










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.37 no.2 Santa Marta July/Dec. 2008
DISTRIBUCIÓN DE ESPONJAS SOBRE LA PLATAFORMA CONTINENTAL DE LA GUAJIRA, CARIBE COLOMBIANO
DISTRIBUTION OF SPONGES ON THE LA GUAJIRA CONTINENTAL SHELF, COLOMBIAN CARIBBEAN
Michael Díaz1 y Sven Zea2
1Instituto de Investigaciones Marinas y Costeras - INVEMAR, Cerro Punta de Betín, Santa Marta, Colombia. biomichaeldiaz@yahoo.es
2Universidad Nacional de Colombia, Sede Caribe, Departamento de Biología y Centro de Estudios en Ciencias del Mar - CECIMAR. INVEMAR, Cerro Punta de Betín, Santa Marta, Colombia. szea@invemar.org.co
RESUMEN
Durante la exploración de los fondos blandos calcáreos de la plataforma continental del departamento de La Guajira se identificaron 47 especies de esponjas de las clases Demospongiae y Calcárea. A partir de arrastres de fondo a lo largo de la plataforma continental sobre las franjas batimétricas de 10 y 50 m, se encontró que el número de especies es mayor a 50 m de profundidad, principalmente en áreas al norte y sur del departamento donde resaltan sedimentos tipo arena-lodo. Sin embargo, estos esquemas no son consistentes en todos los casos de acuerdo con las bajas similitudes basadas en la presencia-ausencia de esponjas, indicando una composición faunística muy heterogénea. Los fondos blandos permiten el crecimiento de esponjas sésiles y no sésiles, debido a que se colectaron especies con crecimiento tipo masivo, incrustante, ramificado y esférico, todas sueltas pero aglutinando fragmentos grandes del sustrato sobre los que se apoyan.
PALABRAS CLAVE: Esponjas, Plataforma continental, La Guajira, Caribe, Distribución.
ABSTRACT
During the exploration of the soft calcareous bottoms of the continental shelf of La Guajira Department, 47 different species of sponges were identified in the classes Demospongiae and Calcarea. From bottom trawling carried out throughout the continental shelf on the bathymetric fringes of 10 and 50 m, it was found that the number of species is larger at 50 m depth, mainly in areas to the north and south where sand and mud sediments prevail. However, the patterns are not consistent in all the cases, owing to the low similarities between stations, based on the presence-absence of sponges, indicating a rather heterogeneous faunal composition. The soft bottoms allow the growth of non-sessile and sessile sponges; collected species show massive, incrusting ramified and spherical growth, all loosely attached but agglutinating large substratum fragments to which they hold.
KEY WORDS: Red seaweed, Pilot culture, Relative growth rates (RGR), Mariculture, Portete bay.
INTRODUCCIÓN
Las esponjas (phylum Porifera) son importantes componentes de las comunidades bentónicas tropicales. Las diversas estrategias de vida de las esponjas dentro de los ecosistemas marinos se relacionan con funciones ecológicas como incorporación de materia orgánica suspendida al fondo, producción primaria y remineralización (Díaz y Ward, 1997), conformación de hábitats para procariotes y eucariotes (Hentschel et al., 2002) y participación en la construcción de la estructura tridimensional del fondo (Reed y Pomponi, 1997). Estas funciones han sesgado su estudio ecológico hacia ecosistemas de fondos duros litorales (Alcolado, 1999; Zea, 2002; Bell y Barnes, 2003), ya que en fondos blandos generalmente se han estudiado para definir áreas biogeográficas (Van Soest, 1993; Worheide et al., 2005).
Sobre los fondos blandos someros de las plataformas continentales tropicales las comunidades de esponjas son menos diversas que las de fondos duros litorales, debido en parte al relieve homogéneo, la inestabilidad del sedimento y la baja disponibilidad de sustrato firme para apoyarse (Pansini y Musso, 1991; Ilan y Abelson, 1995). Se considera que sobre fondos blandos existen dos componentes o microhábitats del sustrato para el asentamiento de las esponjas. Por un lado el componente fino como arena y lodo para especies con hábitos de anclaje y por el otro el componente grueso, como cascajo, conchas de moluscos y exoesqueletos de crustáceos principalmente, para especies que requieren alguna forma de adhesión al sustrato (Van Soest, 1993; Rützler 1997). En fondos rocosos el relieve heterogéneo y la mayor estabilidad del sustrato permiten la presencia de gran número de especies de poríferos, que resaltan entre diversos invertebrados bentónicos, característicos de la fauna tropical marina (Parra-Velandia y Zea, 2003).
En Colombia más de la cuarta parte de las especies de esponjas han sido estudiadas en fondos duros litorales (Zea, 1987; Valderrama y Zea, 2003; entre otros). Concretamente para la península Guajira hay registros de esponjas en litorales rocosos someros (Zea, 1987), en praderas marinas de Thalassia testudinum (Montoya-Maya, 2002) y en sustratos artificiales como las plataformas marinas de extracción de hidrocarburos (INVEMAR, 2004). No se tiene información sobre esponjas en las amplias y dominantes extensiones de fondos blandos carbonatados de la plataforma continental de la península.
Los patrones de abundancia y distribución de las poblaciones de esponjas en un área se explican por la interacción de varios factores (Zea, 1993, 2002) siguiendo los gradientes de profundidad (Alcolado, 1999), sedimento (Pansini y Musso, 1991), estabilidad del fondo (Bell y Barnes, 2003), entre otros. En la plataforma continental de La Guajira el gradiente de profundidad es el factor que mejor explica la distribución de la biota marina (INVEMAR-CORPOGUAJIRA, 2006); en el caso de las especies bentónicas, se cree que el tamaño del sedimento también podría explicar como se distribuye la macrofauna marina en el área.
El principal objetivo del presente trabajo es mostrar y discutir algunas particularidades de la distribución de esponjas marinas que habitan en la plataforma continental de la península Guajira, nor-oriente del Caribe colombiano. Al mismo tiempo, busca contribuir al entendimiento del papel que pueda estar jugando este grupo de la macrofauna dentro de los sistemas tropicales.
ÁREA DE ESTUDIO
La plataforma continental de la península Guajira se ubica al extremo norte del continente sur americano, en la región nor-oriental de Colombia en el mar Caribe. Sobre la plataforma continental, que finaliza cerca de los 80 m de profundidad, se presentan amplias extensiones de fondos blandos constituidos por arena y lodo cargados con abundante material carbonatado (Romero, 2006). En La Guajira se presenta un mosaico ambiental influenciado por los vientos Alisios; está compuesto por aguas someras bastante turbias, afectadas por núcleos de afloramiento de aguas profundas frías. En el mar de La Guajira existe una alta productividad biológica (Suzunaga et al., 1992) y una red trófica donde predomina el componente bentónico (Criales-Hernández et al., 2006).
En las costas de la península resaltan amplias extensiones de playa y estrechos litorales rocosos, e importantes áreas con manglares y pastos marinos. Dichos ecosistemas presentan atributos naturales específicos, los cuales definen regiones de gran interés tanto para el uso como para la conservación. De tal manera que hacia el norte de Riohacha se reconoce la ecorregión Guajira y hacia el sur la ecorregión Palomino (INVEMAR, 2000). Igualmente, de acuerdo con la geomorfología de la plataforma continental, Álvarez-León et al., (1995) dividen en tres sectores el departamento de La Guajira. En el sector geográfico alto (al nor-oriente del Cabo de la Vela) la plataforma es inclinada y predominan sedimentos tipo arena y lodo de origen bioclástico, que se mezclan al norte con las aguas estuarinas del Golfo de Venezuela. En el sector bajo (al sur-occidente del Santuario de Fauna y Flora Los Flamencos) la plataforma se hace inclinada por las estribaciones de la Sierra Nevada de Santa Marta con predominancia de sedimentos de arena y lodo litoclásticos. En el sector medio (entre el Santuario Los Flamencos y el Cabo de la Vela) la plataforma es ancha, poco inclinada y contiene sedimentos tipo arena bioclástica (Figura 1).
MÉTODOS
La exploración de la plataforma continental de La Guajira se realizó del 30 de marzo al 9 de abril de 2005 abarcando las franjas batimétricas de 10 y 50 m, del área comprendida entre Dibulla, río Palomino y hasta Castilletes, límite geográfico con Venezuela (73° 40'- 70° 30' W). Se ubicaron 26 estaciones de muestreo con base en cartas batimétricas; 13 de éstas a 10 m de profundidad y otras 13 a 50 m. Se recolectaron muestras biológicas mediante una red de arrastre demersal tipo semibalón que alcanza hasta 500 m de profundidad. Se realizaron dos arrastres por cada estación siendo efectivos 44 de los 46 arrastres. Se obtuvieron además muestras de sedimento con una draga Van Veen con capacidad para 500 g de sedimento; éstas fueron analizados por personal del INVEMAR, quienes generaron la información al respecto con la que cuenta este estudio.
En cubierta las esponjas fueron separadas por estaciones y guardadas en bolsas plásticas con etanol 70 % o formol 10 %. En el laboratorio se preservaron en etanol 70 %. Las identificaciones hasta el nivel de especie fueron realizadas a partir de montajes permanentes de espículas, fibras y cortes histológicos según la metodología de Zea (1987). Las claves taxonómicas empleadas para la clasificación de esponjas son las propuestas por Hooper y Van Soest (2002). El material estudiado fue depositado en el Museo Natural de Historia Marina de Colombia (MHNMC) en el INVEMAR, proyecto 1410, consecutivo del museo 31557, sección porífera (POR), intervalo del catálogo 1057-1147.
La distribución de esponjas fue analizada mediante método estadístico de análisis multivariado. El análisis se realizó a partir de la presencia y ausencia de especies, empleando el coeficiente de similaridad de Jaccard, la técnica análisis clúster y el algoritmo de promedio de grupos (Clarke y Warwick, 2001), de acuerdo con el software Palaeontological Statistic (PAST) versión 1.34. Con el dendograma que mejor representara las asociaciones, se determinó cuales estaciones se relacionaban consistentemente según el sector geográfico, la ecorregión, la profundidad y el tipo de sedimento.
RESULTADOS
Únicamente en 13 de las 26 estaciones muestreadas en la plataforma continental de La Guajira aparecieron poríferos (Figura 1). En total fueron identificadas 46 especies pertenecientes a la clase Demospongiae, mientras solo una especie pertenece a la clase Calcarea. Por otro lado, al agrupar las esponjas por órdenes, Hadromerida, Halichondrida y Poecilosclerida contienen el 50 % de especies. Otro 40 % de las especies se clasificaron en los órdenes Haplosclerida, Dyctioceratida, Astrophorida, Spirophorida y Verongida. Las Demospongiae agrupadas en los órdenes Homosclerophorida, Chondrosida, Dendroceratida, Agelasida, y el orden Clathrinida perteneciente a la clase Calcarea, contribuyeron con el restante 10 % de las especies (Tabla 1).
En La Guajira, el máximo número de especies en una sola estación (233) fue de 22, mientras una fue el mínimo (estación 201). La mayoría de esponjas identificadas presentaron distribuciones puntuales; entre ellas resaltan Negombo sp., Chondrilla sp., Aulospongus samariensis, Lissodendoryx sigmata, Rhaphidistia sp., Ancorina megastylifera, Geodia gibberosa, Hyatella cavernosa y Cynachyrella alloclada. Mientras que sólo cuatro especies tuvieron una amplia distribución sobre la plataforma continental, cada una presente en un mínimo de cuatro estaciones. Niphates erecta apareció en seis estaciones, Ircinia felix apareció en cinco estaciones, mientras Hymeniacidon caerulea y Dragmaxia undata lo hicieron en cuatro.
Distribución de las esponjas
Al comparar el número de esponjas presentes en el área de estudio, 41 especies se encontraron a 50 m, franja en donde siete de las 13 estaciones muestreadas tuvieron representantes. A los 10 m se identificaron 16 especies distribuidas en seis de las 13 estaciones. Sin embargo, ya que hubo estaciones en ambas franjas batimétricas donde no aparecieron esponjas, el esquema de número de especies vs. profundidad no fue consistente a lo largo de toda el área de estudio.
De acuerdo con el dendograma de la Figura 2 se observa en primer lugar que las semejanzas entre estaciones fueron en general bajas, ninguna superando el 50 % de similaridad, y la mayoría asemejándose en menos de un 10 %. El análisis confirma que la identidad de las especies tiende a ser diferente entre las franjas batimétricas de 10 y 50 m, debido a que el grupo 1 contiene cinco de las seis estaciones a 10 m y los grupos 2 y 3 las siete estaciones a 50 m. En efecto, únicamente las especies Spirastrella coccinea, D. undata, Callyspongia vaginalis, Xestospongia muta, Dysidea sp., Aplysina fulva, Igernella notabilis y N. erecta se presentaron en ambas franjas batimétricas. Por otra parte, comparando los grupos del dendograma de acuerdo con el sector geográfico o con la ecorregión a la que pertenecen las estaciones, no se encontraron semejanzas o diferencias consistentes para agrupar áreas de acuerdo con la composición de esponjas; sin embargo, resalta que del total de especies (46) tan solo siete: Placospongia intermedia, P. amaza, Aaptos aff. aaptos, Tethya sp., H. caerulea, I. felix y N. erecta, aparecieron a lo largo de los tres sectores geográficos; aunque D. undata, Amorphinopsis maza, Axinyssa flavolivescens y Agelas wiedenmayeri aparecieron tanto en el alto como en el bajo. Estas once especies muestran las mayores distribuciones latitudinales para esponjas de La Guajira, considerando que desde Punta Gallinas hasta la frontera con Venezuela no se presentaron esponjas (Figura 1) y que hacia las estribaciones de la Sierra Nevada de Santa Marta algunas especies son comunes (Zea, 1987). El mayor número de especies se presentó en el sector geográfico alto, donde aparecieron 31 especies, seguido por el sector bajo con 24 y en menor número el sector geográfico medio con 22 especies.
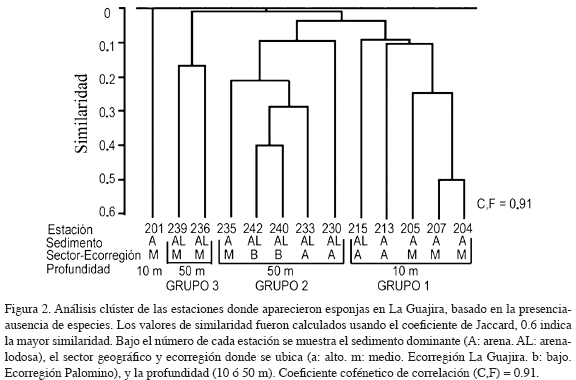
De igual forma, las estaciones no se agruparon de manera general según el tipo de sedimento y, aunque este factor tuvo cierta relación con la profundidad, esta no se refleja en la presencia de las especies de esponjas. En la plataforma de La Guajira se presentan tres tipos de sedimentos, arena, arena-lodo y lodo (Figura 1). En general, se puede decir que la distribución de los tipos de fondo se relaciona con la profundidad, más gruesos a 10 m y más finos a 50 m, aunque dentro de cada sector geográfico y en ambas profundidades los parches de fondos areno-lodosos generan un mosaico de sedimentos en algunas áreas. Cinco de las seis estaciones someras (grupo 1 y est. 201) estaban sobre fondo de arenas, en la restante (215) el sedimento era areno-lodoso. Mientras en seis de las siete estaciones profundas (grupos 2 y 3) el fondo era areno-lodoso y en la 235 arenoso. Sin embargo, resalta que ninguna de las especies de esponjas identificadas fue colectada en áreas con sedimentos exclusivamente tipo lodo. De esta manera, con excepción del lodo, que parece no favorecer la presencia de esponjas, la profundidad, más que el tipo de sedimento, determinó la composición de especies.
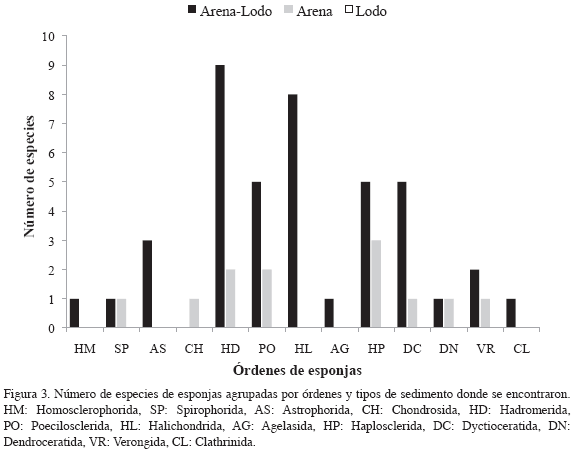
Interesantemente, las especies agrupadas en los órdenes Halichondrida, Astrophorida, Agelasida, Homosclerophorida y Clathrinida fueron recolectadas sobre fondos exclusivamente areno-lodosos. Algunas esponjas de los órdenes Hadromerida y Poecilosclerida también aparecieron sobre este tipo de fondo en la plataforma continental (Figura 3). De esta manera, según los grupos definidos en el dendograma y la sistemática de las especies identificadas (Tabla 1), se observa que las estaciones someras del grupo 1 presentaron pocas especies exclusivas (Cynachyrella kuenkenthali, A. samariensis, Desmapsamma anchorata, Phorbas amaranthus, Dragmacidon reticulata y Oceanapia nodosa). Mientras que las estaciones del grupo 2, todas profundas, presentaron numerosas especies exclusivas tales como Plakinastrella onkodes, C. alloclada, G. gibberosa, A. megastylifera, Cliona varians, P. intermedia, P. amaza, A. aff. aaptos, Tethya sp., Rhaphidistia sp., Cornulella santamartae, L. sigmata, lotrochota birotulata, Myrmekioderma gyroderma, Negombo sp., A. maza, Topsentia ophirhaphidites, A. wiedenmayeri, Xestospongia proxima, l. felix, Hyrtios violacea, H. cavernosa, Hyatella sp. 1, Aplisina cauliformis y Leucetta floridiana. Por su parte el grupo 3, representado por estaciones profundas, presentó especies únicamente colectadas en el sector medio, tales como Clathria adioristica, Haliclona sp. y Stelleta fibrosa var. globulariformis. La estación 201 se diferencia de las demás por la presencia exclusiva de Chondrilla sp.
Morfología de las esponjas
Al contrario de los fondos lodosos, los hábitats que generan los sedimentos predominantemente arenosos y carbonatados en las franjas batimétricas de 10 y 50 m, no parecen generar dificultades para el asentamiento y crecimiento de las esponjas. No obstante, debido a que la comunidad de especies colectadas en La Guajira no fue estudiada en su hábitat natural, las observaciones sobre la morfología externa sólo sugieren múltiples hábitos de crecimiento entre las especies, caracterizándose el de aglutinar, a medida que crecen, fragmentos grandes del medio como conchas y fragmentos bioclásticos (Figura 4). Esta característica fue común tanto para esponjas de crecimiento masivo, forma que presentaron la mayoría de especies en la península, como para las de crecimiento ramificado, esférico e incrustante. Sin embargo, entre los poríferos que presentaron estas dos ultimas formas de crecimiento resaltan especies como S. fibrosa var. globulariformis, C. varians y Tethya sp., las cuales presentaron individuos con forma esférica bien definida. Entre las incrustantes resaltan L. sigmata, P. melanos y Rhaphidistia sp., que se caracterizaron por crecer como masas amorfas que aglutinan en su ectosoma y endosoma partículas grandes del medio, adquiriendo así mayor consistencia física y aparentemente apoyándose sobre el fondo blando.

DISCUSIÓN
El número de especies de esponjas encontradas sobre la plataforma continental de La Guajira, antes del presente estudio, era el menor respecto a áreas del Caribe colombiano como Santa Marta, San Andrés, Islas del Rosario (Zea, 1987) y Urabá (Valderrama y Zea, 2003). Luego de la exploración de los fondos blandos de la península, el conocimiento de la fauna porífera que habita en La Guajira se cuadruplica, pasando de 12 (Zea, 1987; Montoya-Maya; 2002; INVEMAR, 2004) a 53 especies, la mayoría pertenecientes a órdenes de la clase Demospongiae (Díaz, 2007). Sin embargo, siguen siendo pocos los inventarios taxonómicos porífera en el área de estudio, especialmente hacia el talud continental.
Las comparaciones realizadas a partir de la presencia y ausencia de especies de esponjas en el área de estudio muestran bajas similaridades (< 0.5 coeficiente de Jaccard) y únicamente sustentan los agrupamientos de las 13 estaciones donde aparecieron poríferos como efecto de la profundidad (Figura 2). Esto sugiere que la composición de especies de esponjas que habitan en la plataforma varía a lo largo de dicho gradiente. Dicho esquema ocurre en fondos rocosos continentales e insulares del Caribe colombiano, donde a mayores profundidades las esponjas constituyen un componente dominante (Zea, 2002; Valderrama y Zea, 2003).
Por su parte, al comparar el número de especies por sectores geográficos o por ecorregiones, no se observa un claro gradiente, ni tampoco se sustentan las bajas similaridades detectadas entre las estaciones (Figura 2). Igualmente sucede con los estudios sobre la macrofauna móvil (INVEMAR-CORPOGUAJIRA, 2006) colectada en esta plataforma continental, con la que no se encontraron diferencias significativas entre ecorregiones, ni tampoco entre sectores geográficos. Dichos estudios también indican que organismos capturados como crustáceos, moluscos y peces, con los que sí se pudo hacer un análisis de estadística inferencial a partir de abundancias relativas e índices ecológicos, permiten proponer que, en áreas donde los sedimentos tipo arena-bioclástos-lodo predominan, los macro y microhábitats que se generan para la macrofauna contribuyen a que sectores geográficos como el alto y el bajo sean bastante diversos y contengan especies exclusivas habitando en áreas puntuales. Claros ejemplos constituyen las comunidades de corales azooxantelados estudiadas por Reyes et al. (2005) en bahía Portete al nor-oriente de la ecorregión Guajira y en inmediaciones de Palomino, Mingueo y Dibulla; donde también los llamados lechos de esponjas-briozoos-octocorales soportan la amplia diversidad de macrofauna identificada en la ecorregión Palomino (INVEMAR, 2005).
En la plataforma continental de La Guajira existen áreas puntuales y exclusivas donde se presentan numerosas especies, caso de la estación 233 localizada en el sector geográfico alto y de las estaciones 240 - 242 localizadas en el sector bajo (Tabla 1). Al mismo tiempo, es escaso el número de especies presentes en ambas franjas batimétricas (ocho especies), o las que muestran individuos al norte y sur de la plataforma continental (once especies). En consecuencia, la comunidad porífera que habita las aguas someras de la península no debe ser considerada como homogénea, debido a que la distribución de las especies no sigue un claro gradiente latitudinal. Dicho escenario se observa en la mayoría de fondos blandos donde las intervenciones generadas por la pesca comercial de arrastre, destruyen y casualmente transportan-reubican las esponjas que captura (Maldonado y Young, 1998; Kaiser et al., 2001). De esta manera, la distribución y abundancia de varias de las especies exclusivas de los grupos 1 y 2 (Tabla 1), podrían estar limitadas principalmente hacia el sector geográfico medio, por las intervenciones que genera la pesca comercial de arrastre. Este método de explotación es empleado desde hace décadas por las flotas pesqueras en el departamento de La Guajira (Zúñiga et al., 2004).
Sin embargo, la falta de homogeneidad en la distribución de las esponjas dentro del área de estudio, y en especial hacia el sector geográfico medio, también puede atribuirse a la disponibilidad de larvas y fragmentos que colonizan sustrato disponible en los fondos blandos. Al respecto, Pansini y Musso (1991) han propuesto que la composición por especies en los fondos blandos es producto de biocenosis vecinas que habitan en fondos duros. No obstante, contrario con lo discutido anteriormente, luego de la exploración realizada en La Guajira, 13 de las esponjas identificadas no han sido registradas en otra región del Caribe colombiano (Díaz, 2007); de esta manera, queda pendiente por definir si sobre esta plataforma continental, además de la exportación de larvas de fondos rocosos adyacentes como los de la Sierra Nevada de Santa Marta, y los presentes en el Golfo de Venezuela, la composición de la fauna de esponjas es sostenida por poblaciones fuente que habitan los mismos fondos blandos. En especial si consideramos que la subprovincia Guajira propuesta por Díaz (1995), a partir de la distribución de moluscos gasterópodos en el Caribe sur, debe ser evaluada con diferentes taxas marinos, entre ellos los de hábitos sésiles.
Los fondos blandos someros de la plataforma continental estudiada presentan arena y material calcáreo bioclástico de considerable tamaño, tal y como ocurre en la mayoría de plataformas continentales tropicales (Romero, 2006); sin embargo, hacia aguas profundas (> 80 m) en el sur de La Guajira y al norte, en el exterior del Golfo de Venezuela, predomina el lodo. Este tipo de fondo tan fino funciona como barrera que impide el establecimiento y el paso de larvas de especies con hábitos sedentarios marcados, fenómeno observado en diferentes latitudes (Van Soest, 1993). Por ello se podría postular de manera preliminar que la disponibilidad de hábitats para esponjas en la península es mayor en áreas donde se presentan fondos de arena-lodosa, de acuerdo con el número de especies identificadas en las estaciones ubicadas sobre estos mosaicos de sedimentos, con las escasas esponjas presentes únicamente sobre arena y con la ausencia de especies exclusivamente sobre lodo (Figura 3).
La disponibilidad de hábitats sobre fondo de arena-lodosa en aguas de la plataforma continental, se demuestra especialmente con cinco órdenes de esponjas, entre los que resaltan Hadromerida y Halichondrida, por el elevado número de especies que fueron colectadas sobre este tipo de fondo. Al indagar sobre que sustratos colonizan las esponjas a nivel de orden, la cantidad de estrategias sexuales (larvas vivíparas y ovíparas que sobreviven pocos días) y asexuales (gémulas y regeneración por fragmentos) junto con la enorme variabilidad de hábitos entre especies, no permiten establecer un claro escenario. Por lo que la información concluyente a este nivel es ambigua y no evidencia las particularidades que pueden estar favoreciendo la presencia de un mayor número de especies de un mismo orden en La Guajira. De cualquier forma, se hace necesario comparar la fauna de esponjas entre diferentes regiones, para establecer las relaciones biogeográficas y definir la relevancia de los nombres de especies con que cuenta la taxonomía de poríferos en Colombia.
Morfología de las esponjas
Ecosistemas con fondos semejantes a los de La Guajira son conspicuos en todos los mares, incluso donde se presentan comunidades arrecifales, praderas marinas y manglares (Kaiser et al., 2001). Los parches arenosos generan dificultades para el crecimiento de organismos sésiles, debido principalmente a la inestabilidad del sustrato, la acción de las tormentas sobre las corrientes que depositan los sedimentos en el fondo y las altas concentraciones de aminoácidos disueltos libres (Ilan y Abelson, 1995; Rützler, 1997). De ahí que se considere que sobre las plataformas continentales la diversidad de esponjas es mayor en fondos duros litorales que en fondos blandos (Pansini y Musso, 1991).
Conocidas como esponjas psammobióticas, las especies cuyos hábitos de vida les permiten desarrollarse exclusivamente dentro o sobre sedimentos arenosos, favorecen la conformación de microhábitats intersticiales, estabilizan el sustrato y atraen a otros organismos para conformar islas en mares de arena (Rützler, 1997). Ya antes, Werding y Sánchez (1991) demostraron como dos especies de esponjas presentes en Santa Marta, poseen adaptaciones morfológicas "raíces" para vivir enterradas; mientras Ilan y Abelson (1995) presentaron cómo una especie que vive semienterrada en lagos arrecifales someros del Mar Rojo, brinda dentro de su esqueleto espacio a gran cantidad de bacterias simbióticas intracelulares. Hacia el nor-oriente del Caribe colombiano no se conocen estudios sobre las adaptaciones de las esponjas, por lo que los hábitos de vida de las especies que allí habitan no han sido establecidos, debido a la falta de observaciones en el medio natural.
Un ecosistema con fondos semejantes a los de La Guajira se presenta en costas occidentales de la plataforma continental de África. Van Soest (1993) encontró que existen mayor cantidad de hábitats para esponjas en áreas someras y de fondos arenosos, donde la conformación de los nichos se relaciona con la disponibilidad de sustratos fijos y móviles, así como con los hábitos y formas de crecimiento de los individuos. Dicha conclusión es cierta, incluso para la plataforma continental de La Guajira, ya que los sedimentos areno-lodosos, presentes en varias de las estaciones donde se recolectaron poríferos, son áreas que permiten el establecimiento de especies que pueden vivir semienterradas en esos sustratos o de aquellas con la habilidad de aglutinar partículas carbonatadas libres y fragmentos de otros invertebrados, para apoyarse sobre ellas y no hundirse (Figura 4). Por su parte, las formas de crecimiento esféricas o semiesféricas observadas en algunas especies colectadas, deben permitir a los individuos transportarse por rotación y translación gracias a las corrientes y oleaje locales, ampliando las áreas de distribución poblacional, al mismo tiempo que evidencian el esquema sobre el habito no completamente sésil y la plasticidad fenotípica de algunas especies; hábito que ya ha sido observado en esponjas que habitan zonas someras en África occidental (Van Soest, 1993) y el norte del Mediterráneo (Mercurio et al., 2006).
En La Guajira, el afloramiento de núcleos de aguas profundas en áreas costeras, así como el depósito de detritus en los sedimentos blandos es continuo a lo largo del año (Criales-Hernández et al., 2006); de hecho la densidad media de macrofauna fue registrada como la mayor para las plataformas continentales del Caribe colombiano (COLCIENCIAS, 2006). Sobre estos sedimentos existe una variedad de al menos 53 especies de esponjas habitando en diferentes áreas someras (Díaz, 2007). Sin embargo, es necesario que se establezcan semejanzas y diferencias de la diversidad de esponjas entre fondos rocosos y blandos en el Caribe colombiano, con el propósito de corroborar el esquema de mayor diversidad de especies en fondos rocosos vs. fondos blandos mencionado por Pansini y Musso (1991) en costas del norte del Mediterráneo.
AGRADECIMIENTOS
Este estudio se realizó dentro el programa de Biodiversidad y Ecosistemas Marinos (BEM) del Instituto de Investigaciones Marinas y Costeras "José Benito Vives de Andréis" - INVEMAR, dentro del proyecto "Caracterización de la zona costera del departamento de La Guajira: una aproximación para su manejo integrado" desarrollado por el INVEMAR y la Corporación Autónoma Regional de La Guajira CORPOGUAJIRA (convenio marco de cooperación científica y técnica 024 del 2002). Muchas gracias a todas las personas que nos prestaron su colaboración, en especial a Guillermo Duque, Norella Cruz, el grupo de investigación del Museo de Historia Natural Marina de Colombia (MHNMC) y la tripulación del B/I Ancón. Este texto es parte del Trabajo de Grado de M. D. de la Universidad Jorge Tadeo Lozano, programa de Biología Marina, fruto de la colaboración entre los grupos de investigación Macrofauna de INVEMAR y Fauna Marina, Biodiversidad y Usos, de la Universidad Nacional de Colombia.
BIBLIOGRAFÍA
1 Alcolado, P. 1999. Comunidades de esponjas de los arrecifes del archipiélago Sabana-Camagüey, Cuba. Bol. Inv. Mar. Cost., 28: 95-124. [ Links ]
2 Álvarez-León, L., J. Aguilera-Quiñones, C. Andrade-Amaya y P Nowak. 1995. Caracterización general de la zona de surgencia en La Guajira colombiana. Rev. Acad. Col. Cienc., 19 (75): 679-694. [ Links ]
3 Bell, J. y D. Barnes. 2003. Effect of disturbance on assemblages: An example using porffera. Biol. Bull., 2 (205): 144-160. [ Links ]
4 Clarke, K. y R. Warwick. 2001. Change in marine communities: an approach to statistical analysis and interpretation. Manual of statistical software PRIMER-E, Plymouth, UK. 146 p. [ Links ]
5 COLCIENCIAS. 2006. 75 maneras de generar conocimiento en Colombia. Programa Nacional de Ciencia y Tecnología del Mar. Caso No. 22, Bogotá. 112 p. [ Links ]
6 Criales-Hernández, M., C. Garcia y M. Wolff. 2006. Flujos de biomasa y estructura de un ecosistema de surgencia tropical en La Guajira, Caribe colombiano. Rev. Biol. Trop., 54 (4): 1257-1282. [ Links ]
7 Díaz, C.M. 2007. Poriferos de la plataforma continental (10 - 50 m de profundidad) del departamento de La Guajira, Caribe colombiano. Tesis Biol. Mar., Universidad Jorge Tadeo Lozano, Santa Marta. 101 p. [ Links ]
8 Díaz, J.M. 1995. Zoogeography of marine gastropod in the southern Caribbean: A new look at provinciality. Carib. J. Sci., 31 (1-2): 104-121. [ Links ]
9 Díaz, M. y B. Ward. 1997. Sponge-mediated nitrification in tropical benthic communities. Mar. Ecol. Prog. Ser.,156: 97-107. [ Links ]
10 Hentschel, U., J. Hopke, M. Horn, A. Friedrich, J. Wagner y B. Moore. 2002. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl. Environ. Microbiol., 68 (9): 44314440. [ Links ]
11 Hooper, J. y R. Van Soest. 2002. Sistema porífera. A guide to the classification of sponges. Kluwer, Nueva York. 1810 p. [ Links ]
12 Ilan, M. y A. Abelson. 1995. The life of a sponge in a sandy lagoon. Biol. Bull., 189: 363-369. [ Links ]
13 INVEMAR. 2000. Programa Nacional de Investigación en Biodiversidad Marina y Costera PNIBM. INVEMAR-FONADE, MMA, Santa Marta. 83 p. [ Links ]
14 INVEMAR. 2004. Biodiversidad asociada a las plataformas de explotación de gas natural (Campo Chuchupa: La Guajira-Colombia). Programa de Biodiversidad y Ecosistemas Marinos (BEM). Informe final, INVEMAR, Santa Marta. 225 p. [ Links ]
15 INVEMAR. 2005. Informe del estado de los ambientes marinos y costeros en Colombia. INVEMAR, Santa Marta. No 8. 360 p. [ Links ]
16 INVEMAR-CORPOGUAJIRA. 2006. Caracterización de la zona costera del departamento de La Guajira: una aproximación para su manejo integrado. Componente macro-fauna a 10 y 50 m de profundidad. Informe final, INVEMAR, Santa Marta. 86 p. [ Links ]
17 Kaiser, M., J. Collie, S. Hall, J. Poiner y S. Jennings. 2001. Impacts of fishing gear on marine benthic habitats. Reykjavik Conference on Responsible Fisheries in the Marine Ecosystem, Islandia. 19 p. [ Links ]
18 Maldonado, M. y C. Young. 1998. Limits on the bathymetric distribution of keratose sponges: A field test in deep water. Mar. Ecol. Prog. Ser., 174: 123-139. [ Links ]
19 Mercurio, M., G. Corriero y E. Gaino. 2006. Sessile and non-sessile morphs of Geodia cydonium (Porífera, Demospongiae) in two semi-enclosed Mediterranean bays. Mar. Biol., 148: 489-501. [ Links ]
20 Montoya-Maya, P. 2002. Evaluación de la macrofauna epibentónica asociada a praderas de Thalassia testudinum (Banks ex Kóning) en el Caribe colombiano. Tesis Biol. Mar., Universidad Jorge Tadeo Lozano, Santa Marta. 74 p. [ Links ]
21 Pansini, M. y B. Musso. 1991. Sponges from trawl-exploitable bottoms of Ligurian and Tyrrhenian Seas: Distribution and ecology. PSZN I: Mar. Ecol., 12 (4): 317-329. [ Links ]
22 Parra-Velandia, F. y S. Zea. 2003. Comparación de la abundancia y distribución de algunas características de las esponjas del genero Ircinia en dos localidades contraste del área de Santa Marta, Caribe colombiano. Bol. Inv. Mar. Cost., 32: 75-91. [ Links ]
23 Reed, J. y S. Pomponi. 1997. Biodiversity and distribution of deep and shallow water sponges in the Bahamas. Proc. 8th Int. Coral Reef. Sym., 2: 1387-1392. [ Links ]
24 Reyes, J., N. Santodomingo, A. Gracia, G. Borrero-Pérez, L. Mejía-Ladino, A. Bermúdez y M. Benavides. 2005. Southern Caribbean azooxanthellate coral communities off Colombia. Cold-water corals and ecosystems. Springer-Verlag, Berlín. 1243 p. [ Links ]
25 Romero, J. 2006. Estudio sedimentológico de la plataforma continental de La Guajira entre los 10 y 50 m de profundidad. Tesis Geol., Universidad Nacional de Colombia, Bogotá. 96 p. [ Links ]
26 Rützler, K. 1997. The role of psammobiotic sponges in the reef community. Proc. 8th Int. Coral Reef. Symp. Panamá, 2: 1393-1398. [ Links ]
27 Suzunaga, J., L. Calero y A. Molina. 1992. Síntesis del estudio de las condiciones oceanográficas al N-E de La Guajira. Fase II. Boletín Científico C.I.O.H., 11: 59-69. [ Links ]
28 Valderrama, D. y S. Zea. 2003. Esquemas de distribución de esponjas arrecifales (Porifera) del noroccidente del golfo de Urabá, Caribe sur, Colombia. Bol. Inv. Mar. Cost., 32: 37-56. [ Links ]
29 Van Soest, R. 1993. Distribution of sponges on the Mauritanian continental shelf. Hydrobiologia, 25: 95-106. [ Links ]
30 Werding, B. y H. Sánchez. 1991. Life habits and functional morphology of the sediment infaunal sponges Oceanapia oleracea and Oceanapia peltata (Porifera, Haplosclerida). Zoomorphology, 110: 203208. [ Links ]
31 Worheide, G., A. Sole-Cava y J. Hooper. 2005. Biodiversity, molecular ecology and phylogeography of marine sponges: Patterns, implications and outlooks. Integ. Comp. Biol., 45: 377-385. [ Links ]
32 Zea, S. 1987. Esponjas del Caribe colombiano. Catálogo Científico. Bogotá. 286 p. [ Links ]
33 Zea, S. 1993. Recruitment of demosponges (porífera, Demospongiae) in rocky and coral reef habitats of Santa Marta, Colombian Caribbean. Mar. Ecol., 14 (1): 1-21. [ Links ]
34 Zea, S. 2002. Patterns of sponge (Porifera: Demospongiae) distribution in remote oceanic reef complexes of the southwestern Caribbean. Rev. Acad. Col. Cienc., 25: 579-592. [ Links ]
35 Zúñiga, H., J. Altamar y L. Manjarrés. 2004. Caracterización tecnológica de la flota de arrastre camaronero del mar Caribe de Colombia. Proyecto EP/GLO/201/GEF. Reducción by-catch. FAO. Universidad del Magdalena, Santa Marta. 20 p. [ Links ]
FECHA DE RECEPCIÓN: 31/01/08 FECHA DE ACEPTACIÓN: 17/06/08
