










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.39 no.1 Santa Marta Jan./June 2010
COBERTURA DE LOS FONDOS Y PAISAJES BENTÓNICOS ASOCIADOS A FORMACIONES DIAPÍRICAS EN LOS BANCOS DE SALMEDINA, PLATAFORMA CONTINENTAL DEL CARIBE COLOMBIANO
SEAFLOOR COVERAGE AND BENTHIC LANDSCAPES ASSOCIATED TO DIAPIRIC (MUD VOLCANISM) FORMATIONS IN SALMEDINA BANKS, COLOMBIAN CARIBBEAN CONTINENTAL SHELF
Juan Gabriel Domínguez1, Juan Carlos Gómez1, Constanza Ricaurte1, Gisela Mayo1, Jaime Orejarena1, Juan Manuel Díaz2 y Carlos Alberto Andrade1
Escuela Naval de Cadetes "Almirante Padilla", Grupo de Investigación en Oceanología (GIO), Cartagena, Colombia. candrade@enap.edu.co
Corporación para Investigaciones Biológicas, Unidad de Biodiversidad, Medellín, Colombia. jdiaz@cib.org.co
RESUMEN
Se determinaron las unidades ecológicas submarinas alrededor de un volcán activo de lodo en los bancos de Salmedina, Caribe colombiano, y se cartografiaron mediante video-transectos submarinos georreferenciados, análisis de la batimetría e interpretación de imágenes satelitales. Se describen la distribución, la cobertura relativa y los organismos bentónicos sésiles más representativos de las diferentes unidades ecológicas. La zona somera (0-10 m) de los bancos presenta 60 % de cobertura algal y 10 % de coral vivo; en las zonas más profundas (<10 m) la cobertura de coral vivo varía entre 33 % en los cantiles semiverticales y 55 % en las cimas de los bancos menos elevados; las especies de coral dominantes fueron Siderastrea siderea y Montastraea annularis en todos los sectores. Los bancos de Salmedina reciben la influencia cada vez mayor de plumas turbias que probablemente han aumentado la sedimentación, y las comunidades biológicas se ven posiblemente afectadas por el empleo de explosivos para la pesca. Aunque la zona más somera del banco Salmedina es la que evidencia mayor afectación, las comunidades coralinas más profundas aún mantienen una cobertura viva significativa en comparación con otras zonas similares en la región. La actividad diapírica reciente con emanaciones constantes de lodo es determinante en la estructura de la comunidad coralina en el sector occidental.
PALABRAS CLAVE: Coberturas fondo marino, Procesos kársticos, Formaciones diapíricas, Caribe colombiano.
ABSTRACT
The submarine ecological units around an active mud volcano in the offshore banks of Salmedina, Colombian Caribbean, were determined and mapped employing geo-referenced submarine video-transects, bathymetry analysis, and interpretation of satellite imagery. Distribution, relative cover, and the most representative benthic, sessile organisms of the different ecological units are described. In the shallow zone of the banks, 60 % of the substrate was covered by algae and only 10 % by living corals. The relative cover of living corals in deeper zones increased to 33 % in semi vertical slopes and up to 55 % on the upper flats of deeper located banks. The dominant coral species overall were Siderastrea siderea and Montastraea annularis. A determining natural factor in the reef structure in the zone seems to be recent diapiric activity, with a constant mud emanation in the western sector. The Banks of Salmedina are under the influence of turbid plumes that seemingly increase sedimentation and are impacted by blast fishing as well. Although the shallowest zone of the bank shows the highest affectation, the coral community in the deeper zones still exhibit a noticeable cover of living coral, even higher than in other areas in the region with similar features. The recent diapiric activity of constant mud ejection determines the coral community structure in the western sector.
KEY WORDS: Seafloor coverage, Karstic processes, Diapiric (mud volcanic) formations, Colombian Caribbean.
INTRODUCCIÓN
La importancia ecológica de los arrecifes coralinos es reconocida a nivel mundial debido a su biodiversidad y elevada producción biológica. El progresivo y avanzado deterioro generalizado de estos ecosistemas, especialmente en el Caribe, debido a múltiples factores, entre los que se destacan la sobrepesca, la sedimentación, el enriquecimiento por nutrientes (eutrofización) y el calentamiento global (Burke y Maidens, 2004), ha convertido en una prioridad los estudios sobre el estado y distribución de los arrecifes de coral, principalmente para los países caribeños.
La generación de mapas ecológicos o de coberturas a partir de muestreos in situ y con ayuda de imágenes satelitales y fotografías aéreas, utilizados anteriormente para valorar ecosistemas terrestres, se ha implementado con gran éxito para el mapeo de áreas coralinas (Díaz, 1998). En Colombia se han realizado varios trabajos utilizando este tipo de metodología (Monsalve y Restrepo, 1990; Sánchez et al., 1992; Díaz et al., 1996a, 1996b, 1997, 2000), particularmente en áreas conocidas y de gran extensión, dejando de lado formaciones menos visitadas pero no de menor importancia ecológica.
Localizados en la parte central de la plataforma continental del Caribe colombiano, los bancos de Salmedina, al igual que los demás bajos e islas característicos del área, son producto de una progresiva colonización coralina sobre bajos de origen diapírico (Vernette, 1985), y constituyen el límite norte de un cordón arrecifal que se extiende por unos 200 km en sentido NNE-SSW hasta Isla Fuerte (Geister, 1983; Díaz et al., 1996a). Los bancos de Salmedina ocupan una extensión aproximada de 10 km2, de los cuales 4 km2 corresponden a áreas propiamente coralinas (Ricaurte et al., 2004), prácticamente desconocidas pero de gran importancia para las comunidades de pescadores de la región.
La presencia de actividad diapírica de lodo en estos bancos los hace particularmente interesantes, ya que esta condición les confiere características especiales y los constituye en un escenario privilegiado para el estudio de este fenómeno y su influencia sobre las comunidades biológicas, especialmente las coralinas. Hasta el momento, el interés por los bancos de Salmedina se ha centrado en el riesgo que representan para la navegación, ya que se localizan en la trayectoria de las rutas de acceso al puerto de Cartagena de Indias, uno de los de mayor tráfico en el Caribe colombiano (Ricaurte et al., 2004). No obstante, aparte de una descripción somera por Díaz et al. (2000), no han sido objeto de estudios detallados. El objetivo de este trabajo es describir la composición, la distribución y el estado de las formaciones coralinas de los bancos de Salmedina como un aporte a su conocimiento y un insumo para la implementación de medidas de manejo tendientes a la conservación de este sector poco conocido del Caribe colombiano.
MATERIALES Y MÉTODOS
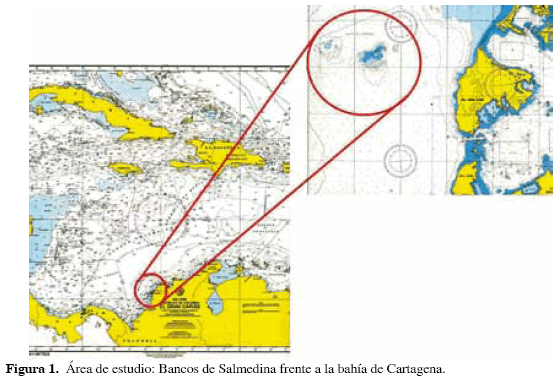
Los bancos de Salmedina se localizan entre las latitudes 10°21'15'' y 10°23'10''N y las longitudes 75°38'05'' y 75°41'45'' W, aproximadamente a 7 km al W de la punta norte de la isla de Tierra Bomba, frente a la ciudad de Cartagena. (Figura 1). Están conformados por cinco elevaciones, a saber: banco Salmedina, banco Burbujas, banco Ukura, banco Kubina y banco Ygio, que suman una extensión de 10.5 km2, de los cuales 4.45 km2 corresponden a las plataformas más altas (Ricaurte et al., 2004).
Durante 2004 se llevaron a cabo 17 transectos de video sobre los bancos de Salmedina, cuya longitud varió entre 240 y 960 m, trazados intentando cubrir homogéneamente el área de estudio; trece de ellos fueron dispuestos en sentido sur-norte siguiendo la corriente predominante en la zona, con el fin de economizar tiempo y energía, y los cuatro restantes se trazaron en sentido este-oeste para cubrir las zonas intermedias. Los videos se realizaron mediante buceo autónomo empleando una cámara digital (Panasonic 3CCD, PV-GS70®) dentro de un recipiente estanco (Equinos PROPAC 6-GS70®). Durante las filmaciones, la cámara se mantuvo siempre aproximadamente a una distancia de 2 m del fondo, con la lente perpendicular a éste; para mantener la dirección constante a lo largo del transecto, un buzo acompañante guiaba al camarógrafo unos metros adelante de éste empleando una brújula. Los transectos fueron georeferenciados usando un boyarín en superficie, equipado con un GPS (Garmín MAP76®), que registraba la posición aproximadamente cada 30 segundos; un tercer buzo, en superficie, se encargaba de mantener el boyarín perpendicular sobre el camarógrafo para registrar datos precisos de posición (Figura 2).
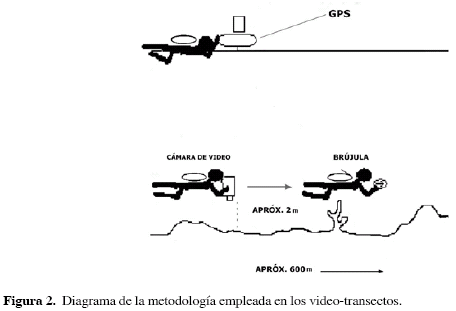
Los datos obtenidos en los transectos se complementaron con 42 inmersiones en apnea. En cada punto de inmersión se anotó el tipo de sustrato y la posición geográfica; los puntos georreferenciados, tanto de los video-transectos como en las observaciones complementarias, fueron superpuestos sobre un mapa batimétrico de la zona a escala 1:12500, utilizando el programa computacional MapInfo Professional 5®.
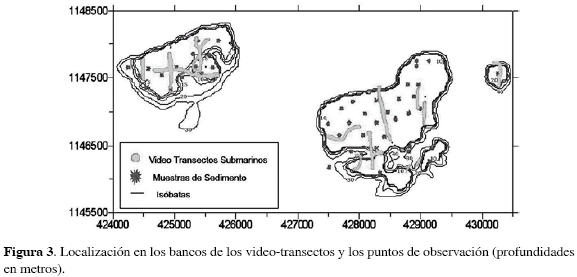
Las filmaciones fueron examinadas imagen por imagen cada 30 segundos, hasta obtener toda la secuencia a lo largo del video-transecto y cada una fue proyectada sobre el mapa batimétrico, incorporando los datos de posición registrados con ayuda del programa MapInfo Professional 7.0® (Figura 3). Se elaboró un mapa de coberturas detallado del área con base en los cambios topográficos y su relación con las variaciones en el tipo de cobertura detectado en las imágenes y en las observaciones hechas in situ.
El mapa de coberturas fue sobrepuesto a una imagen del satélite SPOT de diciembre 20 de 1990, utilizando el programa Surfer 5®. Con ello, al detectarse variaciones en la coloración del fondo atribuibles a cambios en la cobertura, inferidos por los registros de las filmaciones y distintos a los producidos por los cambios de profundidad, se complementó la información obtenida mediante interpolación de los video-transectos y los muestreos puntuales.
Los tipos de cobertura se determinaron con base en las unidades ecológicas de áreas coralinas propuestas por Díaz et al. (2000) para permitir la comparación de la información con la existente para otras áreas del Caribe colombiano. No obstante, dicha clasificación fue complementada adicionando nuevas unidades ecológicas y subdividiendo otras según características morfológicas propias del área de estudio. Para la identificación de los organismos más representativos de cada unidad ecológica, se examinaron las filmaciones y se ubicaron cinco puntos al azar sobre la pantalla; la filmación fue detenida cada 30 s y se anotó el tipo de sustrato u organismo marcado en cada uno de los puntos; en promedio se obtuvieron unos 200 puntos por transecto; según lo permitió la resolución del video, se anotó hasta el menor nivel taxonómico posible y se calcularon las abundancias relativas.
El procesamiento de la imagen satelital se efectuó mediante el programa ENVI (Environment for Visualizing Images) versión 3.5® y fue georreferenciada a partir de una imagen ya corregida, usando la técnica de "registro imagen-imagen" y empleando el algoritmo bilinear para el ajuste de los puntos de control mediante translación y rotación de la imagen. Las principales características, bandas espectrales y resolución de la imagen se presentan en la Tabla 1. La clasificación multiespectral se realizó en forma no supervisada por "clases". El sistema agrupó los píxeles con características espectrales similares, permitiendo clasificar posteriormente los tipos de cobertura, y se agruparon las propiedades espectrales similares de los píxeles.
Con base en la batimetría de alta resolución levantada para la zona (Andrade et al., 2005) se elaboró una imagen "batimétrica", que fue transformada hasta una resolución espacial de 15 m, coincidente con la de la imagen satelital. Posteriormente se extrajo una ventana correspondiente al área de los bancos de Salmedina y se utilizó la imagen batimétrica para construir una máscara que descartó los valores de píxeles superiores a 40 m de la imagen satelital, con lo cual se resaltaron los píxeles correspondientes a los bancos y se eliminó el ruido introducido por las aguas turbias circundantes que pudiera alterar la eficiencia de los algoritmos de clasificación. Para esta clasificación no supervisada se utilizó el algoritmo de agrupación que aplica la técnica de análisis de datos auto- organizados "ISODATA" (Iterative Self-Organizing Data Analysis Technique). Se calcularon los valores promedio de cada clase que estaban distribuidos dentro de todos los datos e iterativamente se agruparon los píxeles sobrantes mediante la técnica de la mínima distancia Cada iteración recalcula el valor medio y reclasifica los píxeles con respecto al nuevo valor medio (Jensen, 1996).
Con la clasificación obtenida se identificaron las zonas donde las clases son similares y se hizo una comparación espacial con los sitios donde se identificaron determinado tipo de cobertura respetando los valores de profundidad similar ISODATA representa un completo y equilibrado grupo de procesos heurísticos que han sido incorporados en un algoritmo de clasificación iterativa Muchos de esos procesos son el resultado de experiencia ganada a través de la experimentación y es auto-organizativo porque requiere relativamente poca información previa dada por el usuario.
RESULTADOS
Las observaciones realizadas y la interpretación de las imágenes satelitales revelaron que los bancos de Salmedina poseen una inusitada diversidad de paisajes; se obtuvo un mapa de coberturas del área que discrimina 12 unidades ecológicas diferentes (Figura 4) que difieren entre sí en el tipo de cobertura dominante o en características morfológicas relevantes que permiten adoptarlas como unidades independientes; los rasgos predominantes de cada unidad se describen en la Tabla 2. Las zonas menos profundas se encuentran sobre la meseta del banco Salmedina, el mayor de los cinco que componen el área de estudio, que alcanza profundidades entre 4 y 8 m en su parte más somera; la meseta está cubierta en su mayoría por sustrato calcáreo colonizado por algas verdes, algas calcáreas y macroalgas verdes y pardas, principalmente Dictyota sp. con un 60 % del área total. Sobreviven colonias muy aisladas de corales masivos (Montastraea annularis, Siderastrea siderea) que conforman menos del 10 % de la cobertura (Figura 5A). En algunos sectores de la meseta el sustrato se eleva 2 m sobre el fondo, formando terrazas surcadas por canales en los que se acumula arena y escombros pequeños de coral; la cobertura en las crestas de las terrazas es similar al tapete de algas del resto de la meseta, con una dominancia clara de las algas (70 %) sobre el coral (Figura 5B), aunque en los surcos sobreviven algunas colonias grandes de corales masivos y octocorales.
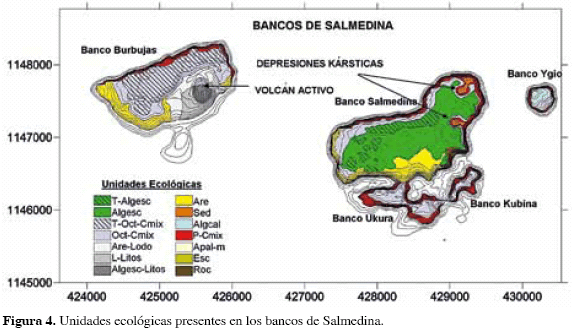
El cantil norte de Salmedina y Burbujas y el cantil alrededor de Ukura, Kubina e Ygio poseen pendientes muy pronunciadas, con paredes semi-verticales que caen varios metros y cuya cobertura coralina corresponde principalmente a colonias costrosas de Montastraea sp. que cubren en promedio un 33 % de la totalidad del sustrato; algunos antipatarios, frondosas macroalgas verdes y grandes esponjas barril (Xetospongia muta) complementan el paisaje (Figura 5C). En contraste con el sector norte, los sectores sur y occidental del cantil de los bancos Burbujas y Salmedina carecen de cobertura viva. Se trata de un sustrato arenoso donde el elemento predominante, casi en un 100 %, es arena fina con algunos parches de coral que desaparecen rápidamente en los primeros metros de profundidad (Figura 5D). Al sur del banco Salmedina la arena da paso a un sustrato totalmente cubierto por escombros de corales ramificados, posiblemente Porites porites, que cubren el cantil hasta varios metros de profundidad (Figura 5E).
El banco Salmedina presenta dos formaciones morfológicas muy especiales y puntuales. La primera se encuentra en el sector suroccidental del cantil donde el sustrato está conformado por rocas de diversos tamaños, dando la apariencia de un "derrumbe" de grandes proporciones (unidad Roc); algunas rocas han sido colonizadas por algas, hidrocorales (Millepora sp.) y esponjas, pero en general el desarrollo coralino es muy bajo en este sector. La otra formación consiste en dos depresiones que se ubican en el sector nororiental de la meseta del Banco; su contorno es circular, de unos 150 m de diámetro y 10 m de profundidad, con paredes muy inclinadas que poseen una cobertura similar a la encontrada en el cantil norte (unidad Cmix); el fondo plano de las depresiones está cubierto por una gruesa capa de sedimentos finos y lodosos, y acumulaciones de octocorales muertos agrupados hacia el centro; los únicos organismos vivos evidentes en esta zona se hallan en un tapete irregular de algas filamentosas que crecen sobre el fango (Figura 5F).

El área con comunidades coralinas vivas cubre un total de 1.5 km2 de las plataformas superiores de los bancos; los bancos del sur, Kubina y Ukura, están dominados por un paisaje de octocorales, principalmente del género Plexaura, que crecen entre formas de corales masivos y costrosos (Diploria sp., Montastraea sp., S. siderea), cubriendo en promedio 55 % del sustrato; también se encuentran representadas varias especies de algas calcáreas, dominando Halimeda sp., esponjas y algas incrustantes; las macroalgas frondosas ocupan menos de 20 % del sustrato (Figura 6A). Las zonas más elevadas en estos dos bajos están conformadas por setos muertos de Acropora palmata que cubren 55 % del sustrato; en algunos sectores se desarrollan parches de colonias costrosas de Montastraea spp., colonias masivas de S. siderea y coral de fuego Millepora sp sobre los esqueletos de A. palmata; la cobertura de algas en esta zona es muy baja, inferior a 5 % (Figura 6B).
La meseta del bajo Ygio está dividida en dos sectores: el sector sur, un poco más profundo, está dominado por corales y octocorales, un ambiente similar al de los bajos Ukura y Kubina (unidad Oct-Cmix), pero con la característica especial de presentar una densidad considerable de esponjas barril (X. muta). El sector norte de la meseta está dominado totalmente por terrazas de origen coralino cubiertas casi en su totalidad por un tapete continuo de algas calcáreas como Halimeda sp. y Amphiroa sp., que cubren 50 % del total del sustrato; el resto de la cobertura es compartida por algas incrustantes, Millepora sp. y esponjas incrustantes (Figura 6C). Este tapete sólo es interrumpido por canales profundos entre terrazas donde se acumulan restos de algas calcáreas que cubren el fondo y en sus paredes prosperan corales de formas foliáceas, masivas y costrosas (Agaricia sp., Montastraea spp. y Siderastrea sp.) y diversas especies de octocorales.
La meseta superior del banco Burbujas presenta una clara división en dos paisajes definidos y muy diferentes. El sector norte corresponde a una unidad ecológica muy similar, en composición y cobertura de coral vivo, a los bancos Kubina y Ukura, pero se diferencia de éstos porque el sustrato se eleva hasta 2 m por encima del fondo formando terrazas que están surcadas por valles arenosos, casi siempre orientados en sentido norte-sur. Sobre las mesetas se desarrollan "jardines" de octocorales de varios géneros que crecen sobre tapetes coralinos de formas costrosas, foliáceas y masivas, Halimeda sp., esponjas de varios tipos, algas incrustantes, algas pardas (Dyctiota sp) y Millepora sp. ; la cobertura de coral vivo es de 40 %, contra 20 % de coral muerto cubierto por algas (Figura 6D).
Hacia el sur del banco Burbujas la zona de terrazas termina abruptamente dando paso a un valle que domina todo el sector sur del banco; el sustrato en el fondo del valle está totalmente cubierto por una capa de lodo fino y lodolitos pequeños, con una aparente ausencia de organismos sésiles (Figura 6E). Hacia la periferia del valle se encuentra una franja de transición, entre la zona coralina y la zona de lodos, donde el sustrato es de arena blanca dispuesta en rizos (ripple marks) paralelos al oleaje en cuyos surcos se acumula lodo y algunos lodolitos; octocorales muy dispersos crecen fijados sobre lodolitos grandes o fragmentos coralinos desprendidos de las terrazas (Figura 6F).

En el centro del valle, en medio de la planicie de lodo, se eleva un domo de 8 m de altura, cuyas laderas presentan dos sectores claramente definidos, uno al norte, dominado por terrazas de coral similares a las que dominan todo el sector norte del banco, aunque un poco más dispersas, y el otro en las secciones occidental, oriental y sur del domo, que están cubiertas por lodolitos de diferentes tamaños, la mayoría colonizados por algas calcáreas. Hacia el norte del área se encuentran escombros de coral entre los lodolitos (Figura 6G). En la porción más elevada de la cresta la actividad diapírica es evidente y se manifiesta con un flujo constante de lodo; los lodos finos en suspensión enturbian el agua dificultando la observación clara del paisaje. Sin embargo, a pesar de esta limitación, se puede afirmar que el sustrato no cambia sustancialmente, siguiendo dominado por rocas de lodo; se logró observar dos octocorales, posiblemente Plexaura sp., sobreviviendo en estas condiciones.
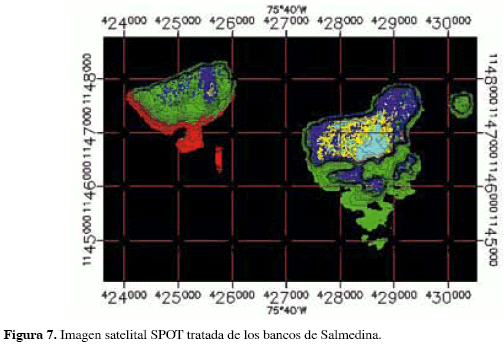
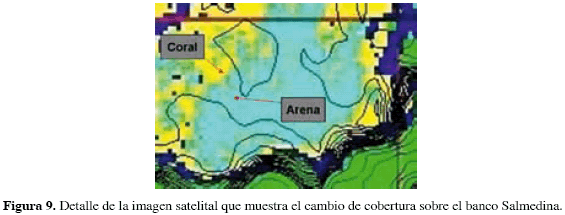
El análisis de la imagen satelital reveló la presencia de cinco diferentes coloraciones agrupando los píxeles por su grado de reflectancia (Figura 7); los cambios de coloración correspondieron primordialmente a variaciones en la profundidad (desde amarillo en las zonas más someras entre 4 y 5 m de profundidad hasta rojo, por debajo de 30 m.). La imagen fue usada para aumentar la resolución de la batimetría en los primeros 10 m de profundidad, ya que permitió ver detalles mucho más finos de la morfológica de la zona y corregir los contornos generados por la interpolación de los datos batimétricos (Figura 8). La imagen también resultó útil para diferenciar cambios de cobertura en ambientes someros y muy contrastantes, siempre que la profundidad se mantenía con escasa variación y que observaciones de campo permitían reconocer el tipo de sustrato mostrado por la imagen (Figura 9); gracias a ello se logró delimitar el parche de arena sobre el banco Salmedina, el cual, debido a su gran extensión, presentaba dificultades para ello a partir de las observaciones in situ.
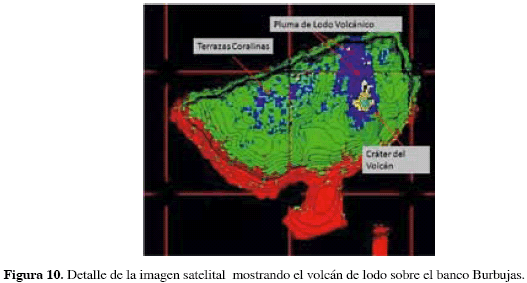
Por otro lado, la panorámica general que ofrece la imagen satelital permitió identificar y ubicar accidentes importantes en el área de estudio, como el volcán de lodo del banco Burbujas o su pluma de lodo (Figura 10), haciendo posible comprender la magnitud del fenómeno diapírico observado in situ. En general, la imagen satelital permitió dar mayor detalle al mapa de coberturas incrementando la resolución de la batimetría y brindando la posibilidad de definir contornos a gran escala de algunos rasgos submarinos a poca profundidad. Debido a que el área de estudio está rodeada por fondos profundos, la caracterización con mayor detalle de éstos a partir de la imagen se hace muy difícil; en estudios realizados en ambientes más someros, la definición de las coberturas desde imágenes satelitales es mucho más detallada (Genet, 1992).

DISCUSIÓN
Estudios recientes en los bancos de Salmedina (Orejarena et al., 2004; Ricaurte et al., 2004) muestran que éstos presentan diferencias muy marcadas entre sí en morfología, profundidad, nivel de la actividad diapírica y grado de exposición al tren de olas y a plumas de agua continental. Esta diversidad de factores favorece el desarrollo de múltiples biotopos en un área relativamente pequeña.
El aporte de aguas continentales superficiales, cargadas de nutrientes y sedimentos, favorecen el crecimiento algal y afectan negativamente el desarrollo coralino (Alvarado y Corchuelo, 1992); aportes de este tipo, provenientes principalmente del Canal del Dique, un tributario artificial del río Magdalena, llegan a los bancos de Salmedina en forma de parches y plumas turbias (Orejarena et al., 2004). Estas masas de agua turbia son muy superficiales y pueden explicar la dominancia de macroalgas en la cobertura en las áreas menos profundas de los bancos, entre 4 y 8 m, donde la cobertura algal alcanza valores de hasta 70 % y las coberturas de coral vivo no sobrepasan 10 %. Por otro lado, la presencia de grandes extensiones de escombros de P. porites y setos colapsados de A. palmata muestran que hasta tiempos recientes la cobertura de coral vivo era considerable pero que resultó severamente afectada aparentemente por la pesca con explosivos, una práctica frecuente en la región (Díaz et al., 2000).
El arrastre continuo de sedimentos y escombros coralinos causado por las fuertes corrientes superficiales, que predominantemente van hacia el sur, determinan en buena parte el paisaje submarino al sur de Salmedina. Éste consiste principalmente de acumulaciones de escombros coralinos y arena desprovistos de cobertura coralina. En contraste, la meseta de los bancos Ukura y Kubina, ubicados a menos de 100 m del banco Salmedina, presentan una cobertura de coral vivo considerable (55 %), superior a la registrada en áreas cercanas similares con mayor desarrollo arrecifal, tales como bajo Tortuga (40 %) y bajo Bushell (31 %) (Díaz et al., 2000). El principal factor que puede explicar esta diferencia es un profundo cañón que alcanza 38 m de profundidad y que separa estos bajos del Salmedina. Este último podría estar actuando a barlovento como una barrera que mantiene protegido el sector de sotavento; sin embargo, algunas especies hermatípicas importantes ya han desaparecido, a juzgar por la abundancia de escombros y setos muertos de A. palmata sobre las crestas superiores en esta área.
La influencia de las plumas de agua turbia disminuye notablemente en los primeros 7 a 8 m de la columna de agua; por debajo de 9 m el agua permanece clara la mayor parte del año, lo que favorece el desarrollo coralino en este sector. Las mesetas de los bajos Ygio y Burbujas exhiben una cobertura de coral vivo de 40 %, aunque la dominancia de especies como S. siderea y M. annularis, particularmente resistentes a la sedimentación y exitosas en la competencia con otras especies coralinas (Díaz et al., 1996b; Ferriz-Domínguez y Horta-Puga, 2001) y la presencia de amplias praderas de algas calcáreas, que suelen desarrollarse sobre el sustrato duro dejado por los escombros de coral (Frydl, 1982), muestran que estas formaciones están sometidas a variaciones fuertes de ciertas condiciones ambientales, especialmente de las tasas de sedimentación.
El alto grado de exposición al oleaje de fondo (swell) proveniente predominantemente del nor-noreste, propicia un mayor crecimiento coralino vertical a barlovento, creando una mayor elevación del andamiaje arrecifal y pendientes pronunciadas (Ricaurte et al., 2004); este patrón se repite en cada uno de los bajos de Salmedina, al igual que en otras estructuras arrecifales de la plataforma continental del Caribe colombiano (Díaz, 1998). La cobertura del talud norte de los bancos, cubierta por formas costrosas de coral, esponjas tubulares y antipatarios, es típica de este tipo de paisaje con alto grado de pendiente y escasa iluminación (Sánchez y Ortiz, 1992; Díaz, 1998).
De acuerdo con Ricaurte et al. (2004), el origen del valle que domina el sur del bajo Burbujas podría estar relacionado con el desplazamiento de material diapírico al formarse un domo descrito por Vernette (1985).La estructura prominente que se observa en medio del valle es muy probablemente de origen diapírico, pues actualmente se encuentra en actividad; en la cima de la prominencia existe una emanación constante de lodo en un área de unos 50 m2 acompañada de una copiosa salida de burbujas de gas en varios sectores y la formación de una "nube" turbia de varias decenas de metros. El flujo continuo de lodo que se solidificó sobre los flancos oriental, occidental y sur del domo evitan el desarrollo de formaciones coralinas; la dominancia de algas costrosas en este ambiente sugiere una alta resistencia de estos organismos a perturbaciones ambientales y a la turbidez. En contraste, el flanco norte está menos expuesto al flujo de lodo, y la fuerte corriente predominante del norte la excluye de la influencia de los sedimentos finos, propiciando el desarrollo coralino.
El resto del valle consiste en sustrato cubierto por una capa de lodos finos desprovista de organismos epibentónicos, debido posiblemente al aporte continuo de lodo proveniente de las estructuras diapíricas vecinas (Ricaurte et al., 2004). Hacia los límites del valle, donde la influencia del lodo disminuye, el paisaje cambia abruptamente a terrazas de jardines de octocorales sobre terrazas de coral, donde se concentra gran cantidad de peces; ello sugiere la existencia de un efecto secundario de la actividad diapírica en la comunidad arrecifal, pero hace falta mayor información que sustente esta apreciación.
Las dos conspicuas depresiones circulares que se encuentran en el bajo Salmedina son evidentemente de origen kárstico. Al igual que en los complejos arrecifales de las islas del Rosario, el bajo Tortuga y el archipiélago de San Bernardo, estas estructuras debieron originarse durante una de las glaciaciones del Pleistoceno, como resultado de la disolución de las formaciones calcáreas pre-existentes, al quedar éstas expuestas a la acción de las aguas de lluvia, creándose un intrincado sistema de cavernas; posteriormente, al elevarse nuevamente el nivel de mar, inundando las estructuras, los techos de las cavernas colapsaron en algunas zonas produciendo hondas depresiones que moldean el actual paisaje de los arrecifes (Díaz et al., 2000; López-Victoria y Díaz, 2000).
Para otros bajos arrecifales de la plataforma continental del Caribe colombiano se han descrito características similares a las halladas en el bajo Burbujas. Por ejemplo, Díaz et al (1996b) registraron en el bajo Bushnell una notoria disminución de la cobertura coralina hacia la mitad sur del bajo, cambiando de un ambiente coralino a uno dominado por arenas, esponjas y octocorales. Asimismo, Díaz et al. (2000) mencionaron la presencia de aguas de aspecto lechoso en las hondonadas del banco Tortuga, atribuidas a la resuspensión de sedimentos muy finos. La nueva evidencia sobre la influencia que ejercen las emanaciones de lodo sobre las coberturas y comunidades epibentónicas aledañas, sumada a las analogías registradas en otras áreas arrecifales de origen y condiciones similares, permiten sugerir que el diapirismo submarino de lodo es un factor generalizado importante en la configuración de la composición y la estructura de las comunidades coralinas a lo largo del cordón de formaciones coralinas y complejos arrecifales de la región central del Caribe colombiano.
AGRADECIMIENTOS
Los autores desean agradecer a la Dra. Gladys Bernal, Universidad Nacional de Colombia Sede Medellín, por su interés, sus comentarios para el manuscrito así como por su colaboración y la de su grupo en la toma de imágenes submarinas. Al Sr. José Ladeut por su gran pericia en el timón de las embarcaciones. Al Sr. Capitán de Navío Mauricio Ospina, director del CIOH, por su disposición para apoyar todos los esfuerzos necesarios para ir al mar. Este proyecto se llevó a cabo con el apoyo logístico de la Armada Nacional a través de la Escuela Naval "Almirante Padilla", la Dirección General Marítima y sus unidades en Cartagena, y de COLCIENCIAS en la financiación del proyecto "Estudio oceanográfico integral de los bancos de Salmedina, Caribe colombiano". Código: 112609-12399.
BIBLIOGRAFÍA
1 Alvarado, E. M. y M. C. Corchuelo. 1992. Los nutrientes, la temperatura y la salinidad provenientes del Canal del Dique como factores de deterioro en el Parque Nacional Natural Corales del Rosario (Cartagena, Colombia). 277-287. En: CCO (Ed.). Mem. VIII Sem. Nal. Cienc. Tecn. Mar., Santa Marta. 608 p. [ Links ]
2 Andrade, C. A., G. Bernal, C. Ricaurte, G. Mayo, J. Domínguez, J. Orejarena, A. Gutiérrez y W. Castro. 2005. Estudio oceanográfico de los bancos de Salmedina, Caribe colombiano. Escuela Naval Almirante Padilla, Cartagena. 113 p. [ Links ]
3 Burke, L. y J. Maidens. 2004. Reefs at risk in the Caribbean. World Resources Institute, Washington. 81 p. [ Links ]
4 Díaz, J. M. 1998. Maps for Colombian reefs. Reef Encounter, 12: 11-12. [ Links ]
5 Díaz, J. M., J. A. Sánchez, S. Zea y J. Garzón-Ferreira. 1996a. Morphology and marine habitats of two southwestern Caribbean atolls: Albuquerque and Courtown. Atoll Res. Bull., 435: 1-33. [ Links ]
6 Díaz, J. M., J. A. Sánchez y G. Díaz-Pulido. 1996b. Geomorfología y formaciones arrecifales recientes de Isla Fuerte y Bajo Bushnell, plataforma continental del Caribe colombiano. Bol. Invest. Mar. Cost., 25: 87-105. [ Links ]
7 Díaz, J. M., J. A. Sánchez y J. Geister. 1997. Development of lagoonal reefs in oceanic reef complexes of the southwestern Caribbean: geomorphology, structure and distribution. Proc. 8th Internat. Coral Reef Symp., Panamá, 1: 779-784. [ Links ]
8 Díaz, J. M., L. M. Barrios, M. H. Cendales, J. Garzón-Ferreira, J. Geister, M. López-Victoria, G. H. Ospina, F. Parra-Velandia, J. Pinzón, B. Vargas-Angel, F. A. Zapata y S. Zea. 2000. Áreas coralinas de Colombia Serie Publicaciones Especiales No 5, Instituto de Investigaciones Marinas y Costeras- INVEMAR, Santa Marta. 175 p. [ Links ]
9 Ferriz-Domínguez, N. y G. Horta-Puga. 2001. Short-term aggressive behavior in scleractinian corals from La Blanquilla reef, Veracruz Reef System. Rev. Biol. Trop., 49 (1): 67-75. [ Links ]
10 Frydl, P. M. 1982. Holocene ostracods in the southern Boso Peninsula. Mobil Oil Canada, Calgary, Canada. Bull. Univ. Tokio, 20, 10 p. [ Links ]
11 Geister, J. 1983. Holozäne westindische Korallenriffe: Geomorphologie, Ökologie und Fazies, Facies, 9: 173-284. [ Links ]
12 Genet, P. Y. 1992. Informe final proyectos cartografía fondos submarinos islas del Rosario (Colombia) y apreciación de sus áreas, a partir de imágenes de satélite SPOT. Bol. Cient. CIOH, 11: 15-26. [ Links ]
13 Jensen, J. R. 1996. Introductory digital image processing. Segunda edición, Prentice Hall, New Jersey. 450 p. [ Links ]
14 López-Victoria, M. y J. M. Díaz. 2000. Morfología y estructura de las formaciones coralinas del archipiélago de San Bernardo, Caribe colombiano. Rev. Acad. Col. Cienc., 24 (91): 219-230. [ Links ]
15 Monsalve, C. B. y J. D. Restrepo. 1990. Aproximación a la fotointerpretación y cartografía de los ecosistemas arrecifales, Isla Caribarú, Caribe colombiano. 325-336. En: CCO (Ed.). Mem. VII Sem. Nal. Cienc. Tecn. Mar, Cali. 792 p. [ Links ]
16 Orejarena, J. A., J. G. Domínguez, C. Ricaurte, G. Mayo, C. A. Andrade, H. M. Ospina y W. Castro. 2004. Variaciones de la concentración de la clorofila-a y su relación con los parámetros físicos medidos en los bancos de Salmedina durante 2003 - 2004, Caribe colombiano. Bol. Cient. CIOH, 22: 56-63. [ Links ]
17 Ricaurte, C., J. G. Domínguez, G. Mayo, C. A. Andrade, H. M. Ospina y A. de J. Gutiérrez. 2004. Nota sobre algunos rasgos geomorfológicos de los bancos de Salmedina, Bol. Cient. CIOH, 22: 64-76. [ Links ]
18 Sánchez, J. A. y V. Ortiz. 1992. Distribución de los corales negros (Antipatharia) de arrecifes entre Cartagena y el golfo de Urabá, Caribe colombiano. 362-369. En: CCO (Ed.). Mem. VIII Sem. Nal. Cienc Tecn Mar, Santa Marta 608 p. [ Links ]
19 Sánchez, J. A., A. Ramírez y R. Quintero. 1992. Estudio de reconocimiento de la morfología y las comunidades bénticas de los arrecifes de isla Tesoro, Parque Nacional Natural Corales del Rosario (PNNCR), Caribe colombiano 263-276 En: CCO (Ed.). Mem. VIII Sem. Nal. Cienc. Tecn. Mar, Santa Marta. 608 p. [ Links ]
20 Vernette, G. 1985. La plateforme continentale Caraïbe de Colombie (du Débouché du Magdalena au Golfe de Morrosquillo), importance du diapirisme argileux sur la morphologie et la sédimentation. Tesis Doctor de Estado en Ciencias, Univ. de Bordeaux I, Bordeaux, Francia. 381 p. [ Links ]
FECHA DE RECEPCIÓN: 07/10/2009 FECHA DE ACEPTACIÓN: 16/04/2010
