










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.38 no.1 Santa Marta Jan./June 2009
NOTA:
PECES Y MACROINVERTEBRADOS MÓVILES DE HÁBITATS ARTIFICIALES EN LA BAHÍA DE TAGANGA, CARIBE COLOMBIANO
FISHES AND MOBILE MACROINVERTEBRATES OF ARTIFICIAL HABITATS IN TAGANGA BAY, COLOMBIAN CARIBBEAN
Óscar Delgadillo-Garzón
MoAm Monitoreos Ambientales, Calle 127a # 53a-68, Apto. 502, Bogotá, Colombia. oskadelgadillo@gmail.com
ABSTRACT
Fishes and macroinvertebrates associated to two types of artificial habitats deployed in Taganga Bay were assessed by stationary visual census to compare these assemblages. Forty-one species of fishes and nine macroinvertebrates were observed in eleven visual censuses. The distribution patterns depicted in abundance and richness between the two types of habitats used (concrete blocks vs. artificial shelters), were a possible response to dissimilarities in their location and number of visual census. A relative high fish richness and diversity were found in these artificial habitats, in comparison to other artificial structures in Colombia, probably owing to their proximity with heterogeneous natural habitats. The use of complex artificial habitats could have potential useful to conserve, restore and manage some marine resources supported in planned researches in this area.
KEY WORDS: Artificial habitats, Fishes, Macroinvertebrates, Resource management, Taganga Bay.
La bahía de Taganga y la región de Santa Marta tienen una gran variedad de comunidades marinas presentes en diferentes ecosistemas y ambientes, con evidente importancia para la economía local, las cuales han estado sometidas al impacto de actividades antrópicas, mostrando actualmente claros signos de degradación ambiental (Díaz et al., 2000). En esta zona no se ha explorado el potencial de los hábitats artificiales (HAs), como herramientas útiles para el manejo y conservación de la biodiversidad. Los HAs pueden ser definidos como uno o más objetos dispuestos intencionalmente sobre el fondo marino, con el fin de influenciar procesos físicos, biológicos y socioeconómicos relacionados con los recursos marinos (Seaman y Jensen, 2000). En otras áreas del Caribe colombiano como Cartagena (Botero et al., 1981), islas del Rosario (Muñoz y Obregón, 1994), golfo de Morrosquillo (Herazo y Torres, 1998; Gámez et al., 2001; Delgadillo et al., 2004; Delgadillo, 2005), Santa Marta (Martínez, 2005) y La Guajira (Santodomingo et al., 2005) se han realizado evaluaciones de peces e invertebrados en HAs que han documentado su eficacia como refugio para las especies. A pesar de esto, hasta el momento, en el país se han efectuado pocas investigaciones para evaluar el potencial a largo plazo de los HAs como herramientas de manejo ambiental.
Para el caso de la langosta espinosa (Panulirus argus), entre el 2003 y el 2004 la Fundación Sila Kangama evaluó el reclutamiento de este crustáceo utilizando HAs, con el fin de mejorar su manejo en la región de Santa Marta (Jaimes et al., 2004). Dados los resultados positivos de los HAs diseñados para langostas, surge la pregunta de si los mismos son útiles para agregar peces y otros invertebrados. Por ello, este estudio describe y compara dichas asociaciones en dos tipos de hábitats en la bahía de Taganga, con fines de determinar su potencial uso en la región.
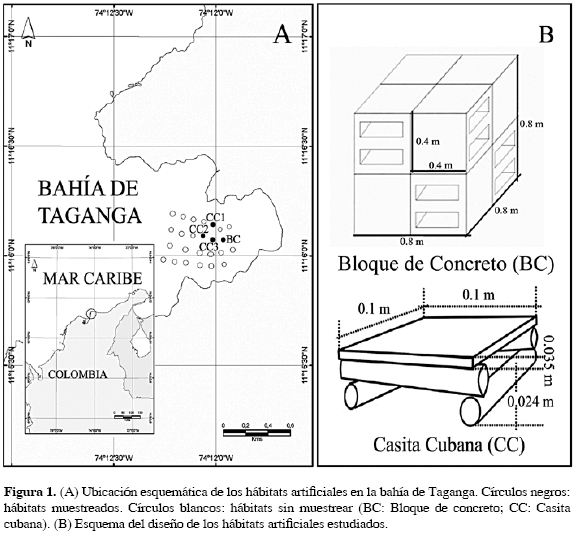
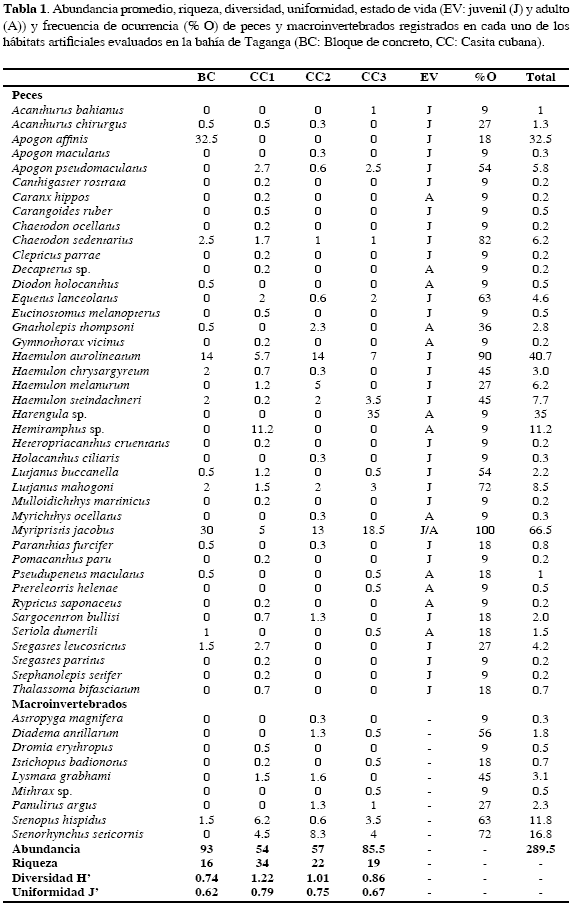
En noviembre de 2002 se instalaron 20 casitas cubanas (CC) y cinco módulos de bloques de concreto (BC), dispuestos aproximadamente en forma de cuadrícula y separados cada uno alrededor de 100 m, entre 20 y 30 m de profundidad en la bahía de Taganga (11° 16' - 11° 14' N, 74° 10' - 73° 56' W; Figura 1). Teniendo en cuenta las limitaciones de buceo y la accesibilidad a los hábitats, se escogieron tres CC y un módulo de BC a 25 m de profundidad, como muestra representativa de los hábitats instalados. En abril de 2004 se realizaron once censos visuales utilizando SCUBA, con el método de cilindro estacionario (Bohnsack y Bannerot, 1986), para estimar la composición, abundancia, frecuencia y estados de vida de peces y macroinvertebrados en cada HA. El número de censos por habitat se distribuyó de la siguiente forma: CC1 (4), CC2 (3), CC3 (2) y BC (2), consecuente con la facilidad de encontrar los hábitats durante el muestreo, puesto que sólo uno de ellos (CC1) estaba señalizado con boya. Se calcularon los índices de diversidad y uniformidad de Shannon y Pielou, respectivamente (Clarke y Warwick, 2001), utilizando el programa Biodiversity 2.0 ®. Estos resultados se usaron para comparar las asociaciones de organismos entre CC y BC, así como con los resultados de otros hábitats artificiales evaluados en Colombia. Con el fin de determinar diferencias en la abundancia y debido a la gran cantidad de ceros encontrados en las matrices, estos valores fueron reemplazados por uno y posteriormente los datos se transformaron con logaritmo natural para hacer un análisis de varianza de una vía (ANOVA) con el programa StatGraphics Plus 4.0 ® (Zar, 1996).
En los cuatro hábitats evaluados se observaron 41 especies de peces y 9 de macroinvertebrados móviles, cuya sumatoria total de abundancia fue de 289±20 ind. y su promedio de 1. 43±0.39. El BC mostró una sumatoria promedio de 92±19 ind., mientras que las CC concentraron 63±32 ind. El promedio de abundancia para cada uno de los hábitats fue: BC = 1.84±6. 4; CC1 = 1.08±2.09; CC2 = 1.10±2.9; CC3 = 1.70±5.6; por lo demás, no se encontraron diferencias significativas de abundancia entre el BC y las CC.
Se observaron seis especies comunes en los hábitats y 27 exclusivas de cada uno (Tabla 1) Los peces de mayor abundancia en orden descendente fueron Myripristis jacobus, Haemulon aurolineatum y Harengula sp.; mientras que los individuos juveniles ocuparon el 76.6 % de la abundancia. La riqueza y abundancia de invertebrados fue substancialmente menor en el BC (1 y 1.5, respectivamente) comparado con las CC (9 y 12.1, respectivamente). Por su parte, los crustáceos conformaron el 67 % de la abundancia de invertebrados, entre tanto Stenorhynchus seticornis y Stenopus hispidus tuvieron los valores más altos.
Los resultados estuvieron influenciados por el corto período de muestreo, pero reflejan la naturaleza de las asociaciones en los HAs en esa época del año. La riqueza de peces (41 especies) fue menor comparada con la mayoría de estructuras artificiales en Colombia, como plataformas de gas (92 especies), arrecifes artificiales de acero (62 especies), colectores flotantes (54 especies) y una avioneta (40 especies). No obstante, estos hábitats tuvieron mayor volumen, complejidad estructural, tiempo de disposición y muestreo que los HAs de Taganga, por lo cual la riqueza en los HAs puede considerarse relativamente alta (Muñoz y Obregón, 1994; Delgadillo et al. 2004; Martínez, 2005; Santodomingo et al., 2005). Aunque los hábitats naturales adyacentes a los HAs ejercen inluencia sobre las variables evaluadas e igualmente afectan la validez de las comparaciones, es de anotar que los colectores, la avioneta, el BC y las CC tuvieron un gradiente similar en cuanto a su distancia de los hábitats naturales (entre 20 y 200 m, aproximadamente), siendo mayor la de los HAs, lo cual soporta la idea de una diversidad alta en los hábitats de Taganga.
La elevada riqueza en los HAs fue el resultado de su capacidad para incrementar el espacio de asentamiento de las especies, causado por la cantidad de hábitats instalados y la cercanía a los hábitats naturales. Esto es concordante con otros HAs de volumen reducido, los cuales mantienen una riqueza, abundancia y densidad similar o superior que los HAs de gran tamaño, debido al efecto de borde y a un mayor perímetro con respecto al radio del área (Bohnsack et al., 1994; Frazer y Lindberg, 1994; Delgadillo et al., 2004). La anterior idea también tiene sustento en el hecho de que los HAs en Taganga tuvieron un porcentaje considerable (entre 22. 7 % y 27.5 %) de los peces registrados en arrecifes naturales de la región (Reyes-Nivia et al., 2004; Olaya-Restrepo et al., 2008) y mayor riqueza de lo observado en pastos marinos de la bahía (33 especies; Ardila et al., 1997). Igualmente, el alto porcentaje de peces juveniles en los HAs de Taganga fue similar a módulos de piedra caliza con un 67 % (Cummings, 1994) y una avioneta con el 61 % (Muñoz y Obregón, 1994), lo que demuestra la importancia de estos hábitats como sitios de reclutamiento para las especies.
El número de especies de invertebrados en Taganga fue menor en comparación con otros HAs (Delgadillo, 2005; Santodomingo et al., 2005), debido probablemente a la escasa complejidad estructural, disponibilidad de refugio, reducido esfuerzo de muestreo y fuerte depredación, como lo registraron Frazer y Lindberg (1994) para estos organismos. Por su parte, la escasa abundancia de P. argus en comparación con el trabajo de Arango et al., (2001) y Briones-Fourzán y Lozano-Álvarez (2001) en HAs, puede ser consecuencia de la alta y continua presión sobre el recurso por parte de los pescadores, lo que sumado a la evidente degradación del hábitat, posiblemente ha afectado las poblaciones de langosta espinosa (Díaz et al., 2000; Jaimes et al., 2004).
Los patrones de distribución en abundancia y riqueza en los dos tipos de HAs, estuvieron mediados aparentemente por el número de censos realizados en cada uno y por la distancia de los ecosistemas naturales, más que por el diseño de los mismos, ya que no se encontraron diferencias entre el BC y las CC, además de observar valores totales y promedio similares entre el BC y la CC3. Los efectos de los factores anteriormente mencionados, que en determinados casos no se pueden controlar (Portier et al., 2000), se hacen evidentes en la CC1, donde el mayor número de censos y cercanía de los hábitats naturales, probablemente ocasionaron las diferencias con el BC y las otras CC, a pesar de no existir una base para comparar las asociaciones en los mismos tipos de hábitats (CC). Por otra parte, la baja riqueza de especies en el BC con respecto a las CC, en especial de los invertebrados y su abundancia, puede tener relación con la presencia de peces depredadores y competitivamente dominantes como Apogon affinis y M. jacobus, que excluyen a otras especies, como observaron Delgadillo et al. (2004), en arrecifes artificiales.
Finalmente, en vista del alto número de organismos observados, la presencia de especies de importancia comercial (P. argus y Lutjanus bucanella) y ecológica (Diadema antillarum), algunas de las cuales son vulnerables y se encuentran amenazadas, existe evidencia del potencial uso de los HAs. Es importante implementar diseños de hábitats que cumplan los requerimientos de estas especies, y realizar investigaciones enfocadas a evaluar la dinámica de las asociaciones en el tiempo, con el fin de mejorar el manejo y conservación de los recursos marinos en la región.
AGRADECIMIENTOS
Este trabajo fue financiado por la Fundación Sila Kangama y el Fondo para la Acción Ambiental, dentro del proyecto "Variabilidad del asentamiento post larval e implementación de refugios artificiales para el manejo de la langosta espinosa Panulirus argus en la región de Santa Marta". Se agradece a Carlos A. Trujillo por su colaboración.
BIBLIOGRAFÍA
1 Arango, L., M. L. Manrique y J. C. Jaimes. 2001. Reclutamiento de juveniles de la langosta Panulirus argus con refugios artificiales en el Caribe colombiano. Rev. Invest. Mar., 22 (3): 221-228. [ Links ]
2 Ardila, N , D. von Schiller, S. Moreno, D. Alonso y G. Ospina. 1997. Peces epibentónicos asociados a Thalassia testudinum, en la bahía de Taganga, Magdalena, Caribe colombiano. 39. En: Santos, A. y L. Nieto (Eds.). IV Simposio Colombiano de Ictiología. Resúmenes de conferencias y exposiciones. Universidad del Magdalena, Santa Marta. 123 p. [ Links ]
3 Bohnsack, J. y S. Bannerot 1986. A stationary visual census technique for quantitatively assessing community structure of coral reef fishes. NOAA Technical report NMFS, 41: 1-15. [ Links ]
4 Bohnsack, J., D. E. Harper, D. B. McClellan y M. Hullsbeck. 1994. Effects of size on colonization and assemblage structure of fishes at artificial reefs off southeastern Florida, U.S.A. Bull. Mar. Sci., 55 (2-3): 796-823. [ Links ]
5 Botero, J., J. Garzón y G. Gutiérrez. 1981. Establecimiento y desarrollo de la comunidad íctica en un arrecife artificial construido con llantas de desecho. Bol. Mus. Mar., 10: 63-81. [ Links ]
6 Briones-Fourzán, P. y E. Lozano-Álvarez. 2001. Effects of artificial shelters (casitas) on the abundance and biomass of juvenile spiny lobsters Panulirus argus in a habitat-limited tropical reef lagoon. Mar Ecol. Prog. Ser , 221: 221-232. [ Links ]
7 Clarke, K. R. y R. M. Warwick. 2001. Changes in marine communities. An approach to statistical analysis and interpretation. Segunda edición. Primer-E Ltd., Plymouth, Reino Unido. 136 p. [ Links ]
8 Cummings, S. L. 1994. Colonization of a nearshore artificial reef at Boca Ratón (Palm Beach county), Florida. Bull. Mar Sci., 55 (2-3): 1193-1215. [ Links ]
9 Delgadillo, O. 2005. Motile macroinvertebrates assemblages in two artificial reefs of the Morrosquillo Gulf, colombian Caribbean. 22. En: Cowan Jr., J.H. (Ed.). 8th International Conference on Artificial Reefs and Artificial Habitats, 8th CARAH. Abstracts, Biloxi, EE. UU. 109 p. [ Links ]
10 Delgadillo, O., C. García y J. Correa. 2004. Dinámica temporal de la asociación de peces en dos arrecifes artificiales del golfo de Morrosquillo, Caribe colombiano. Actual. Biol., 26 (81): 120-131. [ Links ]
11 Díaz, J., L. Barrios, M. Cendales, J. Garzón-Ferreira, J. Geister, M. López-Victoria, G. Ospina, E. Parra-Velandia, J. Pinzón, B. Vargas-Angel, F. Zapata y S. Zea. 2000. Áreas coralinas de Colombia. Serie de Publicaciones Especiales 5. INVEMAR, Santa Marta. 176 p. [ Links ]
12 Frazer, T. K. y W. J. Lindberg. 1994. Refuge spacing similarly affects reef-associated from three phyla. Bull. Mar Sci., 55(2-3): 388-400. [ Links ]
13 Gámez, L., H. Sáenz-Betancourt y L. Mejía. 2001. Estructura de la ictiofauna asociada a dispositivos agregadores de peces (DAP) en aguas costeras del Golfo de Morrosquillo, Caribe colombiano. 229. En: Resúmenes, Noveno Congreso Latinoamericano sobre Ciencias del Mar, IX Colacmar. San Andrés isla, Colombia. 535 p. [ Links ]
14 Herazo, D. y A. Torres. 1998. Evaluación de la efectividad pesquera de dos arrecifes artificiales ubicados en el Golfo de Morrosquillo, Caribe colombiano Tesis Biol Mar , Univ Jorge Tadeo Lozano, Bogotá. 107 p. [ Links ]
15 Jaimes, J., Y. Pinzón y C. Trujillo. 2004. Explorando alternativas de pesca responsable con la langosta espinosa en Santa Marta. Fundación Sila Kangama, Santa Marta. 80 p. [ Links ]
16 Martínez, O. 2005. Evaluación del asentamiento de estadios tempranos de peces en colectores flotantes tipo GuSi en la región de Santa Marta, Caribe colombiano. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá. 142 p. [ Links ]
17 Muñoz, B. y C. Obregón. 1994. Estructura y proceso de colonización de la comunidad de peces en un arrecife artificial en Isla Naval (P.N.N.C.R.) Caribe colombiano. Tesis Biol. Mar., Univ. Jorge Tadeo Lozano, Bogotá 110 p. [ Links ]
18 Olaya-Restrepo, J., M.C. Reyes-Nivia y A. Rodríguez-Ramírez. 2008. Ensamblajes de peces arrecifales del área de Santa Marta y el Parque Nacional Natural Tayrona. Bol. Invest. Mar. Cost., 37(1): 111-127. [ Links ]
19 Portier, K. M., G. Fabi y P. H. Darius. 2000. Study design and data analysis issues. 21-50. En: Seaman Jr., W. (Ed.). Artificial reef evaluation with application to natural marine habitats. CRC Press, Boca Ratón, EE.UU. 246 p. [ Links ]
20 Reyes-Nivia, C., A. Rodríguez-Ramírez y J. Garzón-Ferreira. 2004. Peces asociados a formaciones coralinas de cinco áreas del Caribe colombiano: listado de especies y primeros registros para las áreas. Bol. Invest. Mar. Cost., 33: 87-100. [ Links ]
21 Santodomingo, N. K., A. Gracia, A. Rodríguez, C. Reyes, J. Reyes, J. C. Márquez, N. Cruz, D. P. Flórez, D. Báez, F. Cortés, G. Borrero y M. C. Díaz. 2005. Biodiversidad asociada a las plataformas de explotación de gas natural (Campo Chuchupa: La Guajira, Colombia). Informe Técnico. INVEMAR-ECOPETROL S.A., Santa Marta. 225 p. [ Links ]
22 Seaman, Jr. W. y A. Jensen. 2000. Purposes and practices of artificial reef evaluation. 1-19. En: Seaman Jr., W. (Ed.). Artificial reef evaluation with application to natural marine habitats. CRC Press, Boca Raton, EE.UU. 246 p. [ Links ]
23 Zar, H. 1996. Biostatistical analysis. Tercera edición. Prentice-Hall, Upper Saddle River, EE.UU. 898 p. [ Links ]
FECHA DE RECEPCIÓN: 01/06/07 FECHA DE ACEPTACIÓN: 19/02/09
