studies on the terrestrial fauna and flora of Malpelo Island and neighboring islets.
MATERIALS AND METHODS
From October 2003 thru June 2006 seven visits to Malpelo were made. Location and elevation of relevant spots identified on the island's surface were recorded, using a GPS Garmin Map76S (provided with barometer). The length and width of the eastern, central and southern areas of Malpelo were surveyed, and location [error (x, y) less than 15 m] and elevation [error (z) less than 3 m] of the ground relief were recorded. Additionally, panoramic and detailed photographs were taken from different angles of the islands. High resolution (126 mm format) oblique aerial photographs, taken by the Southwest Fisheries Science Center-SWFSC (Pitman et al., 1995), were also obtained. The general outline and location of the islands were estimated from an image of an Aster satellite (NASA) with a resolution of 15 m (ASTER On-Demand L2 Surface Reflectance).
Data processing and analyses were carried out using the ArcGIS 9.1 software. Field data were loaded to the system of coordinates UTM zone 17N and organized in a same vector map (Shapefile format). The geographical points of GPS taken in the field were used as basis for the geographical reference of the satellite image utilizing the mode for geo-reference of the ArcMap within the system wgs84. A preliminary coastal line was digitalized according to the outline interpreted on the reflectance band. Using the preliminary coastal line as a base line and the positioning data taken in the field, the oblique aerial photos were geo-referenced, giving priority to the coastal line. Based on the finished geo-referenced photography, a photo-mosaic was assembled, covering the majority of the islands (the northern islets and the most remote islet from the eastern coast were excluded from the high resolution aerial-photograph set; Figure 1).
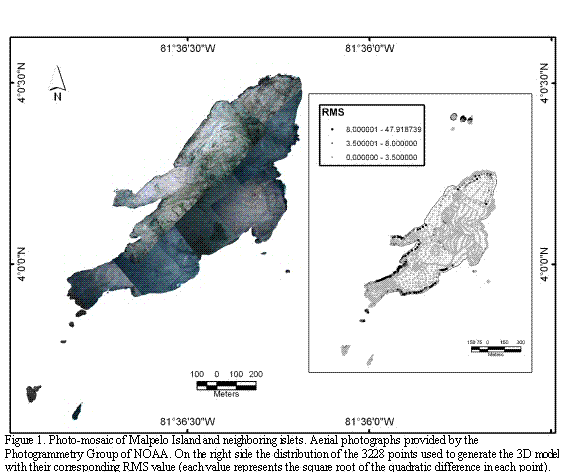
From the aerial photo-mosaic, the coastal line of the islands was re-digitalized with as much detail as possible. Subsequently, the data were standardized and then loaded, including information on position and elevation derived from transects (tracks) made through Malpelo and recorded in the GPS. The data-base obtained using the GPS was complemented using the panoramic photographs taken from boats at different angles and distances from the islands. Using this information, the elevations were established based on the changes of relevant spots of the physiognomy of the islands. These values were calculated by means of linear regression, comparing known elevations to easily identifiable sites of the aerial photography mosaic.
To prevent the edge effect and the deformation of the interpolation due to lack of data on the coastal line, besides estimating the steep slopes of the islands land, a bathymetric file was created using the data of the chart 521 of the Centro de Investigaciones Oceanográficas e Hidrográficas (CIOH).
Once a total of 3228 geographical points were registered, an irregular net was generated using a triangulation of Delaunay (surfaces model type TIN) in order to control the loading of data elevations (Figure 1). Subsequently, a digital elevation model (one meter of resolution) of the islands was generated, based on the technique of the finite iteration of the difference interpolation. The algorithm "topo to raster" implemented in ArcGIS 9.1 was used to simulate the island's surface based on the map of geographical points with their estimated elevations, which was the starting point for generating the contour lines (one meter apart). This sequence was used to determine a triangulated surface (TIN) with minor changes, compared to the direct generation of elevation points.
Preliminary maps were used to mark the main geo-morphological characteristics of Malpelo on the field, as a function of the nests' distribution of the different seabird species (except Masked Booby). Starting from these data and using the geo-referenced mosaic of aerial photography and the contour lines obtained from the estimated surface, the physical limits of zones with homogenous characteristics were digitalized. In addition to field information, the oblique aerial and panoramic photographs were taken into account to establish those limits. Data related to the location vectors of the vegetal coverage, the substratum stability and roughness as a function of the nests presence (four categories), the utilization of the islands by humans (four categories), and the distribution of the nesting sites for six seabirds were loaded (Nazca Booby, Red-footed Booby, Swallow-tailed Gull, Brown Noddy, Black Noddy and White Tern). Through the tool "surface volume" implemented in ArcGIS 9.1 the surface areas (3D) were estimated for each of the attributes of each thematic and a closed analysis was conducted to establish if the error would be acceptable.
Based on the DEM an analysis of slopes was conducted and the surfaces were calculated in two and three dimensions for all the evaluated categories, combining the TIN surface with each of the thematic attributes. Finally, the graphic output for each thematic was generated (maps, views and profiles in 2D and 3D).
RESULTS
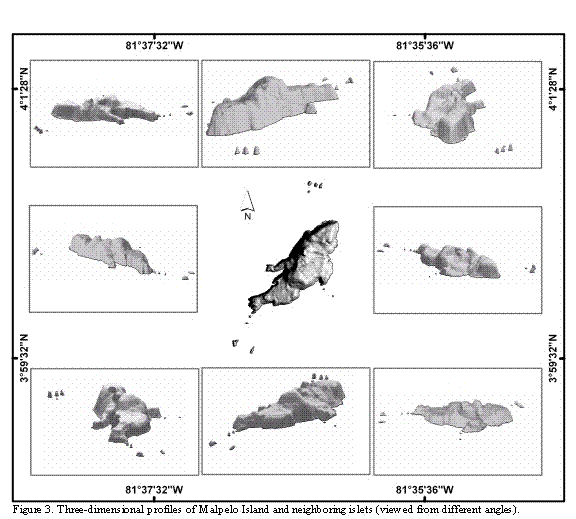
Cartographic data collected for the present study show that Malpelo is an island of elongated form, about 1643 m long and with variable width (maximum width approximately 727 m; Figure 2). The main island is neighbored by 11 to 12 islets, depending on what definition is used for the portions of rock emerging from the sea in the periphery. At least two of the islets are relatively close to the island and one of them is small and low (see farther below). The islets were designated with different names during the last century. We opted to use the names currently given, and widely accepted by divers (Harold Botero. Embarcaciones Asturias de Buenaventura, Colombia. Sandra Bessudo and Krupskaya Narváez. Fundación Malpelo. Bogotá, Colombia. 2006. Pers. Comm.). The islets' distribution and their names are as follows: four islets located 500 m from the northern end of the island, known as Los Mosqueteros; five islets located at different distances from the island's southern point, known as Tres Reyes, La Gringa, and Escuba; two islets located at the eastern coast of the island, in front of an arch or natural bridge, known as Vagamares and La Torta. The last islet, located at the western coast of the island, and considered by some people as a shoal, is known as El Mirador. This last islet and the first one of the southern islets (Salomón) are separated from the main island by, maximum, six and two meters, respectively (Figures 2 and 3).
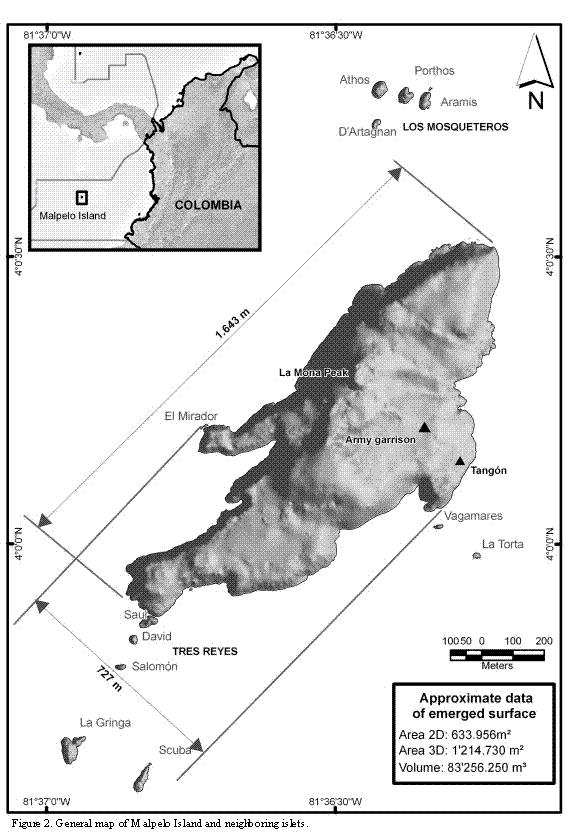
The flat (cartographical) emerged area of the islands (Malpelo and the islets) is about 0.633 km2 (ca. 63 Ha). However, their surface area (3D) is about 1.215 km2 (ca. 121 Ha), i.e. almost twice as large. This large difference results from the uneven ground topography of the islands, and the presence of hills with a maximum height of about 300 m. The northern half of the main island is higher than the southern half and on the top of one of the hills, known as La Mona, a light house was erected (Figures 2 and 3).
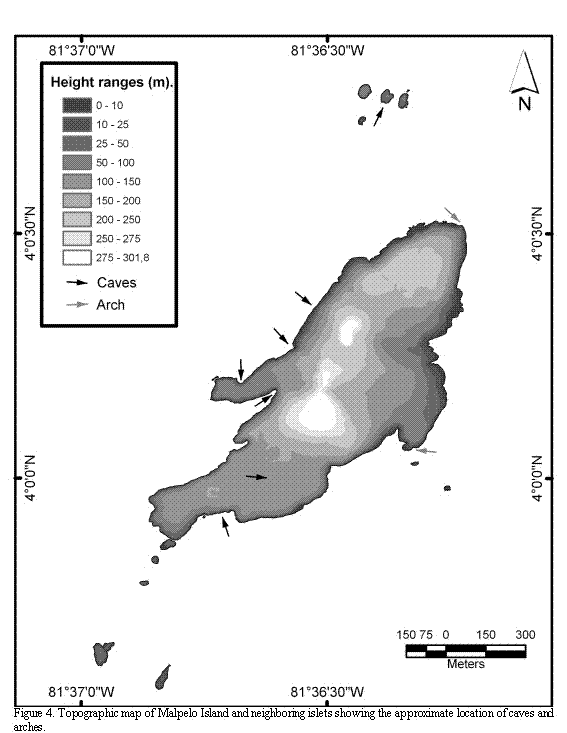
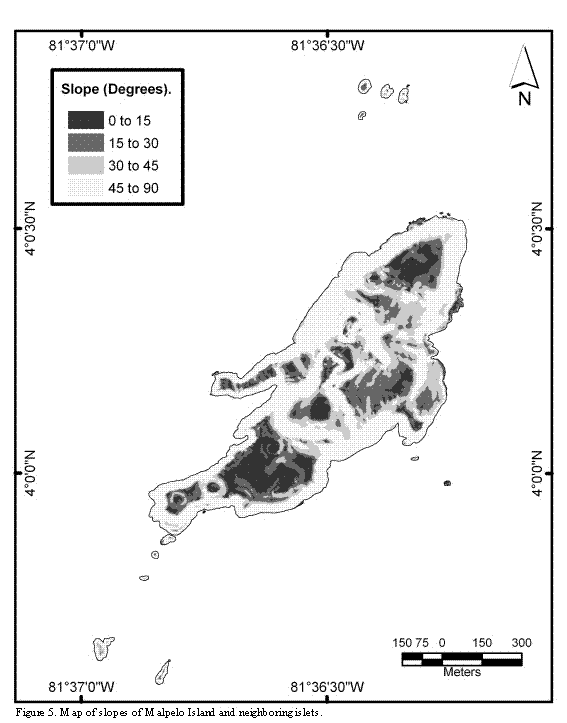
The uneven relief of the island's ground presents several steep slopes, which constitute 72% of the islands surfaces. With the exception of some areas of the northeast of the main island, the cliffs extend 20 to 80 m deep under the sea. There are some areas that are almost flat, located on the southern part of the main island, on the eastern coast (where two navy houses were built), on the higher points, and on the highlands of the northern end of the island. With the exception of La Torta and El Mirador, having an emerged flat relief with surfaces completely swept by the swell and the high tides, all the islets are sharp pointed, rise above the sea level more than five meters, and show an uneven ground relief (Figures 4 and 5).
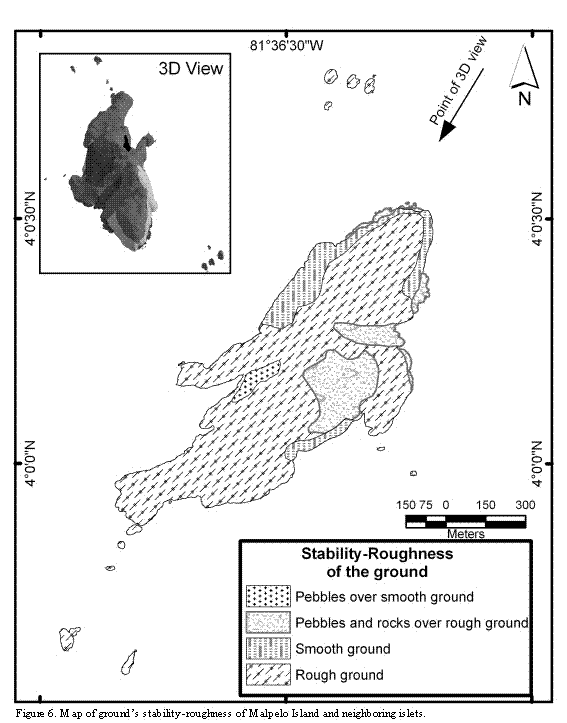
Near 72% of the island's ground surface is solid rock of uneven relief; the remaining 28% of the area is plain rock or loose rock of different sizes (Figure 6). In those surfaces where more rock accumulation is found the slopes are smaller, and rocks falling from the hills are piled up there. These areas also correspond to watersheds, which contribute to the flow of water and rocks into the sea. A reduced number or complete absence of nests in these areas was observed.
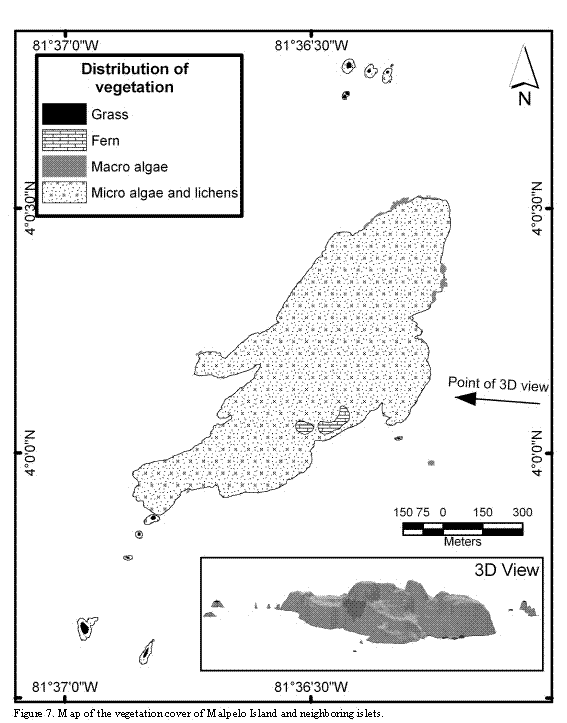
Except for some scattered patches of grasses and ferns, not exceeding an area of 0.02 km2, the vegetal cover of the islands constitutes micro-algae, mosses and lichens. The largest patch of vegetation (vascular plants), comprising the fern Pityrogramma dealbata is located at the center of the main island, towards the western coast, and between 100 and 150 m above the sea level. The grass patches (Poaceae) occur mainly on the top of the islets, and inside crevices and caverns of the hills skirts (Figure 7).
Less than 3% of the main island is frequently used by humans and near 90% is sporadically visited. Inhabited areas correspond to those occupied by houses of the Colombian Navy, to the neighboring pathway linking the access dock to the barracks and, occasionally, the pathway to La Mona Hill. Colombian Navy squads or research teams sporadically explore the southern region of the main island and the hills located at the central part.
The Nazca Boobyis the most abundant species in Malpelo, having nests on almost all islets, with the exception of El Mirador and La Torta (Figure 8). Most nests are found on surfaces with moderate slopes. In those areas with steep slopes the nests are scarce and scattered. Disregarding the general slopes, the micro-relief of the ground where nests occur is flat and made out of solid rock. Nests are made out of pebbles and, in
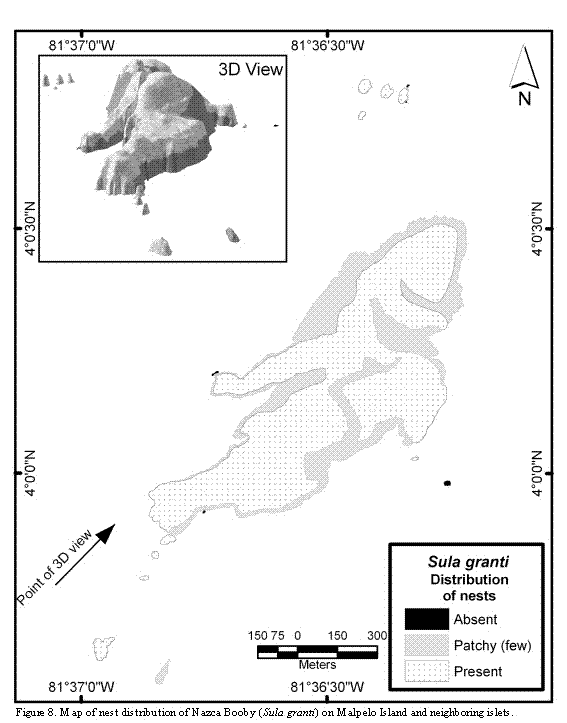
those steep-sloped areas they are located on small terraces or cornices. On the other hand, in those areas where stones or boulders are piled up in great amounts and few places of solid rock are available, nests are scarce and scattered or completely absent.
The Red-footed Booby is present throughout most of the year in the island but is not abundant. In none of the visits a group larger than 60 adult individuals was spotted. All nests are located on the southern part of the main island, on two low areas comprised by fractures of the rock's surface. There are also some nests inaccessible for humans on the highlands of Athos, the largest northern islet (Figure 9). Nests located on the southern part of the main island are made out of grass, apparently brought by males from the northern
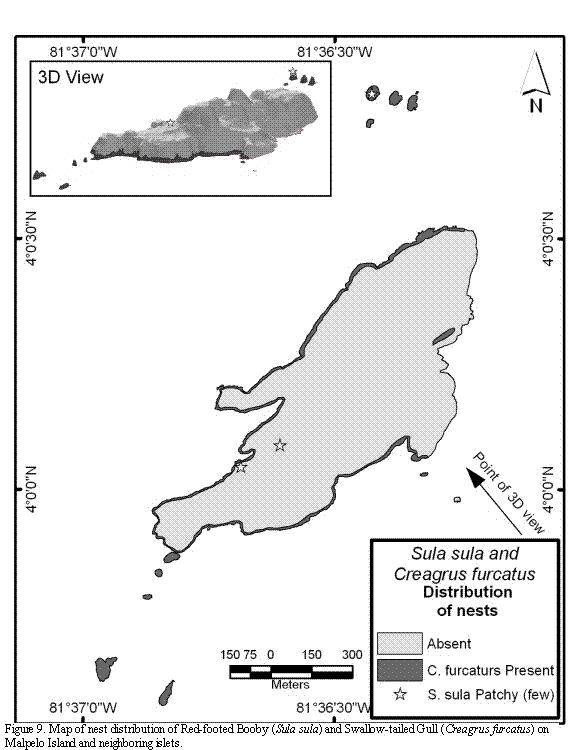
and southern islets. These scarce grasses are placed at the top of small rocky, pedestal or pinnacle-shaped promontories, between 30 and 50 cm in diameter, and between 70-100 cm height. Terrestrial crabs (Gecarcinus malpilensis) were frequently observed feeding on nest materials of this colony, and no apparent reaction from the birds to counteract the crab's predatory activity was observed. It was not possible to visit the nests located on the northern islet, but observations made with binoculars from the water surface suggest that they are located directly on or at the side of small grass patches.
The nesting distribution of Swallow-tailed Gull is restricted to a distance of 10 to 70 m from the water surface. Nests are scattered and located on small cornices, terraces or crevices of the cliffs, always on solid ground, covered with small stones and small fragments of invertebrates (mostly crabs). In the north-east of Malpelo, in the sector known as Tangón, not a single couple of this species in breeding behavior was observed (Figure 9).
The nesting distribution of Brown Noody and Black Noody is sympatric and extends up to the first 5 to 20 meter strip of the northern islets, the coast cliffs, and the north-east and mid-east part of the main island (Figure 10). Brown Noddy also builds some scattered nests at the interior of the main island, located at the upper part of the hills and large crevices. As in the case of Swallow-tailed Gull, these two small, tern-like birds use the cliff's terraces, cornices and crevices for nesting. Although Brown Noddy may or may not build nests to lay its egg, most of them were simple structures, several of them had small stones and remains of invertebrates (mostly Grapsidae crabs), and two were built of feathers of the Nazca Booby and plastic remains (in one occasion with a tooth brush). On the other hand Black Noddy did elaborated nests on all the occasions observed. All nests checked were cup-shaped, and were built with remains of grasses (Poaceae) and synthetic fibers of different kinds (nylon ropes, fishing-mesh, etc.) picked up from the surface of the water, and cemented with excrements. In the Vagamares, La Torta, and El Mirador islets nests were absent.
White Tern nests on the same areas as Noddies do, i.e., on small terraces, cornices and crevices of the cliffs. This bird does not build nests and chooses it's nesting places above 7 to 10 meters high, up to the hill's peaks (Figure 10). Opposite to the case of the two Noddies species mentioned above, White Tern does not breed colonially. Nesting activities were not observed in La Torta, Vagamares and El Mirador islets.
DISCUSSION
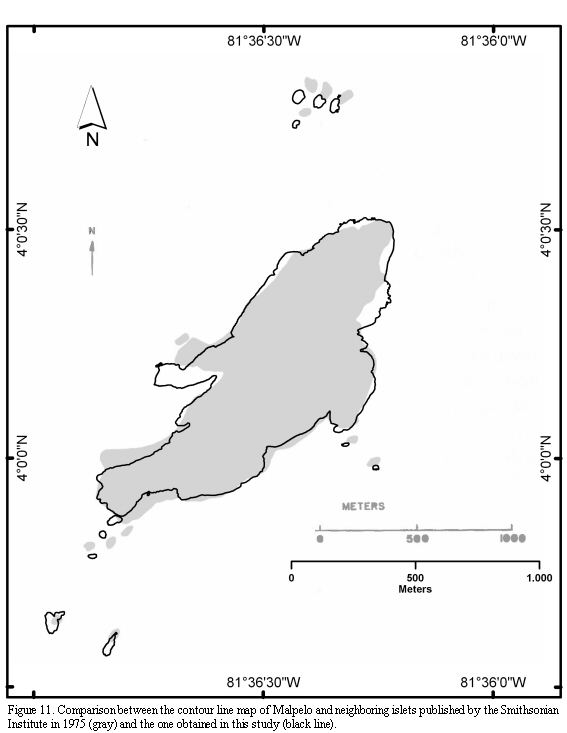
The most accurate cartography on Malpelo consisted of a nautical chart based on data collected by a US-Navy flight in 1954. This preliminary map was complemented and enhanced during a research cruise carried out by the Smithsonian Institute and the US-Navy in 1972 (Kiester and Hoffman, 1975). Comparison of that cartography with the
information obtained in this study allow shedding light on the following four general, significant differences: a) the southern end of Malpelo is narrower, b) the Tres Reyes and La Gringa islets are lined up between them, c) the shape, location and size of most islets is different, and d) the small peninsula located at the mid west of Malpelo has a different orientation and is longer (Figure 11). A further difference between results obtained in this study and those from the study carried out by the Smithsonian regards the island's maximum height, reported by this institution as 376 m (some conversions from the original figure in feet result in about 400 m; Pitman et al., 1995), which is, at least, 76 m higher than the 300 m estimated in this study. These differences result from the triangulation method used by the Smithsonian to measure the heights from the ship. Our results agree with the measures of about 300 m reported by Sarmiento (1953), and those obtained by Caita and Guerrero (2000) using a GPS, in which a figure depicting the hill Cerro La Mona measuring 296 m (±10 m) high, is shown.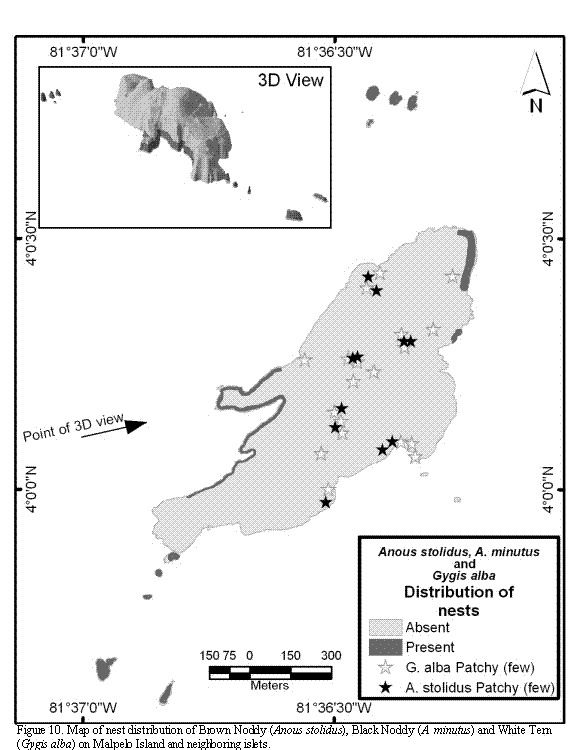
In a number of studies, the emerged flat area of Malpelo has been reported as varying between 35 and 63 Ha (0.35-0.63 km2) (Sarmiento, 1953; Prahl, 1990; Brando et al., 1992; Álvarez-Rebolledo, 2000; Caita and Guerrero, 2000; Díaz et al., 2000). Results obtained in this study show values that coincide with the higher previously reported data for the flat area (cartographic), and even double that given surface area (3D= 1.2 km2). These differences have important implications when estimating the populations of the different organisms living on the islands, especially, when density values are extrapolated. Despite its small size, walking on the surface of Malpelo makes direct observations difficult or impossible, and therefore it is mandatory to know the surface area value to make biological estimations of the different populations as accurate as possible.
The number of islets neighboring Malpelo has been reported with different values, depending on the selected classification system. For instance, Sarmiento (1953) reported six islets, while in the maps published by Kiester and Hoffman (1975) twelve islets are shown (Figure 11); Prahl (1990) mentioned eleven islets while Álvarez-Rebolledo (2000) reported ten. A total of eleven islets, with emerged areas during high tide, and separated from the main island, are currently recognizable. The first southern islet is so close to the main island that, probably, for several researchers, it does not appear as an independent land portion and, hence, the discrepancies found in the literature. In addition, El Mirador is so low that, during heavy swell it is completely covered with water and therefore could be excluded as an islet; on the other hand, it is not a shoal either. In some areas several rocks fallen from the main island and separated by a few centimeters from the coast line are observed, though they cannot be classified as islets.
Another confusing aspect regards the distance from Malpelo to the Pacific coast of Colombian mainland. According to the calculations made in this study, the nearest point (to the Pacific coast of Colombia) is the Corregimiento Playa del Caballo, Mosquera municipality, Nariño department, about 380 km from the main island. On the other hand, the distance from the main island to Buenaventura sea port, located almost at the same latitude of Malpelo, is about 497 km (data obtained from the LabSI-INVEMAR; according to the coast line reported by the Instituto Geográfico Agustín Codazzi-IGAC).
Comparison of panoramic photographs taken during the last century with more recent aerial photos reveals an evident change of the largest vegetation patch formed by the fern Ptyrogramma dealbata. The first researchers did not mention its presence (Slevin, 1928; R. Smith in Bond and Meyer de Schauensee, 1938; Murphy, 1945), although currently it is visible from the ships' anchoring places, close to the eastern coast of the main island. Subsequently, the vegetation is reported in the publication of the Smithsonian Institute, in which the scientific name is recorded for the first time, though without making mention of its coverage (Graham, 1975). Interestingly, in the aerial photographs of 1988 and in some other photographs published in 1990 (Prahl) and 1992 (Brando et al.), the vegetation patch appears larger than the one currently observed. The time of arrival of this fern is uncertain but it is possible that some spores reached the islands swept by winds from Central America; a further possibility is that the first human visitors that traveled among the eastern tropical Pacific islands, introduced the spores adhered to their clothes or travel gear. The presence of the terrestrial crab (Gecarcinus malpilensis) may explain the apparent reduction of the vegetation coverage but this should be confirmed experimentally. On the other hand grass patches growing on the picks of the islets and the presence of algae, lichens and mosses have been mentioned in the literature since early XX century (see Graham, 1975).
The Nazca Booby breeds on the coastal margins of the Galápagos, on areas almost lacking vegetation and close to cliffs (Nelson, 1978; Duffy, 1984; Townsend et al., 2002), apparently due to the need of high and windy places for taking off (Duffy, 1984). In Malpelo, on the other hand, nests are distributed everywhere because the almost complete lack of vegetation and the uneven topography make most of the island's surface available for nesting. Therefore, nest distribution in Malpelo does not seem to be restricted by space, distance to the sea or nest location with respect to the wind; on the contrary, the ground slope and substratum stiffness seem to play an important role in this context.
The Red-footed Booby nests in colonies, almost always on trees or shrubs and, rarely, on grass thickets or even on artificial structures when no other vegetation is available (Schreiber et al., 1996). The Malpelo colony is small and atypical; the building process of nests is very rudimentary and the eggs are laid on the rock ground, because the nest materials are frequently removed and eaten by crabs. The two colonies of this species closest to Malpelo are located on Cocos Island (Costa Rica) and the Galápagos Islands (Schreiber et al., 1996).
The Swallow-tailed Gull nests on the Galápagos Islands under conditions similar to those found in Malpelo (Harris, 1970). The only difference is the maximum height at which nests are located, as that in Malpelo appears to be lower, probably due to the pressure exerted by terrestrial predators (crabs and lizards).
The Brown and Black noodies nest sympatrically in other colonies on cliff cornices as well as on shrubs and trees (Gauger, 1999). The Malpelo colony differs from other colonies because it comprises few dozens of couples; contrastingly, other colonies comprise thousands of couples (Gauger, 1999). The relative low number of couples found in Malpelo may be best explained by the presence of the terrestrial crab and at least one of the lizards (Diploglossus millepunctatus), as well as by the pressure exerted by the frigate birds (Fregata magnificens and F. minor) on the broods of these two species, if nests are located on exposed areas; when nests are protected inside crevices or cornices, it is very difficult for frigatebirds to hunt the broods (Gauger, 1999). Another factor affecting the nest location of the Anous species is the wind pattern that predominates in Malpelo during the second half of the year (winds blowing from the south and the south-west). These species are bly affected by heavy winds and thunders, which kill broods as well as adult individuals (Chardine and Morris, 1996; Gauger, 1999). These species, as well as the other two Laridae species present in Malpelo, nest mostly on sectors protected from tropical winds and thunders characteristic of this season.
The White Tern shares some nesting sectors with the two noddies species in Malpelo, though not at heights within the cliff area. This may be explained because noddies out-compete White Tern for nesting sites, as observed in the colony of Tern Island (Niethammer and Patrick-Castilaw, 1998). Nevertheless, other observations point out that Black Noody is not able to successfully compete for nesting areas against White Tern, which frequently takes up its nests (Gauger, 1999). In any case, White Tern is not colonial as are both noddies species (Chardine and Morris, 1996; Niethammer and Patrick-Castilaw, 1998; Gauger, 1999), and hence, several nests of this species are scattered throughout the whole area of Malpelo, including the top of the hills.
In general terms, data on the nesting distribution of the seabirds of Malpelo collected in this study agrees with those reported in the literature (Harris, 1970; Anderson, 1993; Chardine and Morris, 1996; Niethammer and Patrick, 1998; Gauger, 1999). The predominance of Nazca Booby in most of the islands' surfaces seems to be explained by the following main factors: a) it is the largest bird, in relation to other birds present in the island, and out-competes other species for nesting space (i.e. Red-footed Booby), b) it appears to be the only species in Malpelo able to counteract a permanent predatory pressure from the terrestrial crab and one of the lizards (Diploglossus millepunctatus), and c) it is the best adapted, compared to other birds breeding on the island, as it finds ideal conditions for nesting (i.e., doesn't require vegetation for nesting). In the case of the other species, although there are favorable conditions for nesting on many sectors of the islands' surfaces, they appear to be unable to resist the pressure exerted by crabs and lizards. In those areas where these predators have been observed, only nests of Nazca Booby were found; contrastingly, no crabs or lizards were observed in those areas where nests of other species were found. These evidences are circumstantial and should be confirmed experimentally.
Future studies on the population biology of plant and animal species present in Malpelo should consider as a starting point the current surface area of the island and its uneven geomorphology. More accurate approaches to the DEM will be achieved by loading the model with inputs from remote sensors of better resolution (i.e., radar images and/or aerial photography). The northern sector of Malpelo is still poorly explored and requires more detailed cartographic research.
ACKNOWLEDGEMENTS
This investigation was partially supported by INVEMAR, the association CALIDRIS, COLCIENCIAS (Project 2105-09-13527-03), the Department of Animal Ecology and Special Zoology, Justus-Liebig-University Giessen, Germany (JLU), the German Academic Exchange Service (DAAD), the Malpelo Foundation (in special S. Bessudo, G. Soler and Y. Lefèvre), the Colombian Ministry of Environment, Housing and Territorial Development, the Special Administrative Unit for the System of National Natural Parks of Colombia, the National Navy of Colombia (especially, the marine corps in Malpelo) and the Training Course SUCOMAR 2005, offered by InWent. This study was supported in the field by the Colombian Navy ships ARC Malpelo, ARC Valle del Cauca and ARC Caldas, as well as by the ship María Patricia (especially H. Botero) and the ship Anita. Special gratitude to the advisory and support to the project provided by B. Werding and V. Wolters (JLU), D. Anderson, S. García and F. Estela. We are grateful to M. Rodríguez, P. Herrón, J. C. Botello and J. Zamudio for assistance in the field. The aerial photographs were provided through R. Pitman and F. Zapata by the Photogrammetry Group of NOAA, Southwest Fisheries Science Center, La Jolla, California. The satellite image was provided by the program ASTER of the NASA. T. Gottschalk and A. Misok (JLU), B. Posada, P. Lozano and C. Segura (INVEMAR) provided useful comments and technical support during the analysis of the information. We thank A. Franco and two anonymous reviewers for contributing with the improvement of the manuscript. M. Mendoza and A. Hiller kindly helped to write the English version of this paper.
LITERATURE CITED
1. Álvarez-Rebolledo, M. 2000. Aves de la Isla de Malpelo. Biota Colombiana, 1(2): 203-207. [ Links ]
2. Anderson, D. J. 1993. Masked Booby (Sula dactylatra). In: A. Poole and F. Gill (Eds.). The birds of North America, No. 73. The Academy of Natural Sciences, Philadelphia, PA, and The American Ornithologist's Union, Washington, D.C. [ Links ]
3. Bond, J. and R. Meyer de Schauensee. 1938. Zoological Results of the George Vanderbilt South Pacific Expedition of 1937. Part II,-The birds of Malpelo Island, Colombia. Proceedings of the Academy of Natural Sciences of Philadelphia,90: 155-157. [ Links ]
4. Brando, A., H. von Prahl and J. R. Cantera. 1992. Malpelo: Isla oceánica de Colombia. Banco de Occidente. Santiago de Cali. [ Links ]
5. Caita, C. and R. Guerrero. 2000. Geología de la isla Malpelo. Trabajo de Grado, Universidad Nacional de Colombia, Departamento de Geología. p 127, 5 anexos. [ Links ]
6. Chardine, J. W. and R. D. Morris. 1996. Brown Noddy (Anous stolidus). In: A. Poole and F. Gill (Eds.). The birds of North America, No. 220. The Academy of Natural Sciences, Philadelphia, PA, and The American Ornithologist's Union, Washington, D.C. [ Links ]
7. Díaz, J. M., L. M. Barrios, M. H. Cendales, J. Garzón-Ferreira, J. Geister, M. López-Victoria, G. H. Ospina, F. Parra-Velandia, J. Pinzón, B. Vargas-Angel, F. A. Zapata and S. Zea. 2000. Áreas Coralinas de Colombia. Invemar, Serie Publicaciones Especiales 5, Santa Marta. [ Links ]
8. Duffy, D. C. 1984. Nest site selection by Masked and Blue-footed boobies on Isla Española, Galápagos. The Condor, 86: 301-304. [ Links ]
9. Felicísimo-Pérez, A. M. 1994. Modelos digitales del terreno: principios y aplicaciones en las ciencias ambientales. Pentalfa Ediciones. [ Links ]
10. Gauger, V. H. 1999. Black Noddy (Anous minutus). In: A. Poole and F. Gill (Eds.). The birds of North America, No. 412. The Academy of Natural Sciences, Philadelphia, PA, and The American Ornithologist's Union, Washington, D.C. [ Links ]
11. Gottschalk, T. K., F. Huettmann and M. Ehlers. 2005. Thirty Years of Analysing and Modelling Avian Habitat Relationships using Satellite Imagery Data: A Review. Int. J. Remote Sens., 26: 2631-2656. [ Links ]
12. Graham, J.B. (ed.). 1975. The biological investigation of Malpelo Island, Colombia. Smithsonian Contribution to Zoology, 176. [ Links ]
13. Harris, M. P. 1970. Breeding ecology of the Swallow-tailed Gull, Creagrus furcatus. The Auk, 87(2): 215-243. [ Links ]
14. Kiester, A. R. and J. A. Hoffman. 1975. Reconnaissance and mapping of Malpelo Island. Smithsonian Contribution to Zoology, 176: 13-16. [ Links ]
15. López-Victoria, M. and F. Estela. 2006. Aspectos sobre la ecología del piquero de Nazca (Sula granti) en el SFF Malpelo. 140-156 pp. In: Castrillón-Galvez, A. and B. S. Beltrán-León (Eds.). Santuario de Fauna y Flora Malpelo: Descubrimiento en marcha. UAESPNN - CCCP. Bogotá, 156 p. [ Links ]
16. Murphy, R. C. 1945. Island contrasts. Natural History, 15: 14-23. [ Links ]
17. Nelson, J. B. 1978. The Sulidae: gannets and boobies. Oxford Univ. Press, Oxford. 500 p. [ Links ]
18. Niethammer, K. R. and L. B. Patrick. 1998. White Tern (Gygis alba). In: A. Poole and F. Gill (Eds.). The birds of North America, No. 371. The Academy of Natural Sciences, Philadelphia, PA, and The American Ornithologist's Union, Washington, D.C. [ Links ]
19. Pitman, R. L., L. B. Spear and M. P. Force. 1995. The marine birds of Malpelo Island, Colombia. Colonial Waterbirds 18: 113-119. [ Links ]
20. Pitman, R. L. and J. R. Jehl (Jr.). 1998. Geographic variation and reassessment of species limits in the "Masked" Boobies of the Eastern Pacific Ocean. Wilson Bull., 110(2): 155-170. [ Links ]
21. Prahl, H. von. 1990. Malpelo la roca viviente. FEN Colombia, Editorial Presencia, Bogotá, Colombia. 57 p. [ Links ]
22. Sarmiento, A. 1953. Comisión a la isla de Malpelo. Boletín Geológico, 1(3): 40-57. [ Links ]
23. Schreiber, E. A., R. W. Schreiber and G. A. Schenk. 1996. Red-footed Booby (Sula sula). In: A. Poole and F. Gill (Eds.). The birds of North America, No. 241. The Academy of Natural Sciences, Philadelphia, PA, and The American Ornithologist's Union, Washington, D.C. [ Links ]
24. Slevin, J. R. 1928. Description of a new species of lizard from Malpelo Island. Proceedings of the California Academy of Sciences, 16(21): 681-684. [ Links ]
25. Stead, J. A. 1975. Field observations on the geology of Malpelo Island. Smithsonian Contribution to Zoology, 176: 17-20. [ Links ]
26. Townsend, C. H. 1895. Birds from Cocos and Malpelo Islands, with notes on petrels obatined at sea. Bulletin of the Museum of Comparative Zoology at Harvard College, 27(8): 121-126. [ Links ]
27. Townsend, M. T., K. P. Huyvaert, P. J. Hodum and D. J. Anderson. 2002. Nesting distribution of Galápagos boobies (Aves: Sulidae) an apparent case of amensalism. Oecologia, 132: 419-427. [ Links ]
DATE RECEIVED: 18/05/06 DATE ACCEPTED: 02/10/06
