










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.39 no.1 Santa Marta Jan./June 2010
EFECTO DE LA SEDIMENTACIÓN EN EL RECLUTAMIENTO DE LAS MACROALGAS DICTYOTA SPP. Y LOBOPHORA VARIEGATA: UN ESTUDIO EXPERIMENTAL EN EL PARQUE NACIONAL NATURAL TAYRONA, CARIBE COLOMBIANO*
EFFECTS OF SEDIMENTATION ON THE RECRUITMENT OF THE MACROALGAE DICTYOTA SPP. AND LOBOPHORA VARIEGATA: AN EXPERIMENTAL STUDY IN THE TAYRONA NATIONAL NATURAL PARK, COLOMBIAN CARIBBEAN
Lennin Flórez-Leiva1, Alejandro Rangel-Campo2, Martha Díaz-Ruiz3, Dagoberto E. Venera-Pontón2 y Guillermo Díaz-Pulido4
1 Universidad de Concepción, Departamento de Oceanografía, Chile. lflorez@udec.cl
2 Universidad del Magdalena, Instituto de Investigaciones Tropicales-INTROPIC, Carrera 32 No 22-08, Santa Marta, Colombia. ajrangelc@gmail.com (A.R.C.); dagovenera@gmail.com (D.E.V.P.)
3 Instituto de Investigaciones Marinas y Costeras-INVEMAR, A.A. 1016, Santa Marta, Colombia. mardiaz@invemar.org.co
4 Griffith University, Griffith School of Environment y ARC Centre of Excellence for Coral Reef Studies, Brisbane, Nathan 4111, QLD, Australia. g.diaz-pulido@griffith.edu.au
RESUMEN
La sedimentación está frecuentemente implicada como un factor clave en la degradación arrecifal. Sin embargo son pocos los estudios experimentales que evalúan este importante proceso en la dinámica del reclutamiento de las algas en los arrecifes coralinos. En un experimento de campo se manipularon los niveles de sedimentación y se examinaron los impactos en el reclutamiento y crecimiento de las macroalgas pardas Dictyota spp. y Lobophora variegata en dos bahías (Chengue y Granate) del Parque Nacional Natural Tayrona, Caribe colombiano. Se encontró una gran variabilidad de respuestas de las algas a la sedimentación, la cual dependió del nivel de sedimento empleado, tipo de alga estudiada y parámetro poblacional considerado (reclutamiento o crecimiento). La adición de sedimentos tuvo generalmente un efecto negativo en el reclutamiento y crecimiento de Dictyota spp. Contrariamente, la adición de sedimentos no afectó el reclutamiento de L. variegata, aunque la remoción de sedimentos sí lo hizo. La bahía con mayor tasa de sedimentación (Granate) presentó en general menores valores de reclutamiento y crecimiento de ambas algas que la otra (Chengue). Estos resultados indican que existe una gran complejidad y variabilidad en las respuestas de las algas arrecifales a los efectos de sedimentación. Sugieren también que la degradación arrecifal tiene el potencial de modificar la dinámica poblacional de los estadios tempranos de vida de las algas. Esto último puede tener implicaciones importantes para el entendimiento ecológico de la degradación arrecifal.
PALABRAS CLAVE: Caribe colombiano, Arrecifes coralinos, Macroalgas, Parque Nacional Natural Tayrona, Sedimentación.
ABSTRACT
Sedimentation is frequently associated with coral reef degradation. However, there are few experimental studies evaluating the impacts of this process on the dynamics of algal recruitment on coral reefs. In a field experiment, we manipulated the levels of sedimentation and examined the impacts on the recruitment and growth of the brown macroalgae Dictyota spp. and Lobophora variegata, in two localities (Chengue and Granate) on the Tayrona National Natural Park, Colombian Caribbean. We found considerable variability in the responses of algae to sedimentation. This variability depended on the levels of sediments used, the type of algae employed and the population parameter considered (recruitment or growth). Sediment addition generally had a negative effect on the recruitment and early growth of the alga Dictyota spp. In contrast, sediment addition did not affect the recruitment of L. variegata, while sediment removal had a negative effect on recruitment. The locality with higher sedimentation rates (Granate) generally had lower recruitment and growth of both algae than that of the other locality with lower sedimentation levels (Chengue). Our study indicates that there is substantial variability and complexity in the responses of reef algae to the effects of sedimentation. The results also suggest that the process of reef degradation can modify early population dynamics of benthic algae, with important implications for understanding the ecology of reef degradation.
KEY WORDS: Colombian Caribbean, Coral reefs, Macroalgae, Tayrona National Natural Park, Sedimentation.
INTRODUCCIÓN
La degradación de los arrecifes coralinos se ha incrementado considerablemente en los últimos años debido al deterioro de la calidad del agua (eutroficación e incrementos de sedimentación), sobrepesca, enfermedades de herbívoros y corales, blanqueamiento coralino producido por el calentamiento global y acidificación de las aguas oceánicas (Jackson et al., 2001; Hoegh-Guldberg et al., 2007). Estos procesos de degradación pueden conducir a la pérdida de corales duros y al incremento de las macroalgas, trayendo como consecuencia un cambio de estado ecológico en los arrecifes (Done, 1992; Hughes, 1994; McCook, 1999; Hughes et al., 2003).
Estos cambios de estado pueden ser negativos para el ecosistema, ya que reducen las tasas de acreción arrecifal y empobrecen la biodiversidad marina, además de traer consigo pérdidas económicas importantes (Hughes, 1994; Grimsditch y Salm, 2006). Los cambios de estado pueden además alterar la estructura de los ensamblajes algales, modificando la dominancia y diversidad algal. Al presentarse estas alteraciones se pueden afectar procesos importantes en los que participan las macroalgas como, por ejemplo, la productividad primaria, interacciones ecológicas con los corales y la acreción de los arrecifes coralinos (debido a la reducción de la abundancia de algas calcáreas). Además, pueden reducirse los hábitats para diversas especies de importancia ecológica y comercial (Jensen, 1993; Adey, 1998; Bula-Meyer, 2004).
La dinámica del reclutamiento de algas es un proceso crítico durante su colonización e invasión en los arrecifes coralinos. Para que ocurra la colonización algal se requiere que esporas y propágulos se asienten y recluten en el sustrato duro y que muchos juveniles sobrevivan hasta alcanzar los estadios adultos para sostener la dominancia. Las macroalgas pueden responder de manera diferente a las perturbaciones antropogénicas dependiendo de los estadios de vida. Algunos trabajos han evaluado los efectos de la sedimentación en las algas adultas (Eriksson y Johansson, 2003, 2005); sin embargo, los efectos de la sedimentación sobre los estadios tempranos de las macroalgas han recibido muy poca atención. A pesar de la importancia de las algas en la ecología arrecifal y del reclutamiento como proceso poblacional y ecológico, es muy poco lo que se conoce acerca de la dinámica del reclutamiento de algas en arrecifes de coral y los procesos y factores que la regulan (Díaz-Pulido y McCook, 2004).
Entre los procesos de gran importancia en la degradación arrecifal, a nivel local, se encuentra el incremento de la tasa de sedimentación marina, debida principalmente a las actividades humanas tales como agricultura, desarrollo costero, construcción, minería, dragado y deforestación. Estas actividades, especialmente la deforestación de los bosques para generar zonas ganaderas o agrícolas, colectar madera, realizar construcciones y producir carbón vegetal, modifican las propiedades físicas y químicas del suelo, posibilitando los procesos de erosión y un consecuente incremento de la sedimentación costera. La sedimentación constituye una preocupación importante porque afecta a un gran número de arrecifes coralinos y ha sido relacionada con su salud (Bruijnzeel, 1990; Rogers, 1990; Richardson, 1998). De esta forma, el objetivo de este estudio fue investigar los efectos de los incrementos de la sedimentación sobre el desarrollo temprano de las poblaciones de las algas Dictyota spp. y Lobophora variegata en dos bahías del Parque Nacional Natural Tayrona.
ÁREA DE ESTUDIO
Bahías de Chengue y Granate
Chengue es una pequeña bahía (3.3 km2 de superficie) localizada dentro del Parque Nacional Natural Tayrona (PNNT, Figura 1) a unos 14 Km al nororiente de la ciudad de Santa Marta, en la costa Caribe de Colombia (11°20'N-74°08'W). La topografía costera en el área es muy heterogénea y pendiente, debido a su cercanía con la Sierra Nevada de Santa Marta (Rodríguez-Ramírez y Garzón-Ferreira, 2003).
El clima y las condiciones oceanográficas son similares a la bahía de Santa Marta y están determinados por la influencia de los vientos Alisios del nororiente. La precipitación anual promedio para el área es alrededor de los 350 mm y el promedio anual de temperatura del aire es de 28 °C (Salzwedel y Müller, 1983); su clima puede calificarse como seco y cálido. La oceanografía del área está determinada por la presencia de aguas de surgencia costera, con temperaturas de hasta 21 °C durante la surgencia (diciembre-enero) y de hasta 31 °C durante el periodo de no surgencia, o lluvias (Zea, 1993; Díaz-Pulido y Garzón-Ferreira, 2002).
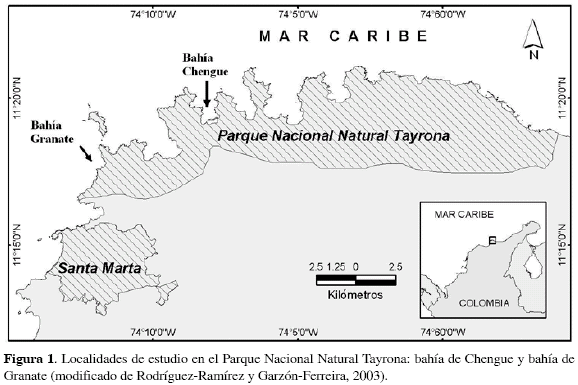
Por su parte, la bahía de Granate está localizada al noroeste de la ciudad de Santa Marta (Figura 1) a 11°18' N -74°12' W. Esta bahía es arenosa en algunas partes de su periferia y hasta los 25 m de profundidad presenta fondos coralinos (Erhardt y Werding, 1975). Esta bahía, aunque abierta al mar, está bien protegida de los fuertes vientos Alisios del nororiente y de la corriente norte por la isla de la Aguja; la cual ocasiona un oleaje tenue durante la mayor parte del año, a excepción del costado occidental que se encuentra más expuesto al impacto directo de los vientos Granate exhibe una profundidad que no sobrepasa los 30 m y, al igual que en otras bahías del PNNT, su parte central está constituida por fondos sedimentarios recientes (Acosta, 1989). Esta bahía se diferencia de Chengue por presentar una elevada tasa de sedimentación natural. En general existe un gradiente de sedimentación natural a lo largo del PNNT, siendo mayor en áreas cercanas a las fuentes de descarga (e. g. Santa Marta, debido a los ríos Manzanares y Magdalena) y decreciendo hacia el nororiente (Zea, 1994).
MATERIALES Y MÉTODOS
Macroalgas experimentales
Las macroalgas pardas Dictyota spp. y Lobophora variegata fueron utilizadas por las siguientes razones: 1) el arrecife de estudio provee abundante sustrato para un buen desarrollo de algas Dictyota spp, 2) se encuentra una alta cobertura de Dictyota spp. de 3 a 10 m de profundidad, 3) son macroalgas de fácil identificación taxonómica a nivel de género, caso de Dictyota spp. (De Clerck, 2003) y 4) ambas algas están implicadas en procesos de degradación arrecifal y existe poca información sobre las causas de su dominancia en el PNNT (Loya y Rinkevich, 1980; Díaz-Pulido y Garzón-Ferreira, 2002; Jompa y McCook, 2002). El género Dictyota es bastante diverso en el PNNT, con al menos 14 especies registradas para el área (Díaz-Pulido y Díaz-Ruiz, 2003). Si bien sería ideal comparar el reclutamiento de Dictyota a nivel de especies, esto es bastante difícil, dados los tamaños microscópicos de los propágulos algales y la falta de estructuras morfológicas y reproductivas únicas entre ellas.
Experimentos en campo
Se analizaron los efectos de la sedimentación en el desarrollo de reclutas de Dictyota spp. y L. variegata utilizando sustratos artificiales expuestos a diferentes niveles de sedimentos, a una profundidad entre los 6 y 9 m. Las algas fueron sembradas en sustratos artificiales, los cuales consistían en placas de cerámica (11 x 11 cm de lado, atornilladas a una estructura de hierro en el fondo). Se trabajaron tres niveles de sedimentación: 1) alta, 2) natural (control) y 3) remoción de sedimentos. Las manipulaciones experimentales se realizaron siguiendo metodologías descritas previamente (Devinny y Volse, 1978; Umar et al., 1998; Chapman y Fletcher, 2002; Eriksson y Johansson, 2003, 2005). El experimento comprendió dos fases: incubación y manipulación experimental.
Incubación
La incubación incluyó el asentamiento natural y crecimiento in situ de las esporas de las algas estudiadas para así disponer de un número de individuos que permitiera desarrollar el experimento La incubación se desarrolló entre abril y junio de 2005, cuando los picos de crecimiento y desarrollo algal fueron altos (e.g. Díaz-Pulido y Garzón-Ferreira, 2002), permitiendo capturar y obtener suficientes propágulos algales. La incubación se desarrolló en Chengue porque Dictyota spp. es abundante en esta bahía (Bula-Meyer, 1985; Díaz-Pulido y Garzón-Ferreira, 2002) Con el fin de determinar los efectos de la localidad en el impacto de la sedimentación sobre las macroalgas, luego del proceso de incubación, la mitad de las placas (15) fue trasladada a Granate y la otra mitad (15) permaneció en Chengue.
Manipulación experimental
Para determinar los efectos de la sedimentación se realizaron manipulaciones experimentales de sedimentos marinos, los cuales fueron adicionados o removidos de las placas. La cantidad de sedimentos adicionados se estandarizó teniendo en cuenta las tasas de sedimentación natural para el Parque Nacional Natural Tayrona. Para evitar que la composición del sedimento adicionado fuera diferente entre localidades, los sedimentos adicionados en ambas bahías fueron obtenidos del plano arenoso adyacente al talud arrecifal de Chengue, entre los 10 y 11 m de profundidad. Las adiciones se hicieron semanalmente durante 12 semanas. Las tasas de sedimentación en los arrecifes del Parque Tayrona son bastante variables, por ejemplo en Chengue se han registrado valores máximos promedio de 1.5 (Garzón-Ferreira, 1998), 4.5 (Garzón-Ferreira et al., 2002) y 14.4 mg/cm2/día (Solano, 1987) y en isla Aguja de 1.5 mg/cm2/día (Zea, 1994). Con el objeto de maximizar los efectos de la adición de sedimentos en nuestro experimento, se usó una tasa de sedimentación registrada para el Parque Tayrona Para determinar las tasas experimentales de sedimentación a ser empleadas por placa se multiplicaron las tasas de sedimentación natural (14.4 mg/cm2/día) por el área de la placa (100 cm2), por el número de días transcurridos entre una adición y otra (aproximadamente una semana). De esta manera se determinó que una tasa de sedimentación de 10 g de sedimento/semana/placa sería adecuada para el experimento. Esta tasa de sedimentación junto con la tasa natural de sedimentación, constituyen el tratamiento de sedimentación alta.
Los tratamientos de remoción de sedimentos consistieron en retirar las placas de las estructuras metálicas, invertirlas in situ y sacudirlas suavemente por un período estandarizado de 10 segundos; las placas fueron colocadas nuevamente en las estructuras metálicas. Se tuvo cuidado de no dañar los reclutas fijados en las placas. Los tratamientos naturales no fueron manipulados y se constituyeron en controles. Cada tratamiento consistió en cinco placas (réplicas). Al final del experimento, las placas con las algas fueron transportadas al laboratorio de la Universidad del Magdalena utilizando tanques plásticos de 60 L de capacidad, protegidos de la luz y en agua de mar permanente.
Variables respuesta
Las variables respuesta analizadas fueron altura, ancho, puntos de bifurcación y reclutamiento algal. Estas mediciones se hicieron en el laboratorio con cubreobjetos milimetrados. La variable reclutamiento para las dos macroalgas se registró como el número de reclutas que aparecieron en cada placa, exceptuando el borde de la misma (aproximadamente 1 cm). Esto último, para evitar el efecto potencial que puedan generar las características particulares del borde de la placa (efecto borde). La altura de Dictyota spp. se obtuvo como la máxima longitud alcanzada por el talo desde el disco de fijación hasta su parte más alta. El ancho se determinó como la distancia entre un extremo y otro del borde del talo en sentido perpendicular al largo; los puntos de bifurcación (PB, en el caso de Dictyota spp.) se definieron como aquellos donde la célula apical separa dos ramas contiguas entre sí. Los puntos de bifurcación dan una idea del número de dicotomías de la planta y se estudian como una medida de crecimiento. La morfometría realizada se basó en De Clerck (2003). Para L. variegata la medición del ancho fue definida como el diámetro del crecimiento lateral del talo (Díaz-Pulido y McCook, 2004).
Análisis de datos
Los datos fueron analizados mediante Análisis de Varianza (ANOVA) bifactoriales con niveles de sedimentación (adición, control y remoción) y áreas de estudio (Chengue y Granate) como factores fijos, y placas como réplicas (n= 5). La altura, ancho y puntos de bifurcación fueron medidos en 20 reclutas algales que fueron seleccionados aleatoriamente para cada placa y luego se calculó el valor promedio por placa para cada variable. Estos valores constituyeron los datos usados (N= 30: 5 réplicas x 3 tratamientos x 2 localidades) en los análisis estadísticos. El reclutamiento y las otras variables de crecimiento se utilizaron como respuesta en los ANOVA. Los datos fueron inspeccionados para revisar los supuestos del ANOVA, incluyendo la homoscedasticidad (prueba de Cochran) y la normalidad de residuales (Underwood, 1997).
RESULTADOS
Efectos de la sedimentación sobre el reclutamiento y crecimiento de Dictyota spp.
Se observó una gran variabilidad de respuestas del alga Dictyota spp. ante la sedimentación, la cual dependió del parámetro poblacional considerado (altura, ancho o reclutamiento) y del nivel de sedimentación (adición o remoción). La adición de sedimentos en Granate redujo a cero el número de reclutas de Dictyota spp., mientras que en Chengue el reclutamiento fue cuatro veces menor, aproximadamente, con respecto al tratamiento control. A pesar de estas claras diferencias, el ANOVA no detectó diferencias significativas. Por su parte, la localidad afectó significativamente el reclutamiento y crecimiento del alga. El crecimiento de Dictyota fue mayor en Chengue que en Granate (p= 0.002; Figuras 2B y C), así como los puntos de bifurcación (probabilidad marginal, p= 0.057); el ancho se vio levemente reducido en Granate, aunque la diferencia no fue significativa (p= 0.110; Tabla 1; Figura 2B).
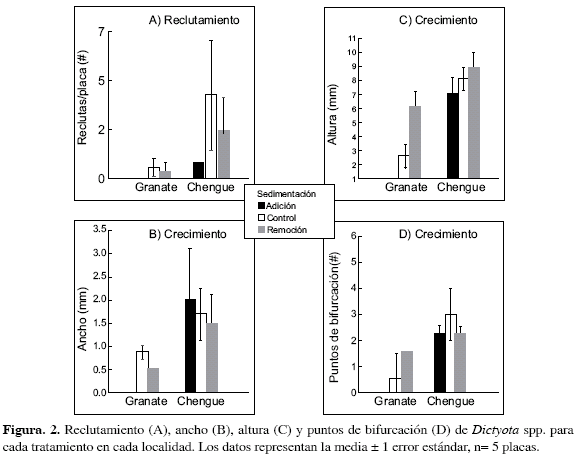
La remoción de sedimentos incrementó ligeramente la altura de Dictyota spp. en ambas bahías (p= 0.065, Tabla 1, Figura 2C). También se observó que el efecto de la remoción de sedimentos fue mayor en Granate (Figura 2B) que en Chengue. Por su parte, Chengue presentó mayores valores de crecimiento en altura (p= 0.002, Tabla 1). Por otra parte, los puntos de bifurcación no presentaron diferencias significativas entre los tratamientos de sedimentación, aunque esta variable presentó mayores valores en Chengue (p= 0.057; Tabla 1, Figura 2D).
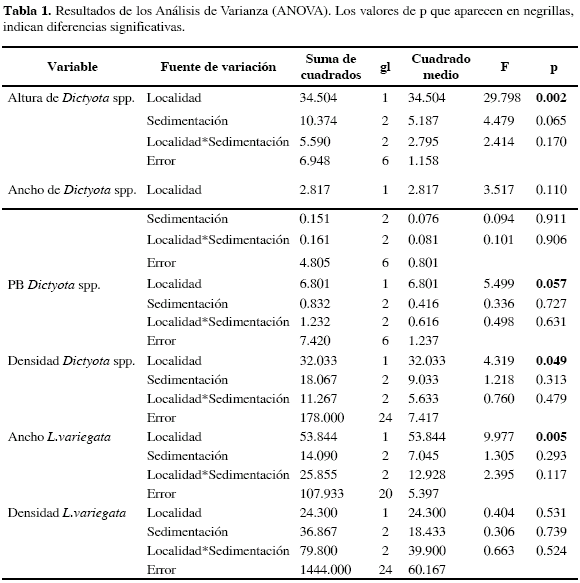
La localidad afectó significativamente el reclutamiento de Dictyota spp, presentándose mayores valores en Chengue (p= 0.049, Tabla 1, Figura 2). Como se mencionó anteriormente, se presentaron menores valores de reclutamiento entre tratamientos de adición que en los de remoción y controles, aunque la diferencia no fue significativa, sugiriendo que pudo haber un efecto negativo de la sedimentación en el reclutamiento de Dictyota.
Efectos de la sedimentación sobre el crecimiento y reclutamiento de Lobophora variegata
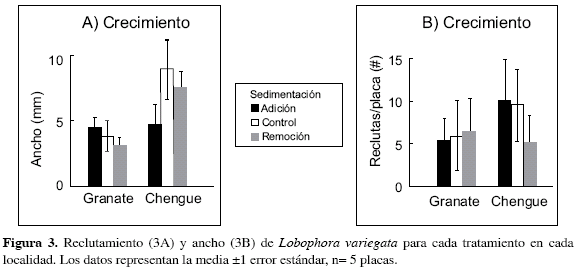
Lobophora variegata tuvo una respuesta leve a los tratamientos de sedimentación. En Chengue, la remoción de sedimentos redujo el reclutamiento a la mitad, con respecto a los tratamientos control o de adición de sedimentos (Figura 3B). Sin embargo, esta diferencia no fue significativa. En Granate no se observaron diferencias en el reclutamiento de L. variegata entre los tratamientos de sedimentación. Por otra parte, sí se observaron diferencias significativas entre las dos bahías con respecto al ancho de los talos de L. variegata, siendo éstos mayores en Chengue que en Granate (p= 0.005; Tabla 1, Figura 3A).
DISCUSIÓN
La manipulación de los niveles de sedimentación tuvo una respuesta variada en las macroalgas Dictyota spp. y L. variegata. La intensidad del efecto de la sedimentación dependió de la localidad, cantidad de sedimentos adicionados (o removidos), además del tipo de alga estudiada y del parámetro poblacional considerado. Esto ilustra la complejidad y variabilidad que existe entre los parámetros poblacionales de Dictyota spp. y L. variegata y su relación con los procesos que afectan su ecología. Por lo tanto, es importante considerar la medición de distintas variables para lograr un mejor entendimiento de los efectos de los procesos ecológicos en los organismos
Efectos de la sedimentación y la localidad en el reclutamiento macroalgal
Los dos grupos de algas respondieron diferente a los efectos de los niveles de sedimentación empleados. A pesar de la baja significancia estadística, los resultados sugieren que existe una tendencia a la disminución del crecimiento (altura) y reclutamiento de Dictyota spp. en respuesta al incremento de niveles de sedimentación. Es posible que la corta duración de los experimentos (tres meses) o la limitada replicación (n= 5), hayan generado un error estadístico Tipo II (Underwood, 1997), en donde los análisis estadísticos muestran una baja significancia a pesar de que el efecto existe. Los resultados de Dictyota son, sin embargo, consistentes con estudios previos, que han demostrado que los incrementos en las cargas de sedimentos en los arrecifes de coral son perjudiciales para las algas adultas (Umar et al., 1998). Estudios experimentales en zonas templadas han demostrado también que altos niveles de sedimentos disminuyen el reclutamiento de Macrocystis (Neushul et al., 1976), impiden la fijación de propágulos en M. pyrifera (Devinny y Volse, 1978) y la fijación de estados microscópicos de varios kelpos como Undaria pinnatifida y Ecklonia cava (Arakawa y Matsuike, 1992). Espinoza y Rodríguez (1987) también encontraron una reducción en las tallas del talo y la capacidad reproductiva de Sargassum sinicola sujeta a incrementos en los niveles de sedimentos en el golfo de California. Resultados similares se han observado en corales, pastos marinos y manglares, reduciendo su reclutamiento, cobertura, establecimiento, supervivencia y tasas de crecimiento (Pringle, 1989; Hodgson, 1990; Rogers, 1990; Umar et al., 1998; Airoldi, 2003; Schiel et al., 2006). Este estudio demuestra por primera vez efectos negativos de la sedimentación sobre los estadios tempranos de macroalgas de arrecifes coralinos
Se han postulado cuatro mecanismos principales mediante los cuales, los incrementos de sedimentos afectan las algas marinas (Airoldi, 2003). 1) Asfixia: implica una reducción de la disponibilidad y calidad de la luz y oxígeno generando potencialmente la acumulación de sulfuro de hidrógeno y desechos de productos metabólicos, cuyo efecto produce cambios en las características químicas del microambiente 2) Reducción del acceso a nutrientes esenciales: posiblemente debido a alteraciones en las capas de intercambio ("boundary layers", Larkum et al, 2003) que afectan la difusión e intercambio de nutrientes 3) Abrasión: causada por el movimiento de sedimentos que puede dañar y remover el alga o sus partes. 4) Cambios en las características físicas de la superficie del fondo: ocurre como consecuencia del reemplazo de sustratos duros estables por partículas inestables y puede resultar en una pérdida de hábitat para el establecimiento algal (Devinny y Volse, 1978; D'Antonio, 1986; Airoldi et al., 1996; Airoldi, 1998). Este trabajo no pretendió examinar cómo estos mecanismos afectan las algas, sin embargo las observaciones sugieren que es posible que todos estos mecanismos operen en conjunto para reducir el reclutamiento y crecimiento del alga Dictyota. Es importante resaltar que se requieren más estudios que profundicen sobre los mecanismos mediante los cuales los sedimentos afectan las algas marinas
La adición de sedimentos no afectó el reclutamiento de L. variegata, contrario a lo observado en Dictyota spp. Este resultado es sorprendente porque generalmente se tiende a asociar el incremento de sedimentación con el deterioro o daño de las poblaciones algales (como se detalló en el caso de Dictyota). Por otro lado, existe evidencia de que algunas algas pueden ser resistentes a la sedimentación (DvAntonio, 1986; Kendrick, 1991). Trabajos en algas rojas muestran que existe una resistencia a la abrasión causada por arena y muestran que estas algas pueden prosperar en áreas donde pocas especies sobreviven (DvAntonio, 1986). Otros trabajos en campo registran que la presencia de limo podría ser benéfica al suministrar nutrientes y proteger las algas del movimiento del agua (Kennelly, 1983; Schaffelke, 1999). Sin embargo, la adición de sedimentos redujo el crecimiento de L. variegata (Figura 3B) y por tanto es posible que el mecanismo de aumento de disponibilidad de nutrientes no opere en estas circunstancias particulares
Estos resultados enfatizan la variabilidad de respuestas encontradas en las poblaciones algales ante los efectos de la sedimentación. La razón de la falta de respuestas en el reclutamiento de L. variegata a la adición de sedimentos no es clara, pero es posible que los sedimentos puedan proteger de una u otra manera al alga del pastoreo de herbívoros, por ejemplo creando una barrera física especialmente contra mesoherbívoros (Kennelly, 1983). Esto puede ser particularmente cierto si se considera que en el tratamiento de remoción de sedimentos, el reclutamiento de L. variegata se redujo en más de la mitad. Sin embargo, la hipótesis de la protección de sedimentos contra el pastoreo necesita verificación experimental. Finalmente, la tolerancia de L. variegata a niveles de sedimentación puede conferir cierto grado de ventaja en hábitats o situaciones (e.g. cambios de estado de corales a algas) con niveles de sedimentación moderados o altos.
Los impactos de la sedimentación sobre las algas variaron entre localidades (arrecifes) expuestas a diferentes grados de sedimentación natural, con valores menores de reclutamiento y crecimiento algal en el arrecife con mayores tasas de sedimentación (Granate). Áreas como Granate y la bahía de Santa Marta, que presentan un mayor grado de impacto antropogénico (al encontrarse cercanas a áreas urbanas, turísticas y por estar cerca a fuentes de descarga continental como ríos y desagües de aguas residuales), son aparentemente menos aptas para el crecimiento de algas pardas rastreras, como es el caso de Dictyota y L. variegata, que áreas con menores tasas de sedimentación (e.g. Chengue). Algas pardas de talos erectos considerables, como por ejemplo Sargassum, sin embargo proliferan en Granate pero son escasas en los arrecifes de Chengue. Diversos factores pueden también jugar papeles claves en determinar las distribuciones espaciales de algas en la región del PNNT, e.g. tasas de pastoreo (Díaz-Pulido et al, 2007), disponibilidad de nutrientes, grado de deterioro del arrecife, intensidad de la surgencia (Bula- Meyer, 1985). Nuestro experimento demuestra que el tipo y magnitud del impacto de los sedimentos es altamente heterogéneo en escalas espaciales, contribuyendo a la complejidad espacial de las comunidades algales de la región del PNNT. Este estudio indica que existe una gran complejidad y variabilidad en las respuestas de las algas arrecifales a los efectos de la escorrentía de sedimentos desde la Sierra Nevada de Santa Marta. Sugiere también que los procesos de degradación arrecifal pueden modificar la dinámica poblacional y estadios tempranos de vida de las algas bentónicas, lo cual puede tener implicaciones importantes en el entendimiento de la ecología de la degradación arrecifal.
AGRADECIMIENTOS
Este estudio fue financiado por el Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología-Colciencias, el Fondo Patrimonial para la Investigación de la Universidad del Magdalena-Fonciencias, y el Instituto de Investigaciones Marinas y Costeras-INVEMAR. Agradecemos a Olga Camacho, Katrín Perilla, Sonia Bejarano, Érika Ortíz, Isabel Chacón, Oscar Martínez y Adriana Santacruz por su ayuda en campo. Igualmente a Laura Airoldi, Bernabé Santelices, Sven Zea, Adolfo Sanjuán y a dos revisores anónimos por sus comentarios constructivos
BIBLIOGRAFÍA
1 Acosta, L. 1989. Composición y estructura de las comunidades de corales hermatípicos en tres zonas representativas de la región de Santa Marta, Punta Betín, isla Morro Grande y ensenada de Granate, mar Caribe colombiano. Tesis de Biol. Univ. Nacional de Colombia, Bogotá. 242 p. [ Links ]
2 Adey, W. H. 1998. Coral reefs: algal structured and mediated ecosystems in shallow, turbulent, alkaline waters. J. Phycol., 34: 393-406. [ Links ]
3 Airoldi, L. 1998. Roles of disturbance, sediment stress, and substratum retention on spatial dominance in algal turf. Ecology, 79: 2759-2770. [ Links ]
4 Airoldi, L. 2003. The effects of sedimentation on rocky coast assemblages. Oceanogr. Mar. Biol. Ann. Rev., 41: 161-236. [ Links ]
5 Airoldi, L., M. Fabiano y F. Cinelli. 1996. Sediment deposition and movement over a turf assemblage in a shallow rocky coastal area on the Ligurian Sea. Mar. Ecol. Prog. Ser., 133: 241-251. [ Links ]
6 Arakawa, H. y K. Matsuike. 1992. Influence on insertion of zoospores, germination, survival, and maturation of gametophytes of brown algae exerted by sediments. Nippon Suisan Gakkaishi, 58: 619-625. [ Links ]
7 Bruijnzeel, L. 1990. Hydrology of moist tropical forest and effects of conversion: a state of the knowledge. UNESCO, París 224 p. [ Links ]
8 Bula-Meyer, G. 1985. Un núcleo nuevo de surgencia en el Caribe colombiano detectado en correlación con las macroalgas. Bol. Ecotrópica, 12: 2-26. [ Links ]
9 Bula-Meyer, G. 2004. Las macroalgas marinas en la agronomía y el uso potencial de Sargassum flotante en la producción de fertilizantes en el archipiélago de San Andrés y Providencia, Colombia Rev Intrópica, 1: 91-103. [ Links ]
10 Chapman, A. S. y R. L. Fletcher. 2002. Differential effects of sediments on survival and growth of Fucus serratus embryos (Fucales, Phaeophyceae). J. Phycology, 38: 894-903. [ Links ]
11 D'Antonio, C. M. 1986. Role of sand in the domination of hard substrata by the intertidal alga Rhodomela larix. Mar. Ecol. Prog. Ser., 27: 263-275. [ Links ]
12 De Clerck, O. 2003. The genus Dictyota (Dictyotales, Phaeophyta) in the Indian Ocean. Opera Bot., Bélgica, 13: 1-205. [ Links ]
13 Devinny, J. S. y L. A. Volse. 1978. Effects of sediments on the development of Macrocystis pyrifera gametophytes. Mar. Biol., 48: 343-348. [ Links ]
14 Díaz-Pulido, G. y M. Díaz-Ruiz. 2003. Diversity of benthic marine algae of the Colombian Atlantic. Biota Colomb 4: 203-246. [ Links ]
15 Díaz-Pulido, G. y J. Garzón-Ferreira. 2002. Seasonality in algal assemblages on upwelling-influenced coral reef in the Colombian Caribbean. Bot. Mar., 45: 284-292. [ Links ]
16 Díaz-Pulido, G. y L. McCook. 2004. Effects of live coral, epilithic algae communities and substrate type on algal recruitment Coral Reefs, 23: 225-233. [ Links ]
17 Díaz-Pulido, G., L. Villamil y V. Almanza. 2007. Herbivory effects on the morphology of the brown alga Padina boergesenii (Phaeophyta). Phycologia, 46: 131-136. [ Links ]
18 Done, T. J. 1992. Phase shifts in coral reef communities and their ecological significance Hydrobiología, 247: 121-132. [ Links ]
19 Erhardt, H. y B. Werding. 1975. Los corales (Antozoa e Hydrozoa) de la ensenada de Granate, pequeña bahía al este de Santa Marta, Colombia. Caldasia, 11 (53): 107-138. [ Links ]
20 Eriksson, B. K. y G. Johansson. 2003. Sedimentation reduces recruitment success of Fucus vesiculosus (Phaeophyceae) in the Baltic Sea. Eur. J. Phycol., 38: 217-222. [ Links ]
21 Eriksson, B K y G Johansson 2005 Effects of sedimentation on macroalgae: species-specific responses are related to reproductive traits Oecologia, 143: 438-448. [ Links ]
22 Espinoza, J. y H. Rodríguez. 1987. Seasonal phenology and reciprocal transplantation of Sargassum sinicola Setchell et Gardner in the southern Gulf of California. J. Exp. Mar. Biol. Ecol., 110: 183195. [ Links ]
23 Garzón-Ferreira, J. 1998. Bahía de Chengue, Parque Nacional Natural Tayrona, Colombia 115-125 En: CARICOMP (Ed.). Caribbean coral reef, seagrass and mangrove sites. UNESCO, París. 345 p. [ Links ]
24 Garzón-Ferreira, J., A. Rodríguez-Ramírez, S. Bejarano-Chavarro, R. Navas-Camacho y C. Reyes- Nivia. 2002. Estado de los arrecifes coralinos. 29-40. En: Ospina-Salazar, G. H. y A. Acero P. (Eds.). Informe del estado de los ambientes marinos y costeros en Colombia: año 2001. Serie de Publicaciones Periódicas No 8, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta 292 p. [ Links ]
25 Grimsditch, G. y R. V. Salm. 2006. Coral reef resilience and resistance to bleaching IUCN, Gland, Suiza 53 p. [ Links ]
26 Hodgson, G. 1990. Tetracycline reduces sedimentation damage to corals. Mar. Biol., 104: 493-496. [ Links ]
27 Hoegh-Guldberg, O., P. J. Mumby, A. J. Hooten, R. S. Steneck, P. Greenfield, E. Gómez, C. D. Harvell, P. F. Sale, A. J. Edwards, K. Caldeira, N. Knowlton, C. M. Eakin, R. Iglesias-Prieto, N. Muthiga, R. H. Bradbury, A. Dubi y M. E. Hatziolos. 2007. Coral reefs under rapid climate change and ocean acidification. Science, 318: 1737-1742. [ Links ]
28 Hughes, T. P. 1994. Catastrophes, phase shifts and large-scale degradation of a Caribbean coral reef. Science, 265 (5178): 1547-1551. [ Links ]
29 Hughes, T. P., A. H. Baird, D. R. Bellwood, M. Card, S. R. Connolly, C. Folke, R. K. Grosberg, O. Hoegh- Guldberg, J. B. C. Jackson, J. Kleypas, J. M. Lough, P. A. Marshall, M. Nystrom, S. R. Palumbi, J. M. Pandolfi, B. R. Rosen y J. Roughgarden. 2003. Climate change, human impacts, and the resilence of coral reefs. Science, 301: 929-933. [ Links ]
30 Jackson, J. B. C., M. X. Kirby, W. H. Berger, K. A. Bjorndal, L. W. Botsford, B. J. Bourque, R. H. Bradbury, R. Cooke, J. Erlandson, J. A. Estes, T. P. Hughes, S. Kidwell, C. B. Lange, H. S. Lenihan, J. M. Pandolfi, C. H. Peterson, R. S. Steneck, M. J. Tegner y R. R. Warner. 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science, 293: 629-638. [ Links ]
31 Jensen, A. 1993. Present and future needs for algae and algal products. Hydrobiologia, 260-261: 15-23. [ Links ]
32 Jompa, J. y L. J. McCook. 2002. The effects of nutrients and herbivory on competition between a hard coral (Porites cylindica) and a brown algae (Lobophora variegata). Limnol. Oceanogr., 47: 527- 534. [ Links ]
33 Kendrick, G. A. 1991. Recruitment of coralline crusts and filamentous turf algae in the Galapagos archipelago: effect of simulated scour, erosion and accretion. J. Exp. Mar. Biol. Ecol, 147: 47-63. [ Links ]
34 Kennelly, S. J. 1983. An experimental approach to the study of factors affecting algal colonization in a sublittoral kelp forest. J. Exp. Mar. Biol. Ecol., 68: 257-276. [ Links ]
35 Larkum, A. W. D., E. C. Koc. y M. Kuehl. 2003. Diffusive boundary layers and photosynthesis of the epilithic algal community of coral reefs. Mar. Biol., 142: 1073-1082. [ Links ]
36 Loya, Y. y B. Rinkevich. 1980. Effect of oil pollution on coral reef communities. Mar. Ecol. Prog. Ser., 3: 176-180. [ Links ]
37 McCook, L. J. 1999. Macroalgae, nutrient and phase shifts on coral reefs: scientific issues and management consequences for the Great Barrier Reef. Coral Reefs, 18: 357-367. [ Links ]
38 Neushul, M., M. S. Foster, D. A. Coon, J. W. Woessner y B. W. Harquer. 1976. An in situ study of recruitment, growth and survival of subtidal marine algae: techniques and preliminary result. J. Phycol., 12: 397-408. [ Links ]
39 Pringle, A. W. 1989. The history of dredging in Cleveland Bay, Queensland and its effect on sediment movement and on growth of mangrove, coral and seagrass. Great Barrier Reef Marine Park Autority, Townsville, Queensland, Australia. 177 p. [ Links ]
40 Richardson, L. L. 1998. Coral diseases: what is really known? Trends Ecol. Evol., 13: 438-443. [ Links ]
41 Rodríguez-Ramírez, A. y J. Garzón-Ferreira. 2003. Monitoreo de arrecifes coralinos, pastos marinos y manglares en la bahía de Chengue (Caribe colombiano) 1993-1999. Series de Publicaciones Especiales No 8, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta. 170 p. [ Links ]
42 Rogers, C. S. 1990. Responses of coral reefs and reefs organisms to sedimentation. Mar. Ecol. Prog. Ser., 62: 185-202. [ Links ]
43 Salzwedel, H. y K. Müller. 1983. A summary of meteorological data for the bay Santa Marta, Colombian Caribbean. An. Inst. Invest. Mar. Punta de Betín, 13: 67-83. [ Links ]
44 Schaffelke, B. 1999. Particulate organic matter as an alternative nutrient source for tropical Sargassum species (Fucales, Pheophyceae). J. Phycol., 35: 1150-1157. [ Links ]
45 Schiel, D. R., S. A. Wood, R. A. Dunmore y D. Taylor. 2006. Sediment on rocky intertidal reefs: effects on early post-settlement stages of habitat-forming seaweeds. J. Exp. Mar. Biol. Ecol., 331: 158-172. [ Links ]
46 Solano, O. 1987. Estructura y diversidad de las comunidades de corales hermatípicos en la bahía de Chengue (PNNT). Tesis M. Sc., Univ. Nacional de Colombia, Bogotá. 111 p. [ Links ]
47 Umar, M. J., L. J. McCooke e I. R. Price. 1998. Effects of sediment deposition on the seaweed Sargassum on a fringing reef. Coral Reefs, 17: 169-177. [ Links ]
48 Underwood, A. J. 1997. Experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge University Press, Cambridge, Reino Unido. 503 p. [ Links ]
49 Zea, S. 1993. Cover of sponges and other sessile organisms in rocky and coral reef habitats of Santa Marta, Colombian Caribbean Sea. Caribb. J. Sci., 29 (1-2): 75-88. [ Links ]
50 Zea, S. 1994. Patterns of coral and sponge abundance in stressed coral reefs at Santa Marta, Colombian Caribbean. 257-264. En: Van Soest, R. W. M., T. M. G. Van Kempen y J. C. Braekman (Eds.). Sponges in time and space. Proceedings of the 4th International Porifera Congress. Ámsterdam. 515 p. [ Links ]
FECHA DE RECEPCIÓN: 16/02/2007 FECHA DE ACEPTACIÓN: 19/03/2010
Contribución No. 1047 del Instituto de Investigaciones Marinas y Costeras INVEMAR
