










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.36 no.1 Santa Marta Jan./Dec. 2007
VARIACIÓN ESPACIAL Y TEMPORAL DE JUVENILES DE PINCTADA IMBRICATA (RÖDING, 1798) Y ARGOPECTEN NUCLEUS (BORN, 1778) EN LA REGIÓN NORTE DEL CARIBE COLOMBIANO
SPATIAL AND TEMPORARY VARIATION OF JUVENILES OF PINCTADA IMBRICATA (RÖDING, 1798) AND ARGOPECTEN NUCLEUS (BORN, 1778) IN THE NORTH OF THE COLOMBIAN CARIBBEAN
Claudia Castellanos1,2 y Néstor H. Campos2
1Instituto de Investigaciones Marinas y Costeras (InveMar), Cerro Punta Betín, Santa Marta, Colombia.
2Universidad nacional de Colombia, Instituto de Ciencias naturales (ICn), Santa Marta, Colombia. e-mail: claudiac@invemar.org.co (CC); nhcampos@invemar.org.co (nHC)
RESUMEN
Con el propósito de conocer la respuesta de juveniles de bivalvos frente a los cambios en la condición hidrográfica, fue evaluada la variación espacial (regiones, bahías y profundidades) y temporal (períodos climáticos) de la abundancia de Pinctada imbricata y Argopecten nucleus en la región norte del Caribe colombiano. Esta variación fue determinada en dos fases, a partir de la fijación de los individuos en colectores artificiales. La primera fase, entre 1994 y 1996 fue realizada en el Parque Nacional Natural Tayrona (PNNT) donde se comparó la abundancia de juveniles en Bahía Concha y Bahía Chengue a 5, 10, 15 y 20 m de profundidad; fueron establecidas diferencias entre los factores considerados; la mayor abundancia promedio a 5 m para Pinctada imbricata (145.3 +/- 91.7 individuos por colector) fue obtenida en Chengue y se colectó mayor número de individuos durante el período seco, caracterizado por la presencia de aguas de surgencia. Argopecten nucleus presentó menor abundancia (10.1+/-4.9 ind/col) y sobresalió igualmente en Chengue a 5 m. En la segunda fase, entre 1997 y 1998, fue comparada la abundancia de juveniles en el PNNT y el Cabo de la Vela (CV); la abundancia promedio de P. imbricata (60.9+/-15.7 ind/col) fue mayor en el PNNT y con más dominancia en el período seco del año; A. nucleus (22.9+/-4.6 ind/col) fue más abundante en el CV durante el mismo período. En el PNNT, la abundancia de P. imbricata estuvo relacionada con la temperatura; en el caso de A. nucleus fue evidenciada cierta relación con la visibilidad en el CV. Aunque no se encontró una clara dependencia de la abundancia de juveniles con los factores ambientales considerados, el mayor número de juveniles fue observado en un período climático específico, donde las variables ambientales poseen características definidas.
PALABRAS CLAVE: abundancia, juveniles, bivalvos, Caribe colombiano.
ABSTRACT
In order to know the response of this group of organisms to changes in hydrographic conditions, spatial (regions, bays, depths) and temporal (climatic periods) variation in the juveniles abundance of two species with potential economical use in the northern Colombian Caribbean region was assessed. This variation was determined from 1994 to 1996, by the fixation of individual in artificial collectors in the Tayrona National Park (PNNT) and comparing the abundance of juveniles in Concha Bay and Chengue bay at depths of 5, 10, 15 and 20 m. Differences between bays and depths were found; Chengue Bay at 5 m had the highest average abundance of Pictada imbricata (145 individuals per collector). Differences between climatic periods were also registered; for the first species the highest number of individuals were collected during the dry period –first semester of the year, which is characterized by the presence of upwelling waters. The abundance of Argopecten nucleus had a lower abundance and an undefined recruitment pattern. From 1997 to 1998 the abundance of juveniles between the PNNT and La Vela Cape (CV) was compared finding significant differences between these regions; the average abundance of P. imbricata (61 ind/col), was greater in the PNNT while individuals of A. nucleus (23 ind/col) were more abundant in the CV. At the temporal scale, these species presented differences in both regions. The first two exhibited a defined pattern in the PNNT with more dominance in the dry period of the year; A. nucleus showed great variation in the PNNT while in the CV they were dominant in the dry period. According to these results, the optimal conditions for recruitment of A. nucleus are given in the CV region during the dry period, and P. imbricata in the region of PNNT during the dry period. The temperature was related with abundance of P. imbricata en el CV and A. nucleus with visibility. Nevertheless, the highest number of juveniles was observed during a specific climatic period in which the environmental variables have well defined characteristics.
KEY WORDS: abundance, juveniles, bivalves, Colombian Caribbean.
INTRODUCCIÓN
Los primeros estadios larvales de los bivalvos son de vida planctónica y luego aparecen estructuras especializadas que inducen a la búsqueda de un sustrato adecuado de asentamiento; en este proceso se presentan alteraciones morfológicas y fisiológicas en el cual ocurren altas mortalidades e intervienen factores físicos y biológicos interrelacionados que definen los ciclos dentro de las poblaciones (Crisp, 1974; Sastry, 1979; Bonar et al., 1990). Actualmente muchas especies de bivalvos son un recurso marino cultivable, que ofrece buenas perspectivas de comercialización; sin embargo, antes de plantear una estrategia para su manejo, es primordial comprender la dinámica de las primeras fases de vida, su distribución, temporada de mayor abundancia y las condiciones ambientales que permitan un óptimo desarrollo para predecir reclutamientos variables o definidos (Rho y Pyen, 1987; Wada, 1993). Trabajos enfocados en el reclutamiento determinaron los períodos y lugares adecuados para la obtención de estos organismos (Ruiz y Cáceres, 1990; Knuckey, 1995; Aguilar y Wolfgang, 2000; Rupp y De Bem, 2004) y encontraron fluctuaciones entre años sucesivos en la abundancia y la duración del período de mayor asentamiento, relacionado con actividad reproductiva de adultos y el comportamiento de los factores ambientales (Monteforte y Wright, 1994; Jiménez et al., 2000; Narvarte et al., 2001). En Colombia, trabajos sobre malacofauna se iniciaron en los años 70, enfocados al estudio taxonómico y ecológico (Perdomo, 1974; Torres, 1974). Los primeros estudios sobre su aprovechamiento se realizaron en la Ciénaga Grande de Santa Marta, con la captación de juveniles de Crassostrea rhizophorae en diferentes tipos de sustrato (Rivera, 1978; Wedler, 1980). Posteriormente, continuaron en el Golfo de Morrosquillo (Bernal y Mosquera, 1975; Ramírez y Salazar, 1977; Daza y Arias, 1994) y en el Parque Nacional Natural Tayrona se generó información sobre dinámica poblacional y asentamiento de Pteria colymbus (Ablanque, 1996), Pinna carnea (García-Valencia, 1997), Nodipecten nodosus y Euvola ziczac (Castellanos, 2006). En el Pacífico destacan los estudios sobre la piangua Anadara tuberculosa (Olave, 1985; Borrero, 1986) como de otras especies de bivalvos (Jaramillo, 2003). Las especies presentadas en este trabajo se distribuyen en el Atlántico occidental; Pinctada imbricata se caracteriza por la calidad del nácar, constituyéndose en un recurso importante para la producción de perlas (Sims, 1993). Desde el siglo XV se reportan extracciones de ostras, para obtención de este valioso material, actividad que ha diezmado con el tiempo, casi hasta la extinción de los bancos naturales (Monteforte y Cariño, 1992; Borrero et al., 1996); esta especie se adhiere a sustratos duros de aguas someras y en praderas de pastos marinos conforma nódulos. Las poblaciones naturales de Argopecten nucleus son de baja densidad y se encuentran en fondos arenosos y en zonas arrecifales hasta 50 m de profundidad (Díaz y Puyana, 1994). Con el fin de contribuir a disminuir el creciente deterioro y escasez de los recursos marinos por el uso de artes y métodos de pesca destructivos, la información generada en este trabajo es un aporte valioso para sentar bases que permitan realizar el cultivo de estos organismos, como una alternativa productiva para la comunidad de pescadores. De acuerdo a esto, el propósito del estudio fue evaluar los cambios espaciales y temporales en la abundancia de juveniles de Pinctada imbricata y Argopecten nucleus en la región norte del Caribe colombiano y determinar la influencia de factores ambientales en su reclutamiento.
ÁREA DE ESTUDIO
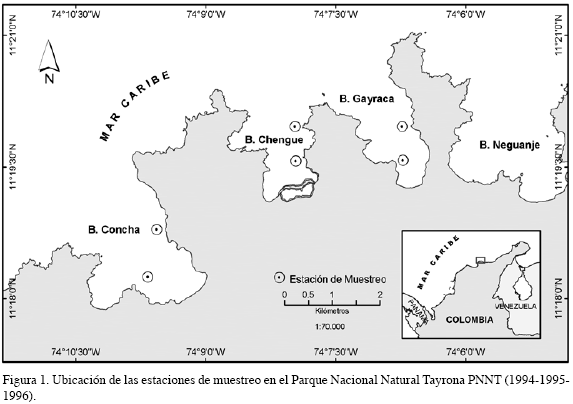
La variación espacial y temporal de juveniles de los bivalvos considerados fue estudiada en el Parque Nacional Natural Tayrona-PNNT (11°21’, 11°18 N’ y 74°6’, 74°10’ W) en el departamento del Magdalena y el Cabo de La Vela-CV (72° 20’ W y 12° 10’N) en el departamento de la Guajira. El PNNT localizado al noreste de la ciudad de Santa Marta, está conformado por las estribaciones de la Sierra Nevada de Santa Marta y se caracteriza por la presencia de acantilados, bahías pequeñas, variedad de ambientes marinos y por tener una plataforma muy estrecha (Garzón-Ferreira y Cano, 1991). La parte interna de las bahías presenta bosques de manglar, formaciones coralinas y praderas de pastos marinos. El CV presenta una línea litoral baja y homogénea, con amplias playas arenosas y lodosas, de escasa inclinación, que se prolongan mar adentro; la plataforma continental es extensa, alejándose de la costa más allá de los 40 km (Galofre y Martínez, 1990). Uno de los ecosistemas costeros predominantes en esta región son los pastos marinos; a lo largo de la franja costera del sector norte se localiza un banco natural de ostras perlíferas (Cuignon, 1987; Borrero et al., 1996). El clima de la región es seco, con una mayor condición de aridez en el CV; recibe la influencia directa de los vientos Alisios provenientes del nororiente, principalmente en el primer semestre del año, que crean una corriente con flujo este-oeste, paralelo a la costa y provocan un fenómeno estacional de surgencia. De esta forma, la capa de aguas superficiales es desplazada mar afuera y es reemplazada por masas de agua ascendentes de estratos profundos, caracterizadas por presentar baja temperatura, alta salinidad y un incremento de la producción fitoplanctónica, propias de aguas subtropicales localizadas entre 100-200 m (Fajardo, 1979; Corredor, 1981; Blanco, 1988). En el segundo semestre del año se incrementan la lluvias y la estabilidad del agua, que ocasiona un aumento de la temperatura y disminución de la salinidad; cuando la fuerza de los Alisios es menor, es contrarrestada por una contracorriente oeste-este, originada en el Caribe sur-occidental, que llega a influir especialmente hasta el PNNT (Ramírez, 1987; Díaz, 1990).
MATERIALES Y MÉTODOS
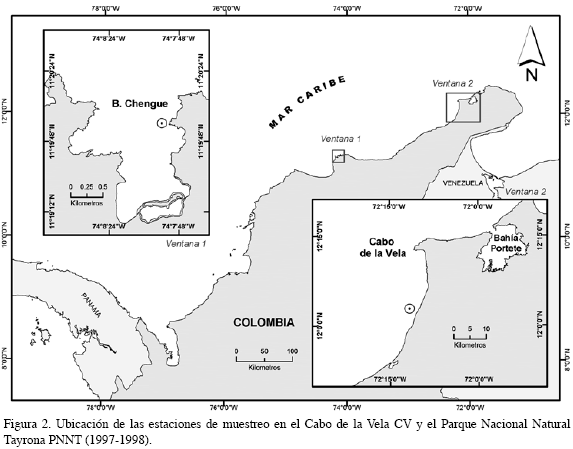
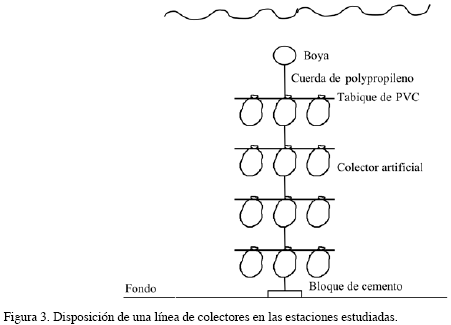
El trabajo fue realizado en dos fases de estudio. Para la primera fase, llevada a cabo en el PNNT entre 1994 y 1996, fue comparada la abundancia de juveniles entre bahías (Concha y Chengue), profundidades (5, 10, 15 y 20 m) y períodos climáticos (seco y lluvioso); cada periodo correspondió a un semestre del año, los impares, correspondieron al período seco y los pares, al lluvioso de cada año (Figura 1); en la segunda fase, entre 1997 y 1998, fue estudiada la abundancia entre regiones (PNNT y CV) e igualmente entre períodos (Figura 2). Un colector artificial fue utilizado como unidad de muestreo, constituido por dos bolsas elaboradas en malla de polipropileno, incluidas dentro de una bolsa de anjeo plástico (32 x 80 cm) con un tamaño de malla de 2 mm. Tres de estos colectores se sujetaron por tabiques de PVC en cada profundidad considerada, amarrados a un cabo suspendido en la columna de agua por medio de un bloque de cemento y una boya subsuperficial, que conformaron la línea de colectores (Figura 3); cada línea fue ubicada mensualmente en el agua. Con el fin de evaluar la variación espacial y temporal de los juveniles de las especies consideradas, fueron relacionados en la primera fase tres factores: período, bahía y profundidad. En la segunda fase, se relacionaron dos factores: región y período; para este caso fue seleccionada la información obtenida en los colectores dispuestos a 5 y 10 m en el PNNT con el fin de comparar la abundancia en el CV, en donde los colectores fueron ubicados a 3 y 6 m, región que se caracteriza por presentar una plataforma continental muy amplia y donde la profundidad máxima encontrada fue de 10 m. Con el propósito de determinar el efecto de los factores considerados en la abundancia de los individuos, fue realizado un ajuste de un modelo lineal generalizado de Poisson, que garantizará la inclusión de la sobredispersión de los datos en el mismo (Zar,
1996; Rico com pers); para este análisis se reunió la información obtenida en dos grupos: los semestres pares (periodo seco) y los impares (periodo lluvioso). Para determinar la influencia de las condiciones ambientales en la abundancia de juveniles, se midieron en forma simultanea a la permanencia de los colectores en el agua la temperatura del agua con un termómetro electrónico que registraba valores en forma continua; la salinidad con un conductímetro, visibilidad con un disco Secchi, contenido de materia orgánica e inorgánica particulada (seston) a partir de muestras de agua tomadas a 3 y 15 m (Bayne et al., 1985). Para relacionar la abundancia de los bivalvos con las variables ambientales medidas, se efectuó un análisis de correlación de Spearman.
RESULTADOS
Pinctada imbricata
Primera fase
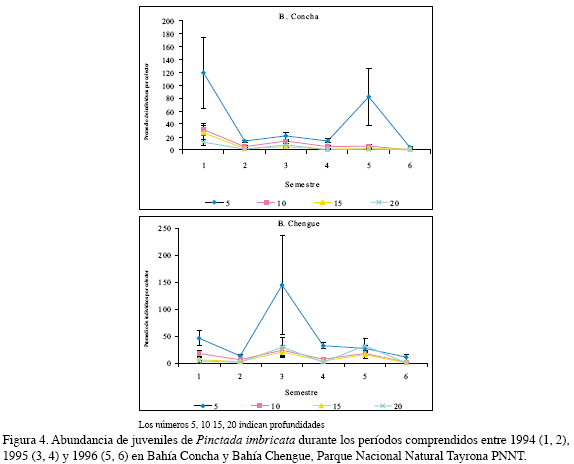
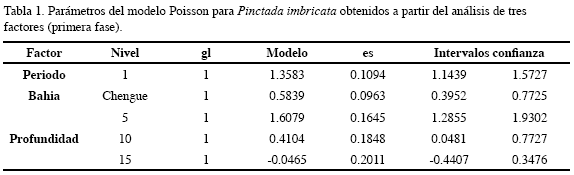
Esta especie presentó una variación similar en las dos bahías, destacando en el periodo seco, es decir en el primer, tercer y quinto semestres estudiados. En Concha fueron registrados valores promedio de 120.0±54.9 individuos por colector, durante el primer semestre y 81.9±43.2 ind/col en el quinto a 5 m; mientras que en Chengue fue más abundante en el tercero (145.3±91.7 ind/col) a la misma profundidad (Figura 4). De acuerdo al análisis por tres factores, el período para esta especie fue un factor significativo, mostrando así que la temporada climática afecta la recolección de los individuos, siendo en la época seca cuando se presentó una mayor abundancia en Chengue y la profundidad que mejores condiciones presentó para la recolección de juveniles de esta especie fue 5 m (Figura 5, Tabla 1).
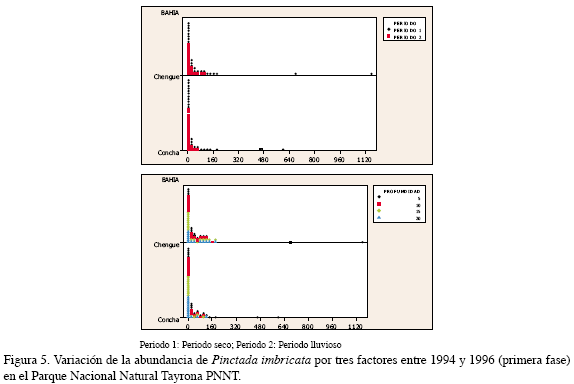
Segunda fase
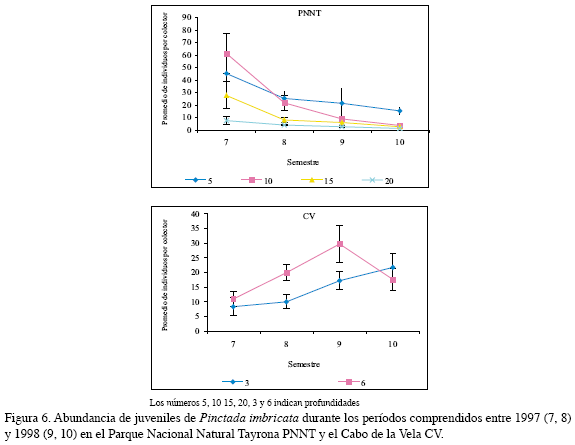
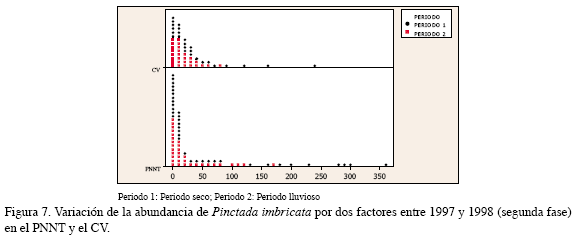
En el PNNT el mayor promedio de individuos de esta especie fue observado en el séptimo semestre a 10 y 5 m (60.9±15.7 ind/col y 45.0±23.5 ind/col, respectivamente); en el CV fue registrada una mayor abundancia a 6 m, especialmente durante el noveno semestre, con 29.6±6.2 ind/col. El análisis de dos factores constató que el período para esta especie fue un factor significativo; en el período seco fue obtenido el mayor número de individuos. De otra parte, la estimación del parámetro del modelo de Poisson mostró que en el período seco aumenta la probabilidad de encontrar una mayor cantidad de especimenes; por el contrario, el factor región no mostró un efecto significativo y la interacción tampoco afectó de manera directa a la abundancia (Figura 7, Tabla 2).

Argopecten nucleus
Primera fase
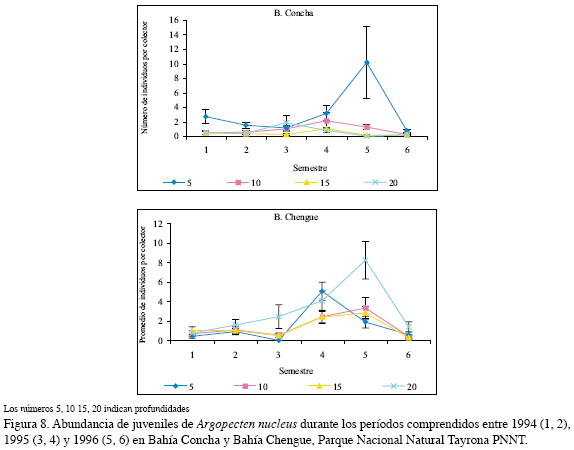
A. nucleus sobresalió en Concha en el quinto semestre (10.1±4.9 ind/col) a 5 m, mientras que en los demás semestres abarcados la abundancia fue similar y baja. En Chengue se observó mayor asentamiento en el quinto con 8.2 ind/col±1.9 a 20 m y en el cuarto semestre 5.0±0.9 a 5 m (Figura 8).
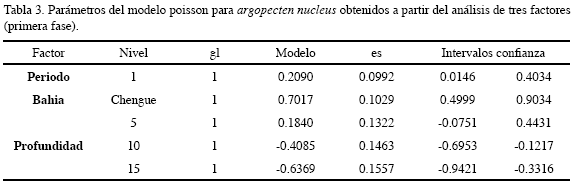
El análisis por tres factores arrojó que el comportamiento es en particular el menos marcado con respecto al periodo y la bahía en que se realice la labor de recolección de juveniles; en el período seco mostró un mayor incremento y la bahía Chengue estuvo por encima de Concha. La profundidad más apropiada para la obtención de juveniles resultó ser 5 m (Figura 9, Tabla 3).
Segunda fase
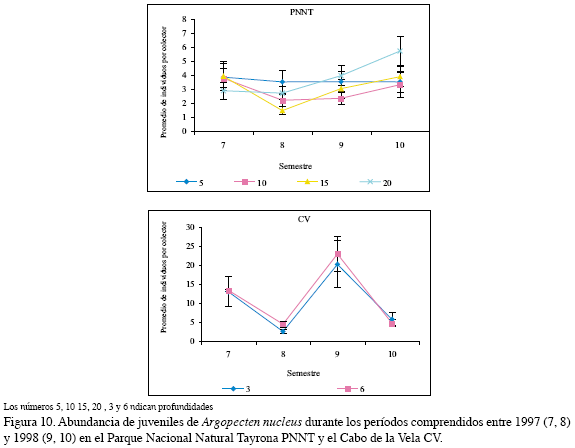
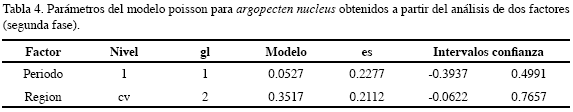
En el PNNT A. nucleus presentó una baja densidad de juveniles (5.7±1.0 ind/ col), mientras que en el CV fue mayor y sobresalió en el séptimo y noveno semestres, es decir, en el período seco (13.3±0.2 y 22.9±4.6 ind/col) (Figura 10). El análisis de dos factores mostró que las mejores condiciones para esta especie están dadas por la combinación período seco y la región CV, las posibilidades de obtener una mayor abundancia de esta especie se incrementaron en 1.2 veces en este período; con respecto al factor región, el CV fue la mejor ubicación para optimizar la captura de esta especie por cuanto las posibilidades se incrementan en 1.4 veces con respecto al PNNT, la otra localidad de estudio (Figura 11, Tabla 4).
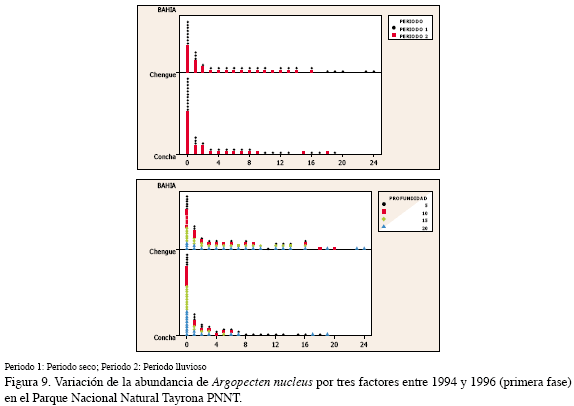
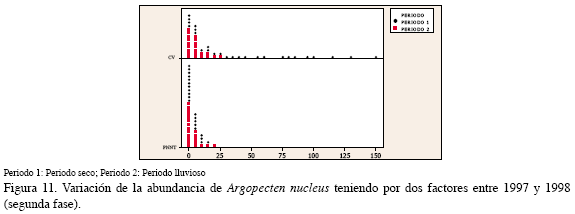
Al relacionar la abundancia de juveniles con las variables ambientales medidas, P. imbricata obtuvo una correlación negativa con la temperatura y con el seston en el PNNT (-0.49, p:0.01; -0.49, p:0.02); en el CV, A. nucleus, alcanzó un valor de asociación significativo con la visibilidad de -0.54 (p: 0.04); con las demás variables no se encontró una relación significativa.
DISCUSIÓN
Los juveniles de las especies estudiadas aparecieron en forma continua durante el estudio, con variaciones en la intensidad de individuos. Es conocido que los bivalvos presentan maduración permanente, con picos de mayor cantidad de individuos maduros (Gaytán et al. 1993; Monteforte et al., 1995; Knuckey, 1995). Assmus (2000), encontró para P. imbricata dos períodos de mayor frecuencia de adultos maduros en el Cabo de la Vela, en mayo y entre agosto y septiembre. El primero coincidió con el reporte de Perdomo (1974), para la región de Santa Marta. En el presente estudio, el asentamiento de juveniles de esta especie sobresalió en el primer semestre del año, cuando el clima es seco, con presencia de surgencia y coincidiendo con una de las temporadas de mayor aparición de individuos maduros. En los bancos naturales del nororiente de Venezuela, León et al. (1987) reportó P. imbricata, especialmente hacia final y comienzo del año (septiembre y enero); Jiménez et al. (2000) registró un pico de asentamiento coincidente con el obtenido en este estudio, a finales del período seco, y otro a finales del período lluvioso.
Individuos adultos de A. nucleus, acondicionados en laboratorio, expulsan los gametos de manera parcial, es decir, que una maduración continua también es típica de esta especie (INVEMAR, 1999). En Venezuela, Lodeiros et al. (1993) observaron el mismo patrón; en México y Panamá fue demostrado que Argopecten circularis exhibió una mayor liberación de gametos durante dos períodos, el primero registrado al final del período seco y el segundo al inicio del lluvioso (Gómez y Villalaz, 1988). En el PNNT fue encontrado bajo número de individuos de A. nucleus, hecho que no permite dilucidar un patrón de reclutamiento de la especie en esta región, que fue evidente en el CV, al predominar en el período seco. En cercanías a esta región última existen reportes de poblaciones naturales y observaciones de conchas fragmentadas de A. nucleus (Rachello, 2003) en cercanías al CV.
La variabilidad en la abundancia de juveniles de las dos especies estudiadas por región fue evidente. En el PNNT, además de presentar un patrón definido de asentamiento, P. imbricata mostró un mayor número de juveniles; por el contrario, A. nucleus predominó en el CV, con preferencia por asentarse en mayor número durante el período seco del año.
La morfología de las bahías del PNNT, protegidas, con retención y recirculación de agua hacia sus costados (INVEMAR, 1995), permitió aumentar el flujo a través de las bolsas colectoras, admitiendo un mayor asentamiento de juveniles de P. imbricata en los colectores dispuestos en el área. En las dos regiones estudiadas se conoce de la existencia de adultos de P. imbricata (Perdomo, 1974; Assmuss, 2000). En el CV se encuentran bancos naturales de esta especie; sin embargo, han sido diezmados con el tiempo (Borrero et al., 1996), y en el PNNT su densidad poblacional es menor. Por la presencia de estos bancos, se esperaba obtener una cantidad apreciable de individuos en el CV, sin embargo las corrientes inducidas por el viento influyen en el transporte de masas de agua e intervienen en el poder de dispersión de las larvas (Criales, 2000), lo que provoca baja retención de juveniles en el sistema donde se encuentran los parentales.
El CV, caracterizado por la exposición fuerte a la acción del oleaje, no influyó para que A. nucleus apareciera en los sistemas de captación, además se ve favorecido por la presencia de adultos en el área; por la circulación de las masas de agua, en sentido noreste-suroeste, es probable que sus poblaciones se encuentren en la parta alta de la Guajira, o cerca de las costas venezolanas; en estas últimas se tienen reportes de la existencia de poblaciones naturales (Lodeiros et al., 1993).
La preferencia que mostraron los juveniles en el PNNT, por asentarse en los primeros metros de la columna de agua, ocurrió, posiblemente, por las migraciones verticales que realizan las larvas en búsqueda de alimento o escapando de algún depredador. En el caso de P. imbricata fue evidente una preferencia para asentarse a 5 m. Ruiz y Cáceres (1990) y Monteforte y Wright (1994) reportaron un comportamiento semejante para Pinctada mazatlanica. A partir de mediciones de la concentración de seston y la biomasa fitoplanctónica a 8 y 21 m. Jiménez et al. (2000) demostraron la preferencia de P. imbricata por sitios someros que correspondieron con los mayores valores de las variables medidas. En este estudio, juveniles de A. nucleus fueron encontrados en mayor cantidad a 5 m y, en algunos casos, también destacaron a 20 m; sin embargo, se determinó preferencia por la primera profundidad. El patrón batimétrico difiere para otras especies de pectinidos, como Argopecten circularis, A. Gibbus y Pecten maximus, las cuales prefieren asentarse en profundidades mayores a 10 m o cerca al fondo marino (Coleman, 1988; Thouzeau, 1991; Avendaño y Cantillanez, 1992; Young et al., 1992).
La baja abundancia de juveniles obtenida, puede deberse a que otros fenómenos influyan (Lodeiros et al., 1993); las condiciones oligotróficas limita el asentamiento de las larvas, que necesitan almacenar mucha energía lipídica para poder realizar la metamorfosis (Thorson, 1964). La productividad de la surgencia presente en el norte del Caribe colombiano es comparativamente mucho menor (190 mgC/m2.h) a la observada en otras regiones de surgencia como Perú (1000 mgC/m2.h) (Corredor, 1992), donde se encuentran grandes bancos naturales de Argopecten purpuratus que permiten soportar cultivos comerciales de esta especie, a partir de la captación de juveniles provenientes del medio natural.
Por otro lado, el reclutamiento natural de P. imbricata y A. nucleus en la región resulta en bajas densidades poblacionales, situación que bien podría ser ocasionada por una dispersión masiva de sus larvas; conforme completan su desarrollo larval, las corrientes las transportan cientos de kilómetros del sitio de desove y definen los lugares y la distancia de dispersión (Cranfield, 1973; Sigurdson et al., 1976; Vélez y Azuaje, 1993); sumando a esto una actividad de extracción pesquera de los adultos.
Las variables ambientales están relacionadas con los procesos fisiológicos de los organismos y sus fluctuaciones influyen en la dispersión y abundancia de las especies (Dame, 1996); por la complejidad del ambiente en que se desarrollan, la respuesta de las larvas obedece a un conjunto de interrelaciones (Hadfield, 1986). En este estudio, la variable relacionada con el asentamiento de juveniles de P. imbricata fue la temperatura, especie que se favoreció en presencia de aguas frías. A. nucleus predominó en aguas turbias, característico en la región del CV. Las demás variables medidas no fueron determinantes en la aparición de los bivalvos; la falta de asociación de abundancias de juveniles con variables ambientales medidas en este trabajo, se ha observado en otras regiones del Caribe (Manzoni, 1994; León et al., 1987). Lo anterior evidencia que la temperatura y el alimento influyen en mayor medida en eventos previos al asentamiento; una mayor disponibilidad de alimento es esencial para que el individuo realice la gametogénesis y madure sus gónadas, porque necesitan acumular energía durante ese proceso, además de asegurar una buena fuente de alimentación para las larvas, mientras que un aumento paulatino de la temperatura desencadena el desove (Loosanoff y Tommers, 1948; Bayne y Newell, 1983, citado por García-Domínguez et al., 1996; Narváez et al., 2000). En el área de estudio, Assmuss (2000) halló relación entre los procesos de maduración y desove de P. imbricata con estas variables ambientales.
Otros factores, no medidos en el estudio, influyen en mayor medida en el proceso de asentamiento; la habilidad de los individuos en su etapa larval para seleccionar en forma gregaria un sustrato y fijarse, podría también responder a estímulos externos de naturaleza biológica o química, como la presencia de sustancias biológicamente activas (feromonas), que sirven para comunicarse entre las larvas durante el proceso de fijación (Morse et al., 1979; Weiner et al., 1989, en Tapia, 1994); de igual manera, aminoácidos excretados por algas y microorganismos son percibidos por las larvas en la zona circundante que les permite encontrar el sustrato adecuado (Pacheco, 2002). En el colector, además de las especies de interés, se obtuvo una gran cantidad de fauna asociada, como otras especies de bivalvos, con mayor probabilidad de ocupar el nuevo sustrato, antes que las especies de interés y especies de gasterópodos y crustáceos, conocidos por ser depredadores de moluscos (Castellanos, 2006).
La oferta de juveniles en los sistemas de captación fue continua; sin embargo, en el período seco se encontraron las mejores condiciones para el asentamiento de juveniles de las especies estudiadas y la profundidad en la que se asentó el mayor número de individuos correspondió a 5 m. Pinctada imbricata presentó mayor abundancia en el PNNT y el escenario óptimo para A. nucleus fue dado en el CV. El factor ambiental que más incidió en la abundancia de juveniles de P. imbricata fue la temperatura, mientras que para A. nucleus fue evidenciada cierta relación con la visibilidad.
AGRADECIMIENTOS
Los autores expresan su agradecimiento para la realización de este trabajo al Instituto de Investigaciones Marinas y Costeras “Jose Benito Vives de Andreis”. A Adriana Valero por el apoyo en la toma de datos y a Jimmy Rico por la asesoría en el manejo de la información. Este artículo hace parte de la tesis titulada “Variación espacio-temporal de la abundancia de juveniles de bivalvos y factores ambientales que la controlan en la región norte del Caribe colombiano” presentada en la Universidad Nacional de Colombia, como requisito para optar al título de M. Sci.
BIBLIOGRAFÍA
1 Ablanque, M. 1996. Ecología de la ostra perlífera Pteria colymbus incluyendo biología reproductiva y crecimiento de poblaciones naturales. Tesis (Biólogo) Pontificia Universidad Javeriana. Santa Marta, 107 p. [ Links ]
2 Aguilar, M. y B. Wolfgang. 2000. Settlement sites of juvenile scallops Argopecten purpuratus in the subtidal zone al Puerto Aldea, Tongoy Bay Chile. J. Shellf. Res., 19(2):749-755. [ Links ]
3 Assmus, J.P. 2000. Dinámica poblacional de la ostra perlífera Pinctada imbricata (Röding, 1798) (Bivalvia: Pteriidae) en el Cabo de la Vela, Guajira, Caribe colombiano. Tesis Pontificia Universidad Javeriana. Departamento de Ciencias. Santafe de Bogotá. [ Links ]
4 Avendaño, M. y M. Cantillanez. 1992. Colecta artificial de semilla de Argopecten purpuratus (Lamarck, 1819) en Bahía de Mejillones, Chile II. Observaciones sobre profundidad óptima de captación. Estud. Oceanol., 11:39-43. [ Links ]
5 Bayne, B.L., D.A. Brown, K. Burns; D.R. Dixon; A. Ivanovici; D.R. Livingstone; D.M. Lowe; M.N. Moore; A.R. Stebbing y J. Widdows. 1985. The effects of stress and pollution on marine animals. Praeger Scientific Publications. New York. 384 p. [ Links ]
6 Bernal, M. y C. Mosquera. 1975. Bioecología y pesquería de la ostra Crassostrea rhizophorae Guilding en la Bahía de Barbacoas. Tesis (Biólogo marino). Universidad Jorge Tadeo Lozano. Facultad de Ciencias del Mar. 71 p. [ Links ]
7 Blanco, J. 1988. Las variaciones ambientales estacionales en las aguas costeras y su importancia para la pesca en la región de Santa Marta, Caribe colombiano. Tesis de Maestría (Biología Marina). Universidad Nacional de Colombia. Santa Marta. 50 p. [ Links ]
8 Bonar, D., S. Coon, M. Walch, R. Weiner y W. Fitt. 1990. Control of oyster settlement and metamorphosis by endogenous and exogenous chemical cues. Bull. Mar. Sci., 46:484-498. [ Links ]
9 Borrero, F. 1986. The collection of early juveniles of Anadara spp. as a potential source for culturing mangrove cockles in the pacific coast of Colombia. Aquaculture, 59:61-66. [ Links ]
10 Borrero, F., J.M. Díaz y A. Seczon 1996. Las ostras perlíferas (Bivalvia: Pteriidae) en el Caribe colombiano: Historia de su explotación, ecología y perspectivas para su aprovechamiento. Serie Publicaciones Especiales de INVEMAR. 1. 53 p. [ Links ]
11 Castellanos, C. 2006. Variación espacial y temporal de la abundancia de juveniles de bivalvos en la región norte del Caribe colombiano. Tesis (Maestría) Universidad Nacional de Colombia. Santa Marta. 106 p. [ Links ]
12 Coleman, N. 1988. Spat catches as indication of recruitment to scallops population in Victorian waters. 51- 60. En: Zacharin, J. (Ed). Proceedings of the australiasian scallop workshop, Dredge, Hobart, Australia. [ Links ]
13 Corredor, J. E. 1981. Apuntes sobre la circulación costera en el Caribe noroccidental colombiano. Bol. Cient. CIOH, 3:3-8. [ Links ]
14 Corredor, J. 1992. Dinámica espacio-temporal de la surgencia de la Guajira. En: Seminario Nacional de Ciencias y Tecnologías del Mar y Congreso Centroamericano y del Caribe en Ciencias del Mar. Santa Marta. 1:206-221. [ Links ]
15 Cranfield, H.J. 1973. Observations on the behaviour of the pediveliger of Ostrea edulis during attachment and cementing. Mar. Biol., 22:203-209. [ Links ]
16 Criales, M. 2000. Patrones de reclutamiento y mecanismos de transporte costero de estadios tempranos de desarrollo de peces y crustáceos decapodos en el área de Santa Marta, Caribe colombiano. Proyecto Código 2105-09-689-96. Informe Final. INVEMAR-Universidad de Miami. 195 p. [ Links ]
17 Crisp, D. 1974. Factors influencing the settlement of marine invertebrate larvae. 177-265. En: P.T. Gran y A.M. Mackie (Eds). Chemoreception in marine organisms Academic Press, London. [ Links ]
18 Cuignon, R. 1987. Estudio de la plataforma del Caribe colombiano, fase Guajira 1985. Bol. Cient. CIOH, 7:53-72. [ Links ]
19 Dame, R. 1996. Ecology of marine bivalves: an ecosystem approach. C.R.C. Mar. Sci. Ser. Florida. [ Links ]
20 Daza, P. y L.M. Arias. 1994. Evaluación preliminar del cultivo de ostra Crassostrea rhizophorae en el estuario de la Bahía de Cispatá. Cienaga de Mestizo y Pepino (San Antero, Córdoba). 38-49. En: Memorias primera reunión grupo de trabajo sobre cultivo de bivalvos en Colombia. INPA-CVS. Córdoba. [ Links ]
21 Díaz, J.M. 1990. Estudio ecológico integrado de la zona costera de Santa Marta y Parque Nacional Tayrona. Informe final. Programa ecosistemas marinos. INVEMAR, Santa Marta. 439 p. [ Links ]
22 Díaz, J.M. y M. Puyana. 1994. Moluscos del Caribe colombiano. Un catálogo ilustrado. COLCIENCIAS. Fundación NATURA-INVEMAR. 291 + il. [ Links ]
23 Fajardo, G. 1979. Surgencia costera en las proximidades de la península colombiana de la Guajira. Boletín Científico Centro de Investigaciones Oceanográficas e Hidrográficas, 2:7-19. [ Links ]
24 Galofre, R. y M. Martínez 1990. Dinámica poblacional del camarón rosado Penaeus (Farfantepenaeus) notialis Pérez Farfante, 1967 y del camarón pintado Penaeus (Farfantepenaeus) brasiliensis Latreille 1817 en la zona norte del Caribe colombiano. Tesis (Biología. marina) Universidad Jorge Tadeo Lozano. 67 p. [ Links ]
25 García-Valencia, C. 1997. Biología y ecología de la hacha Pinna carnea (Gmelin, 1791) (Mollusca, Bivalvia, Pinnidae) en la región de Santa Marta, Caribe colombiano. Tesis de grado. UJTL. Facultad de Biología Marina. Santa Marta. 103 p. [ Links ]
26 García-Dominguez, F., B.P. Ceballos-Vasquez and A. Tripp-Quezada. 1996. Spawning cycle of the pearl oyster, Pinctada mazatlanica (Hanley, 1856) (Pteriidae) at Isla Espiritu Santo, Baja California Sur, Mexico. J. Shellfish Res., 15(2):297-303. [ Links ]
27 Garzón-Ferreira, J. y M. Cano, 1991. Tipos, distribución, extensión y estado de conservación de los ecosistemas marinos costeros y del P.N.N.T. Manuscr. VII Concurso Nal. Ecol. FEN/INVEMAR, Bogotá. 82 p. [ Links ]
28 Gaytán-Mondragón I., C. Cáceres-Martínez y M. Tobías-Sánchez. 1993. Growth of the pearl oysters Pinctada mazatlanica and Pteria sterna in different culture structures at La Paz Bay, Baja California, Sur México. J. World Aquac. Soc., 24(40):541-546. [ Links ]
28 Gómez, J.A y J.R. Villalaz. 1988. Ciclo anual de maduración sexual de la conchuela Argopecten circularis. Scientia. Panamá, 3:67-77. [ Links ]
29 Hadfield, M.G. 1986. Settlement and recruitment of marine invertebrates: a perspective and some proposals. Bull. Mar. Sci., 19(2):418-425. [ Links ]
30 INVEMAR. 1995. Captación de postlarvas de invertebrados marinos en colectores artificiales y factibilidad de fuentes alternas de semilla para el desarrollo de la acuicultura de moluscos bivalvos en la región de Santa Marta. Informe final. Proyecto (Código 2105-09-028-93) COLCIENCIAS-INVEMAR. [ Links ]
31 INVEMAR. 1999. Diagnóstico y Evaluación de la factibilidad biológica, técnica y económica del cultivo experimental de moluscos bivalvos de interés comercial en el Caribe. Informe final INVEMAR. Proyecto BID- Centrum fur International Migration und Entwicklung-CIM de Alemania. Inv-Aid. Santa Marta. 273 p. [ Links ]
32 Jaramillo, S. 2003. Exploración del cultivo de moluscos bivalvos como alternativa productiva y alimentaria en la Costa Norte Chocoana. 14 p. En: Memorias del Seminario - Talleres - Muestra Industrial y Tecnología ACUICULTURA 2003 - alternativa alimentaría. Bogotá. [ Links ]
33 Jiménez, M., C. Lodeiros y B. Márquez. 2000. Captación de juveniles de la Madreperla Pinctada imbricata (Roding, 1798) con colectores artificiales en el Golfo de Cariaco, Venezuela. Carib. J. Sci., 36(3-4):221-226. [ Links ]
34 Knuckey, Y. 1995. Settlement of Pinctada imbricata (Jameson) and other bivalves on artificial collectors in the Timor Sea, Northern Australian. J. Shellf. Res., 14(2):411-416. [ Links ]
35 León, L.; T., Cabrera; L., Traccoli. 1987. Fijación e índice de engorde de la ostra perla Pinctada imbricata Roding 1798 (Mollusca: Bivalvia) en tres bancos naturales del nororiente de Venezuela. Aquac. Res., 35(4):385-396. [ Links ]
36 Lodeiros, C., L., Freites; M. Núñez y J. Himmelman. 1993. Growth of the Caribbean scallop Argopecten nucleus (Born, 1780) in suspended culture. J. Shellfish Res., 12("):291-294. [ Links ]
37 Loosanoff, V.L. y F.D., Tommers. 1948. Effect of suspended silt and other substances on rate of feeding of oysters. Science, 16:69-70. [ Links ]
38 Manzoni, C. 1994. Período reproductivo, asentamiento larval y crecimiento de Nodipecten nodosus (Linnaeus, 1758) (Mollusca: Bivalvia) en los alrededores de Isla Arvoredo (Santa Catarina-Brasil). Tesis de Maestría. CCA de Universidad de Santa Catarina. Florianapolis. [ Links ]
39 Monteforte, M. y M. Cariño. 1992. Exploration and evaluation of natural stocks Pinctada mazatlanica and Pteria sterna (Bivalvia: Pteriidae): La Paz, South Baja California, Mexico. Ambio., 21(4):314-320. [ Links ]
40 Monteforte, M. y H. Wright. 1994. Ecology of pearl oyster spat collection in Bahía de La Paz South Baja California, México: Temporal and vertical distribution, substrate selection, associated species. J. Shellf. Res., 13(1):342-343. [ Links ]
41 Monteforte, M., E. Kappelman-Piña y B. López-Espinosa. 1995. Spatfall of pearl oyster Pteria sterna (Gould) on experimental collectors at Bahía de La Paz, South Baja California, México. Aquac. Res., 26:497- 511. [ Links ]
42 Morse, D.E.; N., Hooker; H., Duncan y L., Jensen. 1979. Y-aminobutyric acid, a-neurotransmitter, induces planktonic abalone Larvae to settle and begin metamorphosis. Science, 204: 407-410. [ Links ]
43 Narváez, N.; C. Lodeiros; L. Freites; M. Núñez; D., Pico y A. Prieto. 2000. Abundancia de juveniles y crecimiento de Pinna carnea (Mytiloida: Pinnacea) en cultivo suspendido. Rev. Biol. Trop., 48(4): 785-797. [ Links ]
44 Narvarte, M.A., E. Félix-Pico y L.A. Ysla-Chee. 2001. Asentamiento larvario de pectínidos en colectores artificiales. 173-192. En: Maeda-Martínez, A.N. (Ed.). Los moluscos pectínidos de Iberoamérica: Ciencia y Acuacultura: McGraw-Hill. Mexico. [ Links ]
45 Olave, E. 1985. Recolección de los primeros estados de moluscos en la Bahía de Buenaventura con la ayuda de colectores artificiales. Tesis de grado (Biología-Zoología) Universidad del Valle, Cali. 145 p. [ Links ]
46 Pacheco, A. 2002. Reclutamiento y supervivencia de Argopecten purpuratus (Lamarck 1819), efectos de la disponibilidad y características del sustrato. Tesis de maestría. Facultad de Ciencias del Mar. Universidad Católica del Norte. Coquimbo. [ Links ]
47 Perdomo, C. 1974. Contribución al conocimiento bioecológico de la ostra perlífera Pinctada radiata (Leach, 1814) en la Bahía de Santa Marta. Tesis (Biología marina). Universidad Jorge Tadeo Lozano. Bogotá. [ Links ]
48 Rachello, P. 2003. Inventario y caracterización estructural de los moluscos de la plataforma continental del Caribe colombiano (20, 70 y 150 m). Tesis (Biología) Pontificia Universidad Javeriana. Facultad de Ciencias. 145 p. [ Links ]
49 Ramírez, G. 1987. Evaluación de los parámetros fisicoquímicos y su relación con la surgencia costera. En: Díaz. J.M. (ed.). Estudio integrado de la zona costera de Santa Marta y Parque Nacional Natural Tayrona. Informe final. Programa de Ecosistemas Marinos. INVEMAR. Santa Marta. 439 p. [ Links ]
50 Ramírez, M. y A. Salazar. 1977. Estudio preliminar sobre el cultivo artificial del ostión de mangle Crasssostrearhizophorae Guilding 1828 en la Bahía de Cispatá, Córdoba. Tesis (Biología marina). Universidad Jorge Tadeo Lozano, Bogota. 139 p. [ Links ]
51 Rho, S. y K. Pyen. 1987. Environmental studies on the culture and spat production of pearl oyster: Basic studies on mass spat production of the pearl oyster Pinctada fucata (Dunter). Bull. Mar. Res.. Inst. Cheju. Nat. Univ., 11:21-40. [ Links ]
52 Rivera, L.F. 1978. Experiencias en el cultivo de la ostra Crassostrea rhizophorae (Guilding, 1828) en la Ciénaga Grande de Santa Marta y estudio preliminar de la dinámica de su población. Tesis (Biología marina). Universidad Jorge Tadeo Lozano. Cartagena. 95 p. [ Links ]
53 Ruiz, C.A. y C. Cáceres. 1990. Estudio preliminar de captación de moluscos bivalvos en la Bahía de la Paz B.C. México. CICIMAR, 5(1):29-38. [ Links ]
54 Rupp,G.S. y M.M de Bem. 2004. Cultivo de Vieiras. 289-308. En: Poli, C. R., A.T.B Poli, E. Andreatta, & E. Beltrame (Eds.) Aquicultura: Experiências Brasileiras. Multitarefa Editora Ltda, Brasil. [ Links ]
55 Sastry, A.N. 1979. Pelecypoda (excluding ostreidae). 113-292. En: Giese AC, JS Pearse (Eds). Reproduction of marine invertebrates: molluscs. New York, Academic Press. [ Links ]
56 Sigurdson, J.B., C.W. Titman y P.A. Davies. 1976. The dispersal of young post-larval bivalve molluscs by byssus threads. Nature, 262:386-387. [ Links ]
57 Sims N. 1993. Size, age and growth of the black-lip pearl oyster, Pinctada margaritifera (Bivalvia; Pteriidae). J. Shellfish Res., 12 (2): 223-228. [ Links ]
58 Tapia, C. 1994. Respuesta al asentamiento y metamorfosis larval en Argopecten purpuratus (Lamarck 1819) inducidas por compuestos neuroactivos. Tesis de maestría. Facultad de Ciencias del Mar. Universidad Católica del Norte. Coquimbo.66 p. [ Links ]
59 Thorson, G. 1964. Some factors influencing the recruitment and establishment of marine benthic communities. Neth. J. Sea. Res., 3(2):267-293. [ Links ]
60 Thouzeau, G. 1991. Experimental collection of poslarvae of Pecten maximus and other benthic macrofaunal species in the Bay of Saint-Brieuc, France. II. Reproduction patterns and postlarval growth of five mollusk species. J. Exp. Mar. Biol. Ecol., 148:181-200. [ Links ]
61 Torres, M.A. 1974. Estudio biológico pesquero del scallop Pecten laurenti (Mollusca, Pelecypoda) en el Golfo de Morrosquillo. Tesis de (Biología marina) Universidad Jorge Tadeo Lozano. Bogotá. 94 p. [ Links ]
62 Vélez, A. y O. Azuaje. 1993. Efectos de algunos factores físicos sobre la fijación larval del mejillón Perna perna (Linnaeus). Bol. Inst. Oceanogr. Universidad de Oriente, 32(1 y 2):49-55. [ Links ]
63 Wada, K. 1993. Bivalve broodstock development in Japan. World Aquac., 24(3):54-57. [ Links ]
64 Wedler, E. 1980. Experimental spat collection and growing of the oyster, Crassostrea rhizophorae, Guilding, in the Ciénaga Grande de Santa Marta, Colombia. Aquaculture, 21:251-259. [ Links ]
65 Young, P.C., R.J. McLoughlin y R.B. Martin. 1992. Scallop (Pecten fumatus) settlement in Bass Strait, Australia. J. Shellf. Res., 11(2):315-323. [ Links ]
66 Zar, H. 1996. Biostatistical analysis. Third Edition. Prentice Hall. New Yersey, 662 p. [ Links ]
FECHA DE RECEPCIÓN: 05/06/06 FECHA DE ACEPTACIÓN: 03/09/07
