










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.38 no.1 Santa Marta Jan./June 2009
STRIOCADULUS MAGDALENENSIS, A NEW DEEP-SEA SCAPHOPOD (MOLLUSCA: SCAPHOPODA: GADILIDAE) FROM THE COLOMBIAN CARIBBEAN*
STRIOCADULUS MAGDALENENSIS UNA NUEVA ESPECIE DE ESCAFÓPODO (MOLLUSCA: SCAPHOPODA: GADILIDAE) DE AGUAS PROFUNDAS DEL CARIBE COLOMBIANO
Adriana Gracia C.1 and Néstor E. Ardila2
1Instituto de Investigaciones Marinas y Costeras-INVEMAR, Museo de Historia Natural Marina de Colombia, Santa Marta, Colombia, A.A. 1016. agracia@invemar.org.co
2Universidad de los Andes, Facultad de Ciencias, Departamento de Ciencias Biológicas, Laboratorio de Biología Molecular Marina (BIOMMAR), Carrera 1E N° 18a -10, Bogotá D.C., Colombia. ne.ardila23@uniandes.edu.co
ABSTRACT
We describe a new bathyal species of scaphopod belonging to the genus Striocadulus Emerson based on conchological and radular characters. The specimens were collected at a depth of 404-412 m off Bocas de Ceniza (Colombian Caribbean). Striocadulus magdalenensis new species can be distinguished from its other congeners by differences in its apical section, and both number and extent of longitudinal striae. This is the first record of this genus in the Atlantic Ocean.
KEY WORDS: Mollusca, Bathyal, Taxonomy, Western Atlantic, Caribbean Sea.
RESUMEN
Se describe una nueva especie batial de escafópodo perteneciente al género Striocadulus Emerson basándose en características de la concha y la rádula. Los especímenes fueron colectados a 404-412 m de profundidad frente a Bocas de Ceniza (Caribe colombiano). Striocadulus magdalenensis spec. nov. se distingue de sus otros congéneres por diferencias en la sección apical y en el número y extensión de las estrías longitudinales. Este es el primer registro del género para el océano Atlántico.
PALABRAS CLAVE: Mollusca, Batial, Taxonomía, Atlántico occidental, Mar Caribe.
INTRODUCTION
Scaphopods are successful deep-sea organisms with over 500 extant species (Scarabino, 1994; Steiner and Linse, 2000; Reynolds and Steiner, 2008). Nearly 70 % of the known species are found below 500 m of depth, with a high percentage of them living below 3000 m, exclusively inhabiting bathyal and abyssal substrates (Scarabino, 1986).
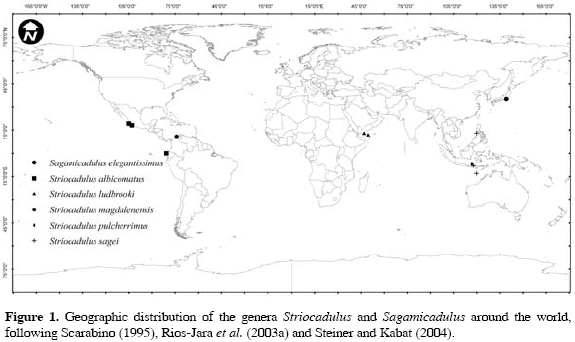
Among the eight genera within Gadilidae (Steiner and Kabat, 2001), Striocadulus Emerson, 1962 is represented by very few species compared to other genera in the family. The genus is very distinct and is comprised by species living in moderately deep waters in tropical oceans that are characterized by having medium to large curved shells, sculptured with numerous longitudinal striae According to Scarabino (1995) and Steiner and Kabat (2001, 2004), the genus currently consists of four living species with well delineated ranges. Striocadulus albicomatus (Dall, 1890) occurs along the west coast of North and Central America at depths of 41730 m; S. ludbrooki Scarabino, 1995 is distributed in the northern Indian Ocean at 528-732 m; S. pulcherrimus (Boissevain, 1906) has been recorded from Indonesia at 530-694 m, and S. sagei Scarabino, 1995 from the Philippines and Indonesia at 640-800 m (Figure 1). Herein we describe a new species recently collected on soft substrates of the upper slope off the Colombian Caribbean coast in two deep-sea trawled samples taken by the R/V Ancón in 1998, during the cruise INVEMAR-MACROFAUNA I (Figure 1). Shells were examined, and photographed under Scanning Electron Microscopy (SEM) (Leica Stereoscan 440 W and Amray 1810 SEM's). Material examined was deposited at the Museo de Historia Natural Marina de Colombia (MHNMC), Santa Marta, Colombia.
RESULTS
Species description
Family Gadilidae Stoliczka, 1868
Genus Striocadulus Emerson, 1962: 478-479, pl. 80, fig. 1 a-d (as a subgenus of Cadulus).
Type species: Cadulus albicomatus Dall, 1890, by original designation. Recent, off Manta, Ecuador; eastern Pacific Ocean, 00°37'S - 81°00'W, 730 m. Syntype, USNM 87539.
Known distribution: Pacific and Indian oceans, Caribbean Sea, 41-800 m.
Striocadulus magdalenensis new species
(Figures 2-8, 16-17)
Type locality:
Holotype: Off Bocas de Ceniza, Colombia, 11°13'N, 74°39'W, 412-406 m of depth, INV MOL2829, Sta. E2 (2-X-1998), on muddy bottom.
Paratypes: Off Bocas de Ceniza, Colombia, 11°13'N, 74°39'W, 404-410 m of depth, two specimens, INV MOL2828, Sta. E1 (2-X-1998), on muddy bottom; off Bocas de Ceniza, Colombia,11°13'N, 74°39'W, 412-406 m depth, one specimen, INV MOL2384, Sta. E2 (2-X-1998), on muddy bottom.
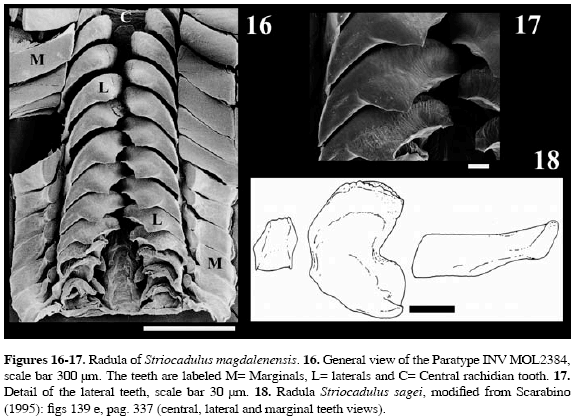
Type Material: Four specimens, collected alive. The measurements of the material are described in the Table 1.
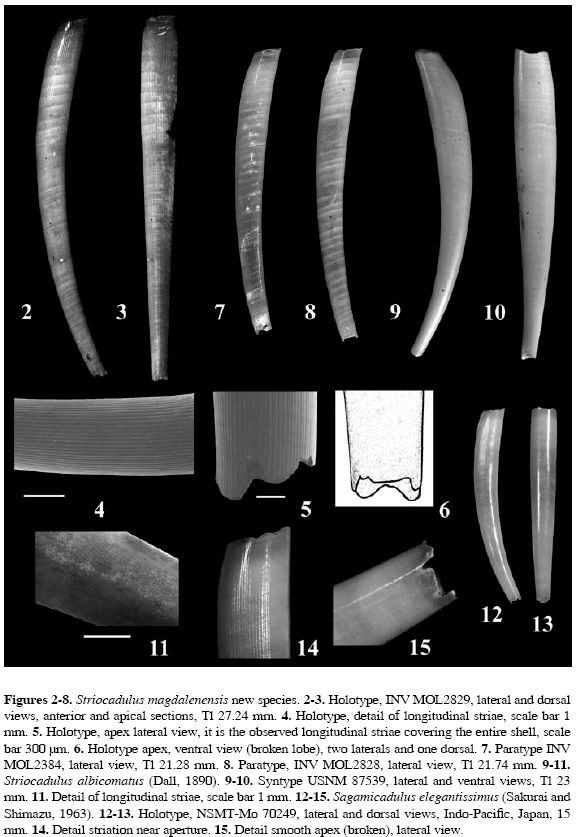
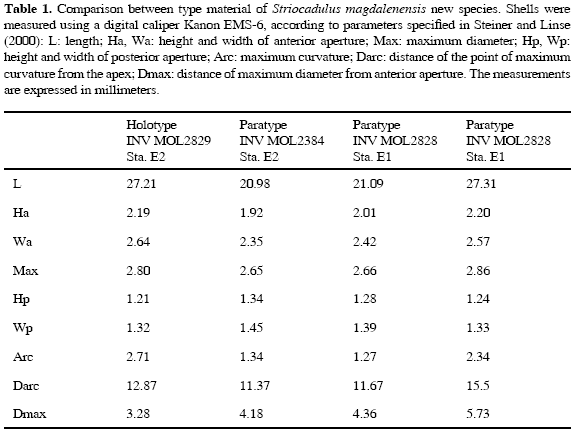
Distribution: This species is known only from the type locality. Description: Shell length up to 27 mm; white, bright, and translucent, with opaque white rings along the surface; ventral side slightly curved; maximum diameter 2.8 mm in anterior third proximal to the aperture Anterior aperture subcircular in cross section, dorsoventrally compressed, 2.19 mm in height and 2.64 mm in width. Apical aperture semicircular, dorsoventrally compressed, 1.21 mm in height and 1.32 mm in width (Table 1); apex with traces of four lobes (Figure 6). The shell is longitudinally sculptured with 66 (Holotype) to 73 fine, flattened striae (Figures 4-5) of equal length and evenly spaced covering the entire surface. The radula typical of the genus consists of a square central rachidean tooth, with large lateral and long marginal teeth; shape and denticulation of the central, lateral, and marginal teeth are not visible (Figures 16-17).
Etymology: The species is named after the Magdalena River, in front of whose mouth the type locality is situated.
Remarks: This species shares common characteristics with the species of the monotypical genus Sagamicadulus, S. elegantissimus (Sakurai and Shimazu, 1963: 250-251, text-fig. 1 Holotype; Hasegawa and Saito, 1995: 29, plate 6, fig. 1 Holotype), recorded off Hayama, Sagami Bay, Japan, at depths between 55 and 128 m (Figures 12-15). This smaller species reaches 15 mm in length (Holotype NSMT-Mo 70249) and its shell also exhibits longitudinal grooves, but they do not cover the entire surface (Figure 15). The shell is smooth posteriorly and longitudinally striated in median and anterior areas, with the striae extending to the aperture (Fig. 14); in S. magdalenensis striae are uniformly distributed over the surface. The apical orifice in S. elegantissimus is rounded in cross section and has three pairs of notches on its truncated margin (Figure 15), two pairs on the lateral surfaces and one pair on the dorso-ventral surfaces (Sakuari and Shimazu, 1963), whereas the semicircular apex of S. magdalenensis has traces of four lobes. The number and shape of the lobes in S. magdalenensis are not exactly described since all the examined specimens were not complete.
It is noteworthy that this diagnostic character differs somewhat from the shape and number of lobes assigned to other species of Striocadulus and could match better to the Sagamicadulus diagnosis However, due to the extension of the longitudinal striae, the longer shell length, and the semicircular apex in cross section, we prefer to place it within the genus Striocadulus.
Striocadulus magdalenensis exhibits unique characteristics compared with the other members of the genus, especially in the sculpture and apex morphology Striocadulus sagei, the largest species in the order Gadilida (Scarabino, 1995), found in Indonesia and Philippines, reaches 43 mm in length, exhibits a wide apex, and is slightly dorsoventrally compressed, with two wide lateral lobes (Scarabino, 1995). The anterior aperture is oblique and dorsoventrally compressed The longitudinal sculpture consists of numerous fine, flattened riblets, which are narrowly spaced apart and crossed throughout by growth lines Conversely, S. ludbrooki is smaller (up to 20 mm) and less sculptured than S. sagei, its apex is oval in cross section and dorsoventrally compressed with two wide, flat lobes (Scarabino, 1995); moreover, the sculpture of the longitudinal striae is most prominent along the anterior three quarters of the shell. Detailed information about the shell features of S. pulcherrimus from Indonesia was unfortunately not available; Scarabino (1995) just illustrates the general shape of the type material (16 mm in length), the apex structures and apical and oral sections.
In regard to the radula, although it was extracted and examined (Paratype INV MOL2384), no additional diagnostic characteristics could be determined (Figures 16-17); for details of the radula of S. sagei, see Scarabino (1995: figs 139 e, pag. 337, and fig. 172 c-d, pag. 367). Here we reproduce the type radula of the genus (Figure 18).
Striocadulus magdalenensis was compared with the syntype (USNM 87539) of S. albicomatus (Dall, 1890), the geographically closest congener, and the main difference is that S. albicomatus exhibits a greater number of longitudinal striae (more than 100), but displays the same outline and sculpture pattern (Figures 9-11). Both species have a slightly oblique aperture and a nearly circular apical-anal aperture. Dall (1890) and Emerson (1962) do not give further details upon the apex, but S. albicomatus clearly has a stronger apex, bearing two weak, flat lateral lobes (Scarabino, 1995). In his description of the subgenus Striocadulus, Emerson (1962) stated that the apical orifice is simple and stands out as a diagnostic characteristic.
It should be emphasized that the diagnostic characters of the two genera of the family Gadilidae characterized by longitudinal striae (Striocadulus and Sagamicadulus) are still incomplete. It is important to obtain more material for a detailed study of its internal and external morphology in order to define the taxonomical status of both genera.
The bathymetric distribution of the genus appears to be homogeneous, as the five described species have been mostly found at depths between 404 and 800 m, with the exception of S. albicomatus, which has been recorded at 41-73 m off the Mexican Pacific coast (Ríos-Jara et al., 2003a, 2003b), although previous records from Panama and Ecuador were from much deeper waters. This new record of Striocadulus extends the distributional range for the genus from the Pacific and Indian Oceans to Atlantic waters (Figure 1).
ACKNOWLEDGEMENTS
We want to thank to INVEMAR-MACROFAUNA and the R/V Ancón teams for collaboration during collecting expeditions We also want to acknowledge the institutional and financial support provided by INVEMAR and COLCIENCIAS (Grants No. 2105-13-079-97, 210509-11248 and 2115-09-16649) and the Colombian Ministry of the Environment. S. Braden (NMNH-USA) assisted with the production of the SEM images. Y. Villacampa (NMNH-USA) and H. Saito (National Museum of Nature and Science NSMT-Japan) kindly provided the Syntype and Holotypes images of the examined material respectively. We are thankful to N. Rangel for providing the map of the study area. The authors thank J. Clayton for the improvement of the English version and J. M. Díaz for the exhaustive review of the document.
LITERATURE CITED
1 Boissevain, M. 1906. The Scaphoda of the Siboga Expedition, treated together with the known Indopacific Scaphopoda. 1-76. In: Weber, M. (Ed.). Uitkomsten op zoologisch, botanisch, oceanographisch en geologisch gebied verzameld in Nederlandsch Oost-Indië 1899-1900 aan boord H. M. Siboga onder commando van Luitenant ter zee le. kl. G. F. Tydeman. E. J. Brill, Leiden (Netherlands), 54 (32): 76 p. [ Links ]
2 Dall, W. H. 1890. Scientific results of explorations by the U. S. fish commission steamer Albatross. VII-preliminary report on the collection of Mollusca and Brachiopoda obtained in 1887-88. Proc. U. S. Nat Mus., 12 (773): 219-362. [ Links ]
3 Emerson, W. K. 1962. A classification of the scaphopod molluscs. J. Paleont., 36 (3): 461-482. [ Links ]
4 Hasegawa, K. and H. Saito. 1995. Ilustrations and annotated checklist of the molluscan type specimens contained in the Sakurai Collection in the National Science Museum, Tokyo. National Science Museum, Tokyo. 55 p. [ Links ]
5 Reynolds, P.D. and G. Steiner. 2008. Scaphopoda. 143-161. In: Ponder, W. F. and D. R. Lindberg (Eds.). Phylogeny and evolution of the Mollusca. University of California Press, Berkeley and Los Angeles, California, EE UU 469 p. [ Links ]
6 Ríos-Jara, E., E. López-Uriarte, M. Pérez-Peña and E. Juárez-Carrillo. 2003a. Nuevos registros de escafópodos para las costas de Jalisco y Colima, México. Hidrobiológica, 13 (2): 167-170. [ Links ]
7 Ríos-Jara, E., M. Pérez-Peña, E. López-Uriarte and E. Juárez-Carrillo. 2003b. Moluscos escafópodos de la campaña Atlas V (Plataforma continental de Jalisco y Colima, México) a bordo del B/O El Puma. Cienc. Mar., 29 (1): 67-76. [ Links ]
8 Sakuarai, K. and T. Shimazu. 1963. A new shell Striocadulus (Sagamicadulus) elegantissimus subgen. et sp. nov. from Sagami Bay, Japan. Bull. Nat. Sci. Mus., 6 (3): 250-251. [ Links ]
9 Scarabino, V. 1986. Nuevos taxa abisales de la clase Scaphopoda (Mollusca). Com. Zool. Mus. Hist. Nat. Montevideo, 11 (155): 1-19. [ Links ]
10 Scarabino, V. 1994. Aproximación al número de especies de Scaphopoda actuales. Com. Soc. Malac. Urug., 7: (66-67): 357-361. [ Links ]
11 Scarabino, V. 1995. Scaphopoda of the tropical Pacific and Indian Oceans, with description of 3 new genera and 42 species. 654 p. In: Bouchet, P. (Ed.), Résultats des Campagnes MUSORSTOM, Vol. 14. Mém. Mus. nat. Hist. natur., 167: 189-379. [ Links ]
12 Steiner, G. and A. R. Kabat. 2001. Catalogue of supraspecific taxa of Scaphopoda (Mollusca). Zoosystema, 23 (3): 433-460. [ Links ]
13 Steiner, G. and A. R. Kabat. 2004. Catalog of species-group names of recent and fossil Scaphopoda (Mollusca). Zoosystema, 26 (4): 549-726. [ Links ]
14 Steiner, G. and K. Linse. 2000. Systematics and distribution of the Scaphopoda (Mollusca) in the Beagle Channel (Chile). Mitt. Hamburg. Zool. Mus. Inst., 97: 13-30. [ Links ]
15 Stoliczka, F. 1868. Cretaceous fauna of Southern India II: Gastropoda (Prosocephala) Paleontologica Indica, being figures and descriptions of the organic remains procured during the progress of the Geological Survey of India. Mem. Geol. Surv. India, Calcutta 5: 13. [ Links ]
FECHA DE RECEPCIÓN: 04/05/07 FECHA DE ACEPTACIÓN: 13/04/09
* Contribución No. 1033 del Instituto de Investigaciones Marinas y Costeras-INVEMAR.
