










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.38 no.1 Santa Marta Jan./June 2009
INDICATORS OF INFLUENCE OF SALINITY IN THE RESISTANCE AND RESILIENCE OF FISH COMMUNITY IN A TROPICAL COASTAL LAGOON (SOUTHEASTERN BRAZIL)
INDICADORES DE LA INFLUENCIA DE SALINIDAD EN LA RESISTENCIA Y RESILIENCIA DE LA ICTIOFAUNA EN LAGUNA COSTERA TROPICAL (SUDOESTE DE BRASIL)
Jorge Iván Sánchez-Botero1, Danielle Sequeira Garcez2, Erica Pellegrini Caramaschi3 and Adriana Miguel Saad4
1 Universidade Federal do Ceará (UFC)- Campus do Pici, Centro de Ciencias, Depto de Biologia-Bloco 906. CEP: 60455-970 Fortaleza, Ceará. jisbar@gmail.com
2 Universidade Federal do Rio de Janeiro, Instituto de Geociências, Departamento de Geografia. CCMN, Caixa Postal 68537, CEP: 21945-970, Rio de Janeiro, Brasil.
3 Universidade Federal do Rio de Janeiro/Instituto de Biologia - Departamento de Ecologia. CCS, Caixa Postal 68020, CEP: 21941-590, Rio de Janeiro, Brasil.
4 Universidade Estácio de Sá, sede Cabo Frio/RJ - Departamento de Biologia, CEP: 28909-800, Cabo Frio, Rio de Janeiro, Brasil.
ABSTRACT
The effects of a disturbance by sandbank opening on the fish fauna community were evaluated through a long-term study in the Imboassica coastal lagoon, Rio de Janeiro State, Brazil. The fish and salinity samples were conducted between 1993 and 2004, with the goal of characterizing the structure and effects of four sandbank openings on this community, such as resilience and resistance measurements. During three sandbank openings, between the years of 1993 and 1996, the resilience time of the fish community could not be identified due to the short time period between openings, their magnitude and the discontinuity of the fish sampling. Monitoring studies previously conducted between 1993 and 2004, indicated that factors structuring the fish fauna of Imboassica lagoon are due to salinity fluctuations, influenced by the sandbank openings to which this environment is subjected. These events were indicated by the recolonization of primary freshwater species, oligohaline salinity values, and small variations in the composition and abundance on the fish community. Due to the magnitude and frequency of the disturbance, the ichthyic populations available that enter the lagoon from the adjacent ocean plus the resistance of this environment to salinity variations, determined the resilience and resistance of the ish fauna in the Imboassica lagoon. Thus, the resilience of the ish fauna in this lagoon was indicated in groupings of: a) Spearman probability values lower than 0. 05 between the pairs of communities pre and post-disturbance; b) presence of freshwater species that indicate resilience; c) salinity values varying between 0 and 5; and d) small alterations in the abundance of fish populations.
KEY WORDS: Disturbance, Fish community, Coastal lagoon, Brazil.
RESUMEN
Los efectos del disturbio de cuatro aberturas de la barra de arena en la laguna costera Imboassica, localizada en el municipio de Macaé, estado de Río de Janeiro, Brasil, en la comunidad de peces fueron evaluados en un estudio a largo plazo. Entre los años 1993 y 2004, colectas de peces y valores de salinidad fueron obtenidos con el objetivo de caracterizar la estructura y los efectos de las aberturas de barra sobre esta comunidad, como medidas de resiliencia y resistencia. Durante las aberturas de la barra realizadas entre los años de 1993 y 1996, el tiempo de resiliencia de la comunidad no fue identificado debido a los cortos intervalos de aberturas, a la magnitud del disturbio y a la discontinuidad de los muestreos de peces. Estudios realizados entre 1993 y 2004 indicaron que los factores que estructuran la ictiofauna de la laguna Imboassica son consecuencia de las variaciones de la salinidad, influenciadas por las aberturas de la barra a que este ambiente es sometido. Estos eventos fueron indicados por la recolonización de las especies primarias de agua dulce, valores de salinidad oligohalinos y bajas alteraciones en la composición y abundancia de la comunidad de peces. Debido a la magnitud y frecuencia de los disturbios, las poblaciones evaluadas de peces que entran en la laguna provenientes del océano adyacente, más la resistencia de este ambiente a las variaciones de salinidad, determinan la resiliencia y resistencia de la fauna de peces en la laguna. Así, la resiliencia de la fauna íctica en la laguna Imboassica fue indicada por el conjunto de: a) valores de probabilidad de Spearman menores de 0. 05 entre los pares de comunidades pre y pos disturbio; b) presencia de especies de peces de agua dulce indicadoras de resiliencia; c) intervalos de salinidad variando entre 0 y 5; y d) bajas alteraciones en la abundancia de las poblaciones de peces.
PALABRAS CLAVE: Disturbio, Comunidad de peces, Laguna costera, Brazil.
INTRODUCTION
In coastal lagoons and estuary systems in general the effects of seasonal fluctuations of abiotic factors on communities are important sources of variations (Longhurst and Pauly, 1987). These seasonal fluctuations affect, primarily, the composition and abundance of species, imposing cyclic and non-cyclic modifications (Ricklefs, 1973; Lowe McConnell, 1987). Although the composition and abundance of species varies in time and space, Livingston (1976) declares that the basic structure of estuarine and coastal lagoons fish communities is stable, or, at least, predictable. This means that there is a spatial distribution and abundance pattern of species that is stable in time or predictable regarding seasonal fluctuations of abiotic factors in the system. Disturbances in the environment and the area history can lead to long-term temporal variations, reflecting certain aspects of the distribution patterns exhibited by some species (Barrela and Petrere Jr., 1993). The characterization of variations in ichthyofauna compositions in coastal lagoon systems, such as local displacements of resident species or certain migratory fluxes, are important to distinguish these natural movements from others, caused by environmental alterations of antropic origin (McErlean et al., 1973).
The severing of the sandbank that separates a lagoon from the adjacent sea, commonly known as a sandbank opening, represents, in the case of environments identified as "isolated lagoons" (Esteves, 1998), a short period of disturbances. These events cause sudden changes, especially in the physical structure of the lagoon and in salinity values, which alter the structure of the communities (Esteves, 1998; Frota and Caramaschi, 1998; Saad et al., 2002; Kozlowsky-Suzuki and Bozelli, 2004, Santangelo et al., 2007). The effects of this disturbance in biotic and abiotic factors were amply documented in coastal lagoons of Africa (Albaret and Ecoutin, 1989), Australia (Pollard, 1994; Griffiths, 1999), Mexico (Yafiez-Arancibia and Nugent, 1997) and Brazil (Kozlowsky-Suzuki et al., 1998; Mello and Suzuki, 1998; Suzuki et al., 1998; Saad et al., 2002; Caramaschi et al., 2004; Kozlowsky-Suzuki and Bozelli, 2004; Santangelo et al., 2007).
The Imboassica lagoon, located in the municipality of Macae (Rio de Janeiro State - southeastern Brazil), is subject to eventual contacts with the Atlantic Ocean, artificially caused by human intervention (Frota and Caramaschi, 1998), consequently causing variations in biotic and abiotic factors (Branco, 1998; Petrucio, 1998; Santangelo, 2002; Meirelles-Pereira, 2003; Caramaschi et al., 2004; Kozlowsky-Suzuki and Bozelli, 2004). Particularly regarding the ichthyofauna, during artificial sandbank openings alterations in the composition and abundance of young, adults and larvae that inhabit this lagoon were registered (Frota, 1998; Soares, 1998, 2004; Saad et al., 2002; Caramaschi et al , 2004). These changes were influenced mainly by salinity, turbidity, and water temperature variations, which stimulated the entrance of estuarine-dependent ish species and the migration and death of freshwater fish (Frota and Caramaschi, 1998; Caramaschi et al., 2004), revealing an unstable taxocenosis. This type of disturbance leaves coastal lagoons permanently closed in conditions of constant ecological instability, since they interrupt the process of natural succession.
Therefore, identifying the sandbank openings as an opportunity to test the effects of this disturbance in the structure of the ish community, the present study evaluated the alterations in the temporal patterns of ish fauna abundance and the consequent variations in salinity, as measurements of resilience and resistance. In this way, resilience, evaluated as the ability of the community to return to its original state (or nearly so) prior the disturbance (Webster et al., 1975; Harrison, 1979; Gunderson, 2000; Begon et al., 2007), was evaluated taking in account the variations of the community composition and the abundance of the ish populations, influenced by the change in salinity. Resistance, however, was evaluated taking into account the small or null variations in population abundance regarding the salinity changes, as a consequence of the various sandbank openings.
STUDY AREA
The Imboassica coastal lagoon, located in a suburban area in the municipality of Macaé/RJ (22° 24' S and 41° 42' W), was formed by the damming of the Imboassica river; it is isolated from the adjacent sea by a 50 m wide sandbank (Figure 1). This environment occupies an area of 326 hectares, with a maximum width of 1.3 km and maximum length of 5.3 km. It is a shallow system, with average depth of 1.1 m, reaching 2-3 m in the deeper areas (Panosso et al., 1998). The marginal vegetation of the Imboassica lagoon is constituted, basically, of taboa banks (Typha domingensis) near to the river and of grasses in the sand barrier region (Palma-Silva, 1998). The sediment is muddy in the middle and far end regions of the lagoon and sandy in the area near the sandbank. Due to the high-energy characteristics of the sea in the adjacent area to the lagoon, the sand strip should not present any more natural openings (Panosso et al., 1998).
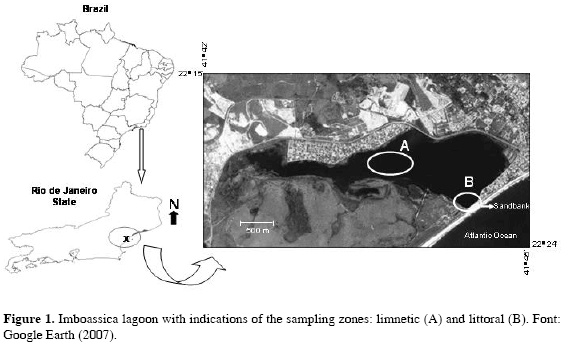
According to Esteves (1998), the Imboassica lagoon suffers mainly from anthropogenic disturbances: landfill along its shores, domestic sewage disposal, and artificial sandbank openings followed by salinity increases. These events cause changes in biotic and abiotic factors (Esteves, 1998; Caramaschi et al., 2004). The partial freshwater replacement by marine water quickly shifted water conditions near the sandbank, within a few days (ten to ifteen) after the sandbank opening, the water change from strongly oligohaline to slightly euhaline.
MATERIAL AND METHODS
Between October/1993 and June/2004 60 experimental fish samplings and salinity measurements with a digital thermosalinometer (YSI model 30SET) were conducted at Imboassica Lagoon. During the entire period, four artificial sandbank openings were monitored in three different studies. In the study conducted by Saad (1997) the sandbank opening occurred in April 1994, with a total of 12 fish samplings, six before and six after the opening. In the study conducted by Frota (1998) two sandbank openings occurred: in November 1995, with four monthly fish samplings, before and after the opening; and in April 1996, when two fish samplings were conducted after the opening. In February 2001, the sandbank opening occurred after being closed for two years Six samplings were conducted before this last opening and 28 after it, until the end of this study in June 2004 (Table 1).

Basically, two methodologies were used, which allowed the identification and comparison of abundance variation and ichthyofauna composition:
a) In the limnetic zone of the lagoon, 14 gill nets (meshing of 15, 20, 25, 30, 35, 40 and 45 mm between adjacent knots) were exposed during approximately 12 hours in the samplings conducted by Saad (1997) and six to nine hours in the present study and in the one conducted by Frota (1998). The exposed battery areas of the gill nets was, in average, 573.82 m2 in the study conducted by Saad (1997), 596.87 m2 in the study conducted by Frota (1998), and 332.28 m2 in the present study. After this, the catch per unit effort (CPUE) (Appendix 1) for each study was calculated as the following:
CPUE = N/(m2 x h)
Where:
N = ∑ of the number of individuals
m2 = ∑ of the net areas
h = Number of hours the nets remained in the water
b) With a seine net of 11.5 m x 1.8 m and 1.5 cm between adjacent knots, three diurnal drags of 50 m distance were conducted by Saad (1997) and in the present study, in shallow littoral zone near the sandbank. In the study conducted by Frota (1998) seven drags of 30 m distance were conducted, with the same net, in the same zone and at the same time. After this, the catch per unit effort (CPUE) for each study was calculated from the number of individuals per capture area dragged per campaign. The ichthyofauna abundances were later converted to m2 using a simple rule of three (Appendix 2).The captured fish were put on ice and transported to the NUPEM/UFRJ (Núcleo de Pesquisas Ecológicas de Macaé - Macaé Ecological Research Center) laboratory, where the species were determined using taxonomic identification keys and quantified.
DATA ANALYSIS
To identify the resilient and resistant fish species regarding the disturbance (the sandbank opening) an analysis of indicator species was conducted (Dufrene and Legendre, 1997), including pre and post sandbank openings, based on the salinity values obtained between 1993 and 2004. The selected factor (salinity) demonstrated an ample variation during the sampling periods (average = 8.5; Cv = 102 %). Salinity was measured using the Practical Salinity Scale. In this way, it was possible to group the campaigns and, consequently, the fish species into categories according to salinity values during the sampling periods in: freshwater (< ±0.5), oligohaline (±0.5 to ±5), mesohaline (±5 to ±18), polyhaline (±18 to ±30), and mixoeuhaline (> ±30), as proposed by Esteves (1988). The indicator species were identified as those occurring most frequently and characteristically in one type of salinity category. Thus, with the values of CPUE in the samplings with gill nets and seine nets, independently, for each species i comparing all pre or post sandbank opening periods j, by Bij, identifying the relative occurrence frequency of i species in the j group as follows:
Aij = CPUEij/CPUEi
Bij = CPUEperiodsij/CPUEperiodsj
INDVALij = Aij x Bij x 100
Where INDVAL corresponds to the indicator value of species i in the grouping of period j. Aij is the specific measurement and CPUEij is the mean number of individuals from species i by periods of group j, while CPUEi corresponds to the mean sum of individuals of species i over all periods. Bij corresponds to the measurement of idelity, with Nperiodsij identified as the number of periods in grouping j where species i is present, while Nperiodsj is the total number of periods in that group.
The Monte Carlo method was applied to test the significance of the indicator value of the species (INDVAL), using 10000 permutations in the four sampling periods that include periods of pre and post-openings of the sandbank, as indicated in Table 1. The probability (p) of the indicator value of the species is indicated by:
p = (1 + number of observed groups)/(1 + number of randomized groups)
Therefore, from this analysis, probabilities of less than 0.05 indicated species with resilience. However, fish species resistant to salinity variations that occurred in the pre and post-openings of the sandbank were indicated by the relative abundance of each group, in percentages. Or else, by the mean abundance of each species by salinity category over the mean abundance of that species in all categories.
With the non-parametric Spearman correlation coefficient (Wolda, 1981), the fish abundances (CPUE) of the campaigns before the opening and post-opening were compared. Probabilities lower than 0.05 indicated significant similarities in the composition and abundance of the ish community between the compared campaigns before the opening, and pre and post sandbank opening. On the other hand, probability values higher than 0.05 represent significant differences between the community composition and abundance, indicating the resilience time of the community right after the disturbance.
Ecological fish guilds indicate the use and importance of a coastal lagoon for species: coastal lagoon resident (CLR), that completes its entire life cycle in a coastal lagoon; marine dependent (MD), that spawn in the sea and use coastal lagoons as nurseries for larvae and juveniles or for adult permanence for long periods; marine visitors (MV), that use coastal lagoons occasionally, and freshwater primary and secondary (FWp and FWs), that have no apparent saline requirements, according to selected bibliography (Myers, 1937; Araujo, 1983; Aranha, 1991; Vieira and Scalabrin, 1991; Aguiaro, 1994; Saad, 1997; Soares, 1998; Carvalho Filho, 1999; Fisher et al., 2004; Froese and Pauly, 2004).
RESULTS
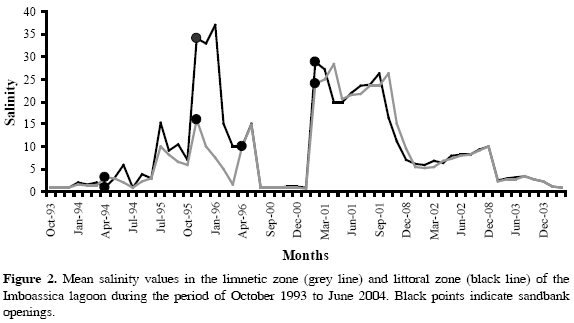
During studies conducted between the years of 1993 and 2004 in Imboassica lagoon, 60 species of fish were collected belonging to 26 families and 14 orders. In the limnetic zone 44 species of fish were captured, belonging to 21 families and 10 orders. In the littoral zone, 44 species of fish were captured, belonging to 22 families and 12 orders. The mean abundance varied with the pre and post-opening sandbank periods (Appendix 1 and 2). The ecological fish guilds were 41.7 % marine dependent (MD), 33.3 % marine visitors (MV), 10.0 % coastal lagoon resident (CLR), 8.3 % primary freshwater (FWp), and 6.7 % secondary freshwater (FWs) by the total lagoon. Salinity values varied between 0.74 and 28.32 (average = 8.04; Cv = 101.07 %) for the limnetic zone and between 0.90 and 37.00 (average = 9.52; Cv = 102.69 %) for the littoral zone, demonstrating an increasing trend during the four sandbank opening periods (Figure 2).
The Indicator Species Analysis based on salinity variations pointed out Astyanax bimaculatus and Hoplias malabaricus (FWp) as resilient when the lagoon reaches oligohaline values in the case of the first species (average salinity = 2.25; Cv = 43.70 %), and mesohaline, in the case of the second species (average salinity = 5.52; Cv = 55.62 %), with significant indicator values (p < 0.05). These values were identified by the Monte Carlo test with 9999 unrestricted permutations, during four pre and post sandbank opening periods and 57 monthly samplings with gill nets in the limnetic zone of the Imboassica lagoon between the years of 1993 and 2004 (Table 2). On the other hand, probabilities lower than 0.05 were not obtained in the littoral zone because of the absence of primary freshwater species that are indicators of resilience.

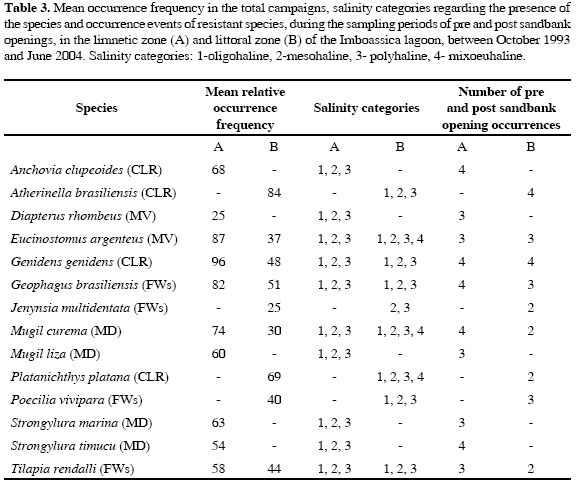
The same analysis identified in the limnetic zone of the lagoon ten resistant fish species during the periods of pre and post sandbank openings: two coastal lagoon residents (CLR), two secondary freshwater species (FWs), two marine visitors (MV) and four marine dependent species (MD) (Table 3). In the littoral zone nine resistant species were identified during the pre and post-sandbank opening periods: one marine visitor (MV), one marine dependent (MD), three coastal lagoon residents (CLR) and four secondary freshwater species (FWs) (Table 3).
In the twelve monthly samplings conducted by Saad (1997) in the sand barrier zone, six occurring before the opening (1 to 6) and six after (7 to 12), 71.0 % of the sampling pairs before the sandbank opening were similar in composition and abundance (Spearman p < 0.05). On the other hand, 83.3 % of the sampling pairs before and after the sandbank opening presented different community compositions and abundances (Spearman p > 0. 05). In the samplings conducted by Frota (1998) in the same zone, 50 % of the campaigns were significantly similar before the opening (1 to 4) and 35 and 30 % were significantly similar after the first and second sandbank openings, respectively (1 to 9 and 5 to 11). In the samplings conducted in the present study of the sand barrier zone, 93.3 % of the sampling pairs before the opening were similar (1 to 6). However, 80.4 % of the sampling pairs compared before and after the opening (1 to 6 with 7 to 34) were significantly similar (Spearman p < 0.05).
Comparing the sampling pairs of the limnetic zone of the study conducted by Saad (1997), 93.3 % of the sampling pairs before the sandbank opening (1 to 6) were significantly similar in composition and abundance. On the other hand, between the pre and post-opening campaigns (1 to 6 with 7 to 12), 17.2 % of these were significantly similar (Spearman p < 0.05). In the samplings conducted by Frota (1998) in the same zone, 100 % of the campaigns were significantly similar before the opening (1 to 2) and 70 and 90 % were significantly similar after the irst and second sandbank openings, respectively (3 to 7 with 8 to 9). In the samplings conducted in the present study of the limnetic zone, 100 % of the sampling pairs before the opening (1 to 6) were significantly similar. However, comparing the sampling pairs before and after the opening (1 to 6 with 7 to 33), 80.2 % were significantly similar (Spearman p < 0.05).
During month 14 (average salinity = 6.40), after the last sandbank opening, the species H. malabaricus (FWp) was recaptured in the limnetic zone of the lagoon. In the capture periods in which this species was present before and after the opening, an average of 95.2 % of the Spearman probability values between communities were lower than 0.05, indicating similar composition and abundance. During the pre and post sandbank openings in the samplings conducted by Saad (1997), this species was also captured with salinity values ranging between 1 and 9.91 The resilience indicator species A. bimaculatus (FWp) was recaptured 27 months after the last sandbank opening (average salinity = 3.10). All the Spearman probability values, comparing the pre and post sandbank opening campaigns where this species was captured were lower than 0.05, indicating 100 % of similarity in these communities composition and abundance.
DISCUSSION
The opening of the sandbank at the Imboassica lagoon causes a drastic reduction in the water volume by draining freshwater to the sea, followed by seawater entrance, which rapidly elevates the salinity values, altering the ish fauna structure. Fish mortality, freshwater ish migrations to refuges at swamps and/or flooded adjacent areas to the lagoon and the entry of estuarine-dependent species, caused primarily by the increase in the salinity values, were registered during these periods. These disturbances compromise the establishment of several freshwater fish populations in Imboassica lagoon and after each opening, a new community was formed, with an abundant number of species considered rare or presenting a low occurrence frequency (Saad, 1997; Frota, 1998; Frota and Caramaschi, 1998; Palma-Silva, 1998; Saad et al., 2002; Caramaschi et al., 2004; Triverio et al., 2004).
The 14 fish species indicated as resistant to salinity variations represented 23 % of the total taxa captured in this environment. Evidence of successful reproductive activity in this environment was remarked for the following species: Platanichthys platana, Atherinella brasiliensis, Genidens genidens (CLR), Jenynsia multidentata, Poecilia vivipara, Tilapia rendalli, and Geophagus brasiliensis (FWs) (Saad, 1997; Caramaschi et al., 2004), indicating that they complete their entire life cycle in this environment, obtaining a continual individual recruitment, independent of the salinity variations. However, the persistence of Anchovia clupeoides (CLR), Mugil curema, Mugil liza, Strongylura timucu, Strongylura marina (MD), Diapterus rhombeus, and Eucinostomus argenteus (MV) without evidence of successful reproduction in Imboassica lagoon was due, possibly, to the entrance of a great number of individuals from these species during the opening events or to marine undertows that go beyond the littoral zone, transporting larvae and juvenile ish into the lagoon (Soares, 1998, 2004).
As ish communities of coastal lagoons are submitted to constant stress conditions by sporadic or permanent contact with adjacent marine and freshwater environments (Winemiller and Leslie, 1992; Yafez-Arancibia and Nugent, 1997; Caramaschi et al., 2004), the number of resistant species found by this study (23 % of the total) almost reached that found in a previous study (30 %) (Saad et al , 2002). This fact indicates the high dependence that most occasional visitor species, marine dependent, coastal lagoon residents, and freshwater fish sampled during these periods in Imboassica lagoon have on the marine and/or freshwater environment in which they complete their life cycle. The presence of resistance indicator species was favored by the artificial sandbank openings and by the ample dominance of the oligohaline conditions in this environment.
Community responses to biotic and abiotic disturbances depend on their frequency and magnitude. For example, the ichthyofauna in North Oregon (USA) streams, when disturbed by floods, presented resilience time of five to six days influenced by the habitat complexity (Pearsons et al., 1992). In the estuarine lagoon system of Swartvlei (South Africa), fish resilience regarding environmental alterations, such as the disappearance of submerse aquatic macrophytes, was approximately three years (Whitfield, 1986). The resilience of the zooplankton community in the Imboassica lagoon remained for three years before opening sandbar, which allowed the establishment of a different community after contact with the sea (Santangelo et al., 2007).
However, in the present study, during three sandbank opening events (between 1993 and 1996), the resilience of the fish community could not be identified, due to the short period of time between the openings, their magnitude, and the discontinuity in the fish sampling in Imboassica Lagoon. Significant values in Spearman correlations during the three pre and post-opening periods were indicated by the similarity in the marine dependent (MD) and coastal lagoon resident (CLR) species composition and abundance in the campaign pairs that were compared or by the frequency of rare species (MV) and their influence in the sensibility of this test On the other hand, the capture of the resilience indicator freshwater species H. malabaricus in the periods after sandbank opening, in the samplings conducted by Saad (1997) with dominant oligohaline values, was influenced by the short time that the sandbank remained open (compared to other evaluated openings) and by the high rain regime during this period However, the capture of H. malabaricus and A. bimaculatus at 14 and 27 months, respectively, after the closure of the last sandbank opening, indicate that resilience time of the ichthyofauna in this environment varied regarding to the opening magnitude, the variation in the salinity values, and the local rain regime. Indicators of the resilience of ichthyofauna in this environment occur when the lagoon reaches oligohaline or near oligohaline salinity values (< 5), the secondary freshwater species G. brasiliensis, T. rendalli, J. multidentata and P. vivipara abundance are maintained constant in the months prior to the disturbance, and the evidence of recolonization of primary freshwater species fish, such as H. malabaricus and A. bimaculatus. The sandbank opening occurred in February 2001 was preceded by an extended drought period (El Niño event) that favored the persistence of mesohaline values of water during twenty-three months after the disturbance. This fact compromised the reestablishment of a fish fauna with freshwater characteristics. In the sand barrier zone this event was even more evident where the absence of resilient freshwater species and the dominance of resident estuarine species and secondary freshwater species were favored by the salinity values.
Significant values in the community's similarity (83.3 % in Spearman comparisons; p < 0.05), with the occurrence of the indicative species H. malabaricus, before and after the sandbank opening in the limnetic zone of Imboassica lagoon, suggest a structural return regarding abundance and composition fourteen months after the disturbance. Evaluating resilience as the community ability to return to its original state as before the disturbance (Webster et al., 1975; Harrison, 1979; Gunderson, 2000), it was observed that the return time of the fish community happened when salinity presented oligohaline or near oligohaline (< 5) values. This proposal is established in the premise that the sandbank that separates the lagoon from the sea will unlikely be open again by natural events; this is due to the high gravity marine waves that dominate this area and keep enlarging the size of the sandbank (Panosso et al , 1998). In this way, the lagoon should naturally present growing freshwater characteristics and the ichthyofauna that would dominate this environment should possess a higher proportion of freshwater fish, and coastal lagoon resident (CLR), marine dependent (MD), or visitor (MV) species in lesser proportions. On the other hand, the marine dependent (MD) or visitor (MV) species, affected negatively by the dominant oligohaline conditions in the lagoon, could suffer an increment in the water content of their muscles, causing abnormal behavior and, in most cases, their deaths (Wu and Woo, 1983). Besides this, the null reproductive ability of some marine dependent (MD) or visitor (MV) species originating from this environment (Saad, 1997) would compromise the recruiting of new individuals, leading these populations to disappear.
Thus, the resilience time of fish fauna in Imboassica lagoon was indicated in groupings of: a) Spearman probability factors lower than 0.05 between pre and post-disturbance pairs of communities; b) the presence of resilience freshwater fish indicator species; c) salinity values ranging between 0 and 5; and d) minor alterations in fish population abundance. A similar situation evaluated during the same months in Imboassica lagoon seems to be related to the resilience of the zooplanktonic community, where increased similarities between the pre and post sandbank openings are associated to salinity values lower than 5.9 (Santangelo, 2002).
From this study, observations regarding the factors that structure the ichthyofauna of Imboassica lagoon could be made, which are in part caused by the sandbank openings where the effects of salinity variations were worth mentioning. In this way, the magnitude and frequency of the sandbank opening disturbance, as well as the available fish populations in the adjacent ocean that enter the lagoon and/or the resistance of this environment to biotic and abiotic variations, determine the resilience and resistance of the fish fauna of Imboassica lagoon. The recolonization of primary freshwater fish species, oligohaline salinity values, and minor alterations in the composition and abundance of the fish populations indicate these events.
CONCLUSIONS
Anthropogenic disturbances alter the fish community in the coastal lagoon Imboassica, north of Rio de Janeiro State. Four artificial openings in the sandbank of the Imboassica lagoon and connectivity with adjacent sea have modified abundance and composition of the ichthyofauna, however fourteen fish species have shown to be resistant to changes resulting from this disturbance. Fourteen months after the last sandbank opening, the recapture of the freshwater species H. malabaricus evidenced a resilience process associated to salinity reduction which showed a similar pattern in the composition and abundance of the pre and post disturbance in the fish communities during these periods. The disturbances mentioned indicate that the knowledge of the fish fauna variability in these coastal environments require a series of continuous monitoring studies that should involve annual and pluriannual cycles. The great variety of ecological guilds of fish species found in the Imboassica lagoon has showed an environment with a large availability of resources to this fauna.
ACKNOWLEDGEMENTS
We are grateful to Dr. Francisco Assis Esteves and M. Sc. Andre Ferreira for valuable suggestions; M. Sc. Pedro Hollanda de Carvalho for help in fish identification; and to the staff of Laboratory of Fish Ecology from Federal University of Rio de Janeiro (UFRJ) for field assistance, specially to M. Sc. Rafael Pereira Leitäo and M. Sc. Victor Trivério Cardoso. CAPES, PELD site-5, CNPq, and FAPERJ supported this study, which was part of the doctorate degree (PhD. Thesis) carried out by the first author (JISB) at PPGE/UFRJ, Brazil.
LITERATURE CITED
1 Aguiaro, T. 1994. Estrutura da comunidade de peixes de très lagoas costeiras da regiäo de Macaé (RJ). M. Sc. thesis, Universidade Federal do Rio de Janeiro. Rio de Janeiro. 114 p. [ Links ]
2 Albaret, J. J. and J. M. Ecoutin. 1989. Communication mer-lagune: impact d'une réouverture sur l'ichtyofaune de la lagune Ébrié (Côte d'Ivore). Hydrob. Trop., 22 (1): 71-81. [ Links ]
3 Aranha, J. M. R. 1991. Distribuiçào longitudinal, ocupaçâo ambiental, alimentaçào e época reprodutiva de quatro espécies de Cyprinodontiformes (Osteichthyes) coexistentes no rio Ubatiba (Maricâ, RJ). M. Sc. thesis, Universidade Federal do Rio de Janeiro. Museu Nacional. Rio de Janeiro, Brasil. 130 p. [ Links ]
4 Araújo, F. G. 1983. Distribuiçao, abundancia, movimentos sazonais e hábitos alimentares de bagres marinhos (Siluriformes, Ariidae) no estuário da lagoa dos Patos (RS), Brasil. M. Sc. thesis, Rio Grande. Fundaçao Universidade do Rio Grande, Brasil. 189 p. [ Links ]
5 Barrela, M. and M. Petrere Jr. 1993. The influence of environmental factors on fish community structure in the Jacaré Pepira River. 161-170. In: Cowx, I. (Ed.). Rehabilitation of inland fisheries. Oxford, U. S. A. 486 p. [ Links ]
6 Begon, M., C. R. Townsend and J. L. Harper. 2007. Ecologia de individuos a ecossistemas. 4th ed Editora Artmed, Porto Alegre. 740 p. [ Links ]
7 Branco, C. W. C. 1998. Composiçao e aspectos ecológicos das comunidades zooplanctônicas nas lagoas Imboassica, Cabiúnas e Comprida. 247-271. In: Esteves, F. A. (Ed.). Ecologia das lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do Municipio de Macaé (RJ). Universidade Federal do Rio de Janeiro, Rio de Janeiro. 464 p. [ Links ]
8 Caramaschi, E. P., J. I. Sánchez-Botero, P. Hollanda-Carvalho, C. A. S. Brandao, C. L. Soares, J. L. C. Novaes and R. B. Araújo. 2004. Peixes das lagoas costeiras de Macaé: Estudos de caso. 309-337. In: Rocha, C.F.D., F.A. Esteves y F.R. Scarano (Eds.). Pesquisas de longa duraçao na Restinga de Jurubatiba: ecologia, historia natural e conservaçao. Sao Carlos, Brasil. 376 p. [ Links ]
9 Carvalho Filho, A. 1999. Peixes da costa brasileira 3rd edition. Ed. Melro, Sao Paulo. 304 p. [ Links ]
10 Dufrene, M. and P. Legendre. 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol. Monog., 67 (3): 345-366. [ Links ]
11 Esteves, F. A. 1988. Fundamentos de limnologia. Interciência/FINEP, Rio de Janeiro. 575 p. [ Links ]
12 Esteves, F. A. 1998. Lagoas costeiras: Origem, funcionamento e possibilidades de manejo. 63-87. In: Esteves, F. A. (Ed.). Ecologia das lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do Municipio de Macaé (RJ). Universidade Federal do Rio de Janeiro, Rio de Janeiro. 464 p. [ Links ]
13 Fisher, L. G., D. L. E. Pereira and J. P. Vieira. 2004. Peixes estuarinos e costeiros. Editora Ecoscientia, Rio Grande do Sul, Brasil. 126 p. [ Links ]
14 Froese, R. and D. Pauly. 2004. Fish base. World Wide Web Electronic Publication. http://www.fishbase.org. 01/2008. [ Links ]
15 Frota, L. O. R. 1998. Dinàmica temporal da taxocenose de peixes de uma lagoa costeira impactada por aberturas artificiais da barra (Lagoa Imboassica, Macaé, RJ). Master thesis, Universidade Federal do Rio de Janeiro. Rio de Janeiro. 132 p. [ Links ]
16 Frota, L. O. R. and E. P. Caramaschi. 1998. Aberturas artificiais da barra da lagoa Imboassica e seus efeitos sobre a ictiofauna de peixes. 327-350. In: Esteves, F. A. (Ed.). Ecologia das lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do Municipio de Macaé (RJ). Universidade Federal do Rio de Janeiro, Rio de Janeiro. 464 p. [ Links ]
17 Google Earth. 2007. http://earth.google.com. [ Links ]
18 Griffiths, S. P. 1999. Consequences of artificially opening coastal lagoons on their fish assemblages. Int. J. Salt. Lake Res., 8: 307-327. [ Links ]
19 Gunderson, L. H. 2000. Ecological resilience - in theory and application. Annu. Rev. Ecol. Syst., 31: 425-439. [ Links ]
20 Harrison, G. W. 1979. Stability under environmental stress: resistance, resilience, persistence and variability. Amer Nat., 113: 659-669. [ Links ]
21 Kozlowsky-Suzuki, B. and R. L. Bozelli. 2004. Resilience of a zooplankton community subjected to marine intrusion in a tropical coastal lagoon. Hydrobiología, 522:165-177. [ Links ]
22 Kozlowsky-Suzuki, B., C. W. C. Branco and R. L. Bozelli. 1998. Comparison of the composition and the temporal variation of the zooplanktonic community of two Brazilian coastal lagoons. Verh. Int. Ver. Limnol., 26: 1487-1490. [ Links ]
23 Livingston, R. J. 1976. Diurnal and seasonal fluctuations of organisms in a north Florida estuary. Est. Coast. Mar. Sci., 4: 373-400. [ Links ]
24 Longhurst, A. R. and D. Pauly. 1987. Ecology of tropical oceans. Academic, San Diego. 470 p. [ Links ]
25 Lowe McConnell, R. H. 1987. Ecological studies in tropical fish communities. Cambridge University, Cambridge. 382 p. [ Links ]
26 McErlean, A. J., S. G. O'Connor, J. A. Mihursky and C. I. Gibson. 1973. Abundance, diversity and seasonal patterns of estuarine fish populations. Est. Coast. Mar. Sci., 1: 19-36. [ Links ]
27 Meirelles-Pereira, F. 2003. Influencia da variacäo da profundidade da coluna d'água sobre a ecologia do Lago Batata (PA, Oriximiná) e da Lagoa Imboassica (RJ, Macaé). M. Sc. thesis, Universidade Federal do Rio de Janeiro. Rio de Janeiro. 115 p. [ Links ]
28 Mello, S. de and M. S. Suzuki. 1998. Variacòes temporais e espaciais do fitoplancton das lagoas Imboassica, Cabiúnas e Comprida. 177-203. In: Esteves, F. A. (Ed.). Ecologia das lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do Municipio de Macaé (RJ). Universidade Federal do Rio de Janeiro, Rio de Janeiro. 464 p. [ Links ]
29 Myers, G. S. 1937. Fresh-water fishes and West Indian zoogeography. Ann. Rep. Smith. Inst., 3465: 339-364. [ Links ]
30 Palma-Silva, C. 1998. Crescimento e producào de TyphadomingensisPers na Lagoa Imboassica. 205-220. In: Esteves, F.A. (Ed.). Ecologia das lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do Municipio de Macaé (RJ). Universidade Federal do Rio de Janeiro, Rio de Janeiro. 464 p. [ Links ]
31 Panosso, R. F., J. L. Attayde and D. Muehe. 1998. Morfometria das Lagoas Imboassica, Comprida e Carapebus: implicacòes para seu funcionamento e manejo. 91-105. In: Esteves, F.A. (Ed.). Ecologia das lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do Municipio de Macaé (RJ). Universidade Federal do Rio de Janeiro, Rio de Janeiro. 464 p. [ Links ]
32 Pearsons, T. N., H. W. Li. and G. A. Lamberti. 1992. Influence of habitat complexity on resistance to flooding and resilience of stream fish assemblages. Trans. Amer. Fish. Soc., 121: 427-436. [ Links ]
33 Petrucio, M. M. 1998. Caracterizacào das lagoas Imboassica, Cabiúnas, Comprida e Carapebus a partir da temperatura, salinidade, condutividade, alcalinidade, O2 dissolvido, pH, transparencia e material em suspensào. 109-122. In: Esteves, F. A. (Ed.). Ecologia das lagoas costeiras do Parque Nacional da Restinga de Jurubatiba e do Municipio de Macaé (RJ). Universidade Federal do Rio de Janeiro, Rio de Janeiro. 464 p. [ Links ]
34 Pollard, D. A. 1994. A comparison of fish assemblages and fisheries intermittently open and permanently open coastal lagoons on the south coast of New South Wales, south-eastern Australia. Estuaries, 17: 631-646. [ Links ]
35 Ricklefs, R. E. 1973. Ecology. Publisher Newton, Massachusetts, Chiron Press, New York. 861 p. [ Links ]
36 Saad, A. M. 1997. Influencia da abertura da barra sobre a comunidade de peixes da lagoa Imboassica, Macaé, RJ. Master thesis, Universidade Federal do Rio de Janeiro. Rio de Janeiro. 130 p. [ Links ]
37 Saad, A. M., A. C. Beaumord and E. P. Caramaschi. 2002. Effects of artificial canal openings on fish community structure of Imboassica coastal lagoon, Rio de Janeiro, Brazil. J. Coast. Res., 36: 634-639. [ Links ]
38 Santangelo, J. M. 2002. Estrutura e resiliência da comunidade zooplanctônica de uma lagoa costeira após uma abertura de barra (Lagoa Imboassica, Macaé, RJ). Monograph, Universidade Federal do Rio de Janeiro. Rio de Janeiro. 19 p. [ Links ]
39 Santangelo, J. M., A. M. Rocha, R. L. Bozelli, L. S. Carneiro and F. A. Esteves. 2007. Zooplankton responses to sandbar opening in a tropical eutrophic coastal lagoon. Estuar. Coast. Shelf. Sci., 71: 657-668. [ Links ]
40 Soares, C. L. 1998. Composiçao e distribuiçao espaço-temporal do ictioplàncton das lagoas Imboassica, Cabiúnas e Comprida, Macaé, RJ. M. Sc. thesis, Universidade Federal do Rio de Janeiro. Rio de Janeiro. 131 p. [ Links ]
41 Soares, C. L. 2004. Densidade, distribuiçao e composiçao do ictioplàncton da lagoa Imboassica (Macaé, RJ) e do litoral adjacente antes e depois de uma abertura de barra. Dr. thesis, Universidade Federal do Rio de Janeiro. Rio de Janeiro. 173 p. [ Links ]
42 Suzuki, M. S., A. R. C. Ovalle and E. A. Pereira. 1998. Effects of sand bar openings on some limnological variables in a hypertrophic tropical coastal lagoon of Brazil. Hydrobiologia, 368: 111-122. [ Links ]
43 Trivério, V. C., J. I. Sánchez-Botero and E. P. Caramaschi. 2004. Refúgio dos peixes dulcicolas quando o mar invade a lagoa Imboassica (Macaé, RJ). Livro de Resumos do XXV Congresso Brasileiro de Zoologia. Brasilia. 333 p. [ Links ]
44 Vieira, J. P. and C. Scalabrin. 1991. Migraçao reprodutiva da tainha (Mugil platanus GUNTER, 1980) no sul do Brasil. Rev. Atlàn., 13: 131-141. [ Links ]
45 Webster, J., J. B. Waide and B. C. Patten. 1975. Nutrient cycling and the stability of ecosystems. In: Howell, F. G. (Ed.) Mineral cycling in southeastern ecosystems. Atomic Energy Comm. Symp. Ser. Conf. 325 p. [ Links ]
46 Whitfield, A. K. 1986. Fish community structure response to major habitat changes within the littoral zone of an estuarine coastal lake. Environ. Biol. Fish., 17: 41-51. [ Links ]
47 Winemiller, K. and M. A. Leslie. 1992. Fish assemblages across a complex, tropical freshwater/marine ecotone. Environ. Biol. Fish., 34: 29-50. [ Links ]
48 Wolda, H. 1981. Similarities indices, sample size, and diversity. Oecologia, 50: 296-302. [ Links ]
49 Wu, R. S. S. and N. Y. S. Woo. 1983. Tolerance of hypo-osmotic salinities in thirteen species of adult marine fish: implications for estuarine fish culture. Aquaculture, 32: 175-181. [ Links ]
50 Yáñez-Arancibia, A. and R. S. Nugent. 1997. El papel ecológico de los peces en estuarios y lagunas costeras. An. Centro Cienc. Mar. Limnol., 4: 107-114. [ Links ]
FECHA DE RECEPCIÓN: 04/05/07 FECHA DE ACEPTACIÓN: 13/04/09
