










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.39 no.2 Santa Marta July/Dec. 2010
EFECTOS DE LA PESCA SOBRE LA BIOLOGÍA REPRODUCTIVA DE LA LISA MUGIL INCILIS (PISCES: MUGILIDAE) EN LA CIÉNAGA GRANDE DE SANTA MARTA, CARIBE COLOMBIANO*
EFFECTS OF FISHING ON THE REPRODUCTIVE BIOLOGY OF THE MULLET MUGIL INCILIS (PISCES: MUGILIDAE) IN THE CIÉNAGA GRANDE DE SANTA MARTA, COLOMBIAN CARIBBEAN
Danetcy Mármol Rada1, Efraín Vitoria Maestre1 y Jacobo Blanco Racedo2
1 Instituto de Investigaciones Marinas y Costeras-INVEMAR, A.A. 1016, Cerro Punta Betín, Santa Marta, Colombia. danetcy_marmol@invemar.org.co (D.M.), eavitoria@invemar.org.co (E.V.)
2 Universidad del Magdalena, Programa de Ingeniería Pesquera, Santa Marta, Colombia. jablarce@gmail.com
RESUMEN
La lisa Mugil incilis representa uno de los recursos más importantes en la Ciénaga Grande de Santa Marta (CGSM). Con base en un monitoreo de las estadísticas de pesca colectadas en los períodos 1994-1996 y 2000-2007 y de aspectos biológicos de la especie entre 2006 y 2007, se evaluaron los efectos de la pesca artesanal sobre la biología reproductiva de M. incilis. Las estadísticas de pesca evidenciaron que la captura y tallas desembarcadas han disminuido en el tiempo. El pico de desove de la especie ocurrió en noviembre y diciembre cuando el índice gonadosomático y el factor de condición presentaron sus valores máximos, mientras la salinidad del agua disminuyó por la época de lluvias. De una muestra de 1954 individuos, se estimó que la proporción macho: hembra fue 1:3 y la talla media de madurez (TMM) fue 23.8 cm de longitud total, inferior a las estimadas previamente. Mugil incilis representó el 30 % de la captura anual de pescado en la CGSM entre 1994-2007, con 65 % de los individuos por debajo de la TMM y capturados principalmente con atarrayas en la época de desove. La menor frecuencia de capturas de peces de mayor tamaño parece explicar la disminución de la TMM. No se observaron evidencias para atribuir dicho decrecimiento a alguna estrategia de la especie. Se discuten y sugieren medidas para el manejo de esta pesquería.
PALABRAS CLAVE: Mugil incilis, Reproducción, Pesquería artesanal, Ciénaga Grande de Santa Marta, Caribe colombiano.
ABSTRACT
The mullet Mugil incilis is one of the most important fish resources in the Ciénaga Grande de Santa Marta. Based on monitoring of fishery statistics collected in the periods 1994-1996 and 2000-2007 and biological aspects of the species between 2006 and 2007, we evaluated fishing effects on the reproductive biology of M. incilis. Fishery statistics revealed that catch and fish size declined with time. Peak spawning was observed to occur between November - December, when the gonadosomatic index and condition factor showed their maximum values, contrasting with water salinity that plunged in the rainy period. Male: female ratio was estimated as 1:3 after a sample of 1954 individuals, and mean maturity size (MMS) calculated as 23. 8 cm total length (TL), lower than that previously reported. Mullet represented 30 % of the whole fish catch in the period 1994-2007, and 65 % of the individuals were found below their MMS, mainly caught with cast-nets at spawning time. Lower occurrence of larger fish in the catch seems to be a reasonable explanation for the reduction of MMS. No evidence was found to attribute this decrease to a species strategy. Management measures for this fishery are discussed and suggested.
KEY WORDS: Mugil incilis, Reproduction, Small-scale fishery, Ciénaga Grande de Santa Marta, Colombian Caribbean.
INTRODUCCIÓN
La Ciénaga Grande de Santa Marta (CGSM) conforma un sistema lagunar costero que posee una gran diversidad de especies de animales y vegetales, debido a que ofrece condiciones propicias para su establecimiento. Entre ellas, la disponibilidad de alimento para muchas especies, debido al intercambio de materia y energía con el río Magdalena, los ríos que bajan de la Sierra Nevada de Santa Marta y el mar Caribe. Esta dinámica hace de la CGSM un ecosistema con condiciones excepcionales para la reproducción de muchas especies de vertebrados e invertebrados, e incluso para su refugio y desarrollo (Wiedemann, 1973; Santos-Martínez y Acero, 1991). Por estas razones, durante décadas la CGSM se ha convertido en una fuente importante de alimento y sustento para sus pobladores, a través de una pesquería artesanal que juega un papel determinante en la economía de estas comunidades (Rueda y Defeo, 2003). Entre los principales recursos que se explotan se encuentran los peces, moluscos y crustáceos Las principales especies de la pesquería son la lisa (Mugil incilis), la mojarra rayada (Eugerres plumieri), el chivo mapalé (Cathorops mapale), el chivo cabezón (Ariopsis sp.), el sábalo (Megalops atlanticus), el macabí (Elops saurus), la mojarra lora (Oreochromis niloticus) y el coroncoro (Micropogonias furnieri) (Santos-Martínez y Acero, 1991) Entre esas especies M. incilis sobresale en la pesquería (Perdomo, 1973; Blanco, 1980; Sánchez et al., 1998; Rueda y Defeo, 2003), principalmente porque se ha mantenido como uno de los recursos más abundantes a lo largo del tiempo a pesar de los cambios ambientales producto de los efectos de origen antropogénicos y naturales ocurridos en este ecosistema (Botero y Salzwedel, 1999; Blanco et al., 2006).
La presión pesquera, los cambios tecnológicos en los artes de pesca y el efecto de algunos factores ambientales han conllevado a cambios en la dinámica de los recursos pesqueros y se han evidenciado signos de sobreexplotación en algunas especies como M. incilis, E. plumieri, C. mapale y Ariopsis sp., entre otras (Sánchez et al., 1998; Tíjaro et al., 1998; Rueda y Defeo, 2003; Narváez et al., 2008). Las investigaciones realizadas para el área, han documentado que la mayoría de especies están siendo capturadas en tallas muy pequeñas, por debajo de sus tallas medias de madurez sexual (TMM) (Santos-Martínez et al., 1998; Rueda y Defeo, 2003; Narváez et al., 2008). En el caso de M. incilis, durante el período 1994-2004 se capturó en un amplio ámbito de tallas (5. 5 a 49 cm), observándose incremento en las capturas de individuos juveniles y una mayor presión sobre tallas por debajo de la TMM (Narváez et al , 2008) Es evidente el efecto sobre su biología reproductiva, que probablemente también es afectada por cambios en las condiciones ambientales de la CGSM, debido a las alteraciones de su régimen hidrológico (Botero y Salzwedel, 1999; Blanco et al., 2006). Debido a lo anterior, la renovación natural de M. incilis posiblemente no está ocurriendo de tal manera que le permita enfrentar sosteniblemente la actividad pesquera En este trabajo se investigó el estado actual de M. incilis en la CGSM, determinando algunos aspectos reproductivos y pesqueros con el propósito de confrontar la aseveración anterior. Se consideró importante generar información actualizada sobre esta especie para contribuir a la adopción de medidas de manejo para su pesquería, siendo una de las especies más importantes (30 % de la captura total anual) para la seguridad alimentaria y la generación de ingresos de las comunidades de pescadores de la CGSM (Zamora et al., 2005).
ÁREA DE ESTUDIO
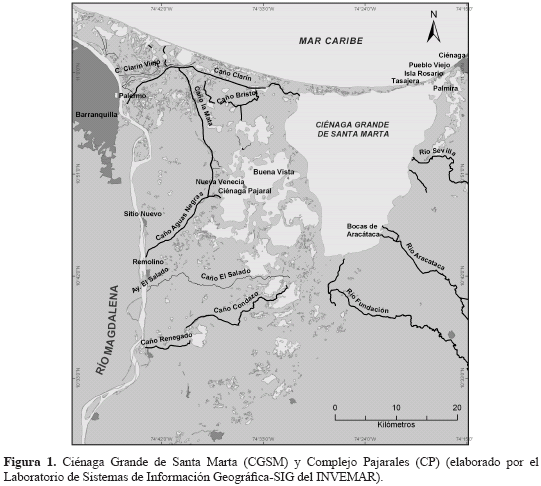
La ecorregión CGSM se encuentra localizada en la costa del departamento del Magdalena, sector delta nororiental del río Magdalena (10° 43' - 11° 00' N y 74° 16' - 74° 43' W; IGAC, 1993; Figura 1). La CGSM ocupa un área de 3812 Km2 aproximadamente, de los cuales 757 Km2 son cuerpos de agua con una profundidad promedio de 1.5 m (Mancera y Botero, 1993; INVEMAR, 2007). El complejo lagunar CGSM está rodeado de extensos bosques de manglar dominados por Rhizophora mangle y Avicennia germinans (Wiedemann, 1973). Las tasas de precipitación anual varían entre 401 y 1321 mm (Blanco et al, 2006). Existen dos períodos climáticos, un período seco que va desde diciembre hasta abril, y otro lluvioso, que va desde abril hasta noviembre, con una pequeña disminución relativa de lluvias entre julio y agosto (Kaufmann y Hevert, 1973; Blanco et al., 2006). La temperatura del agua es alrededor de 30 °C en todo el año (Wiedemann, 1973; Narváez et al., 2008) y el rango de salinidad media está entre 2.3 y 25, variando según las condiciones hidrológicas del lugar (Blanco et al., 2006). Las comunidades de pescadores que viven alrededor del complejo lagunar pueden clasificarse, según su localización, en pueblos costeros, como Tasajera, Palmira, islas del Rosario y Pueblo Viejo, y asentamientos palafíticos, como Bocas de Aracataca, Buenavista y Nueva Venecia. La población aledaña (Ciénaga, Pueblo Viejo, Sitio Nuevo y Remolino) es de aproximadamente 160390 habitantes (DANE, 2005). Se estima que cerca del 80 % de la población económicamente activa extrae artesanalmente y comercializa los recursos pesqueros, tanto para autoconsumo como para el abastecimiento de los centros urbanos periféricos (Ciénaga, Santa Marta y Barranquilla, principalmente) (Zamora, 2005). Una amplia descripción de la CGSM a través del tiempo la presentan Wiedemann (1973), Botero y Mancera (1996) y Botero y Salzwedel (1999).
MATERIALES Y MÉTODOS
Para determinar aspectos de la biología reproductiva de M. incilis, se utilizaron datos provenientes de 12 meses de muestreo efectuados desde diciembre de 2006 hasta noviembre de 2007. En estos muestreos mensuales se obtuvieron ejemplares de M. incilis de las capturas comerciales realizadas por los pescadores del complejo lagunar usando atarrayas, redes agalleras y boliches. Los peces fueron medidos y pesados, conservando muestras en hielo para análisis posteriores. Los datos pesqueros fueron evaluados a partir de información recolectada y compilada en los períodos 1994-1996 y 2000-2007 de los monitoreos pesqueros realizados en la CGSM (Santos-Martínez et al., 1998; INVEMAR, 2007). Los datos fueron colectados semanalmente por encuestadores en los cuatro principales sitios de desembarco en la CGSM, durante cada año monitoreado. Estos datos se almacenaron en el Sistema de Información Pesquera del INVEMAR-SIPEIN (Narváez et al., 2005), con el fin de procesar información Paralelamente a la colecta de las muestras, se registró mensualmente la salinidad y temperatura del agua en 15 estaciones en la CGSM.
Análisis de la información Características reproductivas
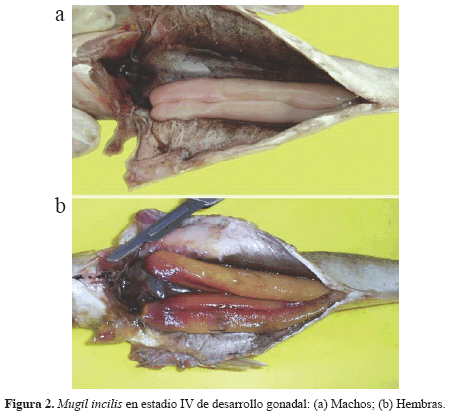
Los ejemplares traídos de campo fueron analizados en laboratorio para determinar el sexo. También fueron medidas su longitud total (LT) y pesados sin visceras con una balanza analítica de 0.001 g de precisión. Se determinaron los estadios de madurez sexual a partir de observación macroscópica del material gonadal, usando la escala de Holden y Raitt (1975) modificada por Santos-Martínez (1989). Esta escala consta de cinco estados (ver Figura 2 para estadio IV): indiferenciados (I); inmaduros (II); madurando (III); maduros (IV); y desovados (V).
Se determinó el factor de condición (K) a partir de los datos de tallas y de pesos de la totalidad de los individuos muestreados (1954), usando la ecuación K = W/aLTb; donde, W es el peso eviscerado de cada individuo en gramos, LT es la longitud total y las constantes a y b son el intercepto y la pendiente de la relación talla- peso (Anderson y Gutreuter, 1983) calculada para las lisas con sexos combinados. Este factor se relacionó con la salinidad del área de estudio. Las diferencias del factor de condición entre muestreos se evaluaron aplicando el análisis de varianza no paramétrico de Kruskal-Wallis. Para conocer cuáles muestreos presentaron los valores significativamente más altos y más bajos del factor de condición, así como la comparación del factor entre sexos, se efectuó la prueba de comparación múltiple de Mann-Whitney. Luego, se determinó la proporción de sexos por meses, utilizando todos los datos obtenidos a partir de la diferenciación de sexos La diferencia en la proporción de sexos se evaluó por medio del estadístico Chi-Cuadrado (χ2). Así mismo, se identificó el pico reproductivo total de M. incilis a partir de las frecuencias de los estadios de madurez sexual con respecto a los muestreos y con el índice gonadosomático (IGS). Este último, se determinó utilizando la relación entre el peso de las gónadas y el peso corporal eviscerado, para así minimizar el error por la variación del peso del contenido estomacal (Anderson y Gutreuter, 1983). Por último, la información de los ejemplares maduros y desovados se utilizó para estimar la talla media de madurez sexual (TMM) de la población Esta estimación relaciona la proporción de individuos maduros con la LT, empleando la ecuación de la curva logística: S(LT)' = 1/[1 + exp(a-b * Lt)]; donde, S(LT)' es la frecuencia acumulada relativa suavizada, y a y b son constantes (Sparre y Venema, 1995).
Aspectos pesqueros y sus asociaciones con las características reproductivas
Las variables pesqueras tomadas del SIPEIN y utilizadas para los análisis fueron la talla media de captura (TMC), la captura total desembarcada (C), el esfuerzo (f) y la captura por unidad de esfuerzo o índice de abundancia relativo (CPUE). Estas estimaciones se hicieron mensualmente, discriminadas por las unidades económicas de pesca (UEPs) que utilizaron los artes atarraya, boliche y trasmallo. Estos artes fueron considerados por ser los de mayor captura para M. incilis (Santos-Martínez et al., 1998; INVEMAR, 2007). Las estimaciones mensuales de TMC se usaron para evaluar las diferencias interanuales aplicando un análisis de varianza no paramétrico de Kruskal-Wallis para cada factor (años, sitios y artes de pesca). Se realizó una comparación entre la TMC y la TMM para evaluar el impacto de la pesca sobre este recurso, donde la segunda fue definida como un punto de referencia límite biológico o tamaño de individuo que no es deseable igualar y menos capturarlos por debajo de éste (Caddy y Mahon, 1995; Rueda y Defeo, 2003).
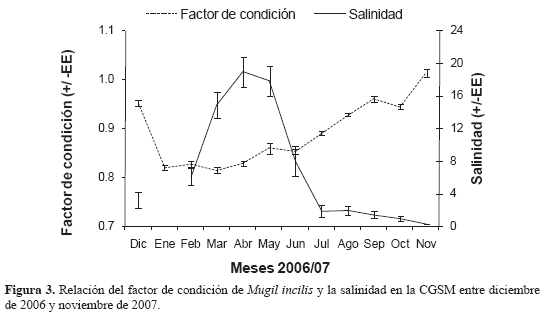
Para determinar tendencias estacionales, se calculó el promedio de todos los años (1994-2007), correspondiente a cada uno de los meses (por ejemplo enero de todos los años) de la CPUE y de la TMC con los principales artes de pesca (atarraya, boliche y trasmallo) y se compararon gráficamente con los registros del IGS y TMM. Mientras que los datos de la temperatura promedio del agua no mostraron variaciones anuales significativas, la salinidad aparentemente es la variable que mejor describe la variabilidad ambiental del sistema lagunar y muestra cambios mensuales Por ello se exploró una relación entre el cambio mensual de la salinidad media y el factor K promedio calculado, como indicadores de la calidad ambiental y la condición fisiológica de la lisa, para observar si ha habido cambios en dicha relación desde los resultados de Blanco (1983), cuando utilizó el factor K de Fulton (1902).
RESULTADOS
Características reproductivas
En los doce meses de muestreo (diciembre de 2006-noviembre de 2007) se obtuvo una muestra total de 1954 individuos, de los cuales 1238 fueron hembras con un intervalo de talla de 17.4-29.9 cm de LT, 482 machos (17.7 - 28.5 cm) y 234 indiferenciados (15-23 cm). La tendencia de los valores del factor K obtenidos mensualmente y su variabilidad se observan en la Figura 3. La relación entre la condición con la salinidad media mostró correlación inversa significativa (r de Spearman = -0.76; p<0.05), siendo el 59 % de la variación de K explicado por la salinidad Los valores del peso de las lisas mostraron la tendencia a incrementarse durante octubre a noviembre, en plena época reproductiva y cuando la salinidad en la CGSM es baja. Por el contrario, el factor de condición descendió entre enero a agosto, en períodos posteriores al desove. De esta manera se observó que el factor de condición presenta diferencias entre muestreos (Kruskal-Wallis: H' = 779.649; n = 1702; p<0. 05), observándose la mejor condición en noviembre y la menor en marzo de 2007. También se observó variación entre sexos (prueba de Mann-Whitney: W = 263755; n = 1702; p<0.05), siendo las hembras las de mejor condición (Figura 3). A su vez, la relación talla-peso calculada para las lisas mostró un crecimiento alométrico relativo en peso, al determinarse un coeficiente de alometría (b) menor a 3 en la ecuación potencial común (Sparre, 1995).
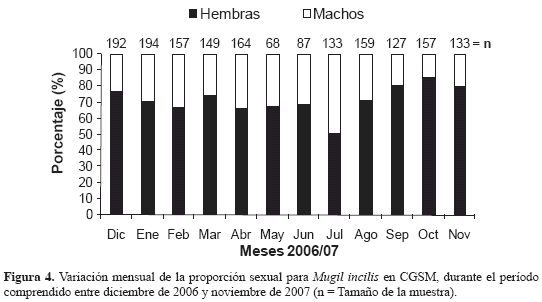
La proporción de machos a hembras observada en M. incilis fue de 1:2.6, representando el 28 y 72 %, respectivamente. Destaca un aumento en la presencia de más hembras que machos en casi todos los meses, excepto en julio cuando la proporción fue de 1:1 (Figura 4) Con respecto a la época de desove, la variación temporal de los estadios de madurez señaló la predominancia de ejemplares inmaduros durante la mayoría de los meses de muestreo, excepto en diciembre 2006 y noviembre 2007, período en el que se observó el porcentaje más alto de individuos maduros para ambos sexos (Figura 5) y los máximos valores del IGS promedio (Figura 6).
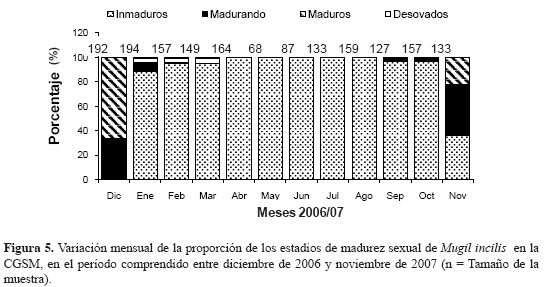
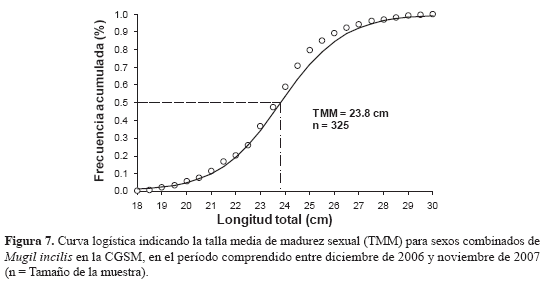
Para determinar la TMM se obtuvieron 325 animales en estadios III, IV y V; de ellos 250 fueron hembras y 75 machos. La TMM para sexos combinados fue de 23.8 cm LT (Figura 7), mientras que la de las hembras (24 cm) fue mayor que la de los machos (23.1 cm). La menor talla en la cual se observó que los machos de M. incilis inician su maduración sexual (TPM) fue de 17.7 cm de LT, mientras que para las hembras fue de 18.6 cm.
Aspectos pesqueros y sus asociaciones con las características reproductivas
Se determinó que las unidades económicas de pesca (UEPs) de atarraya son las que más capturan a M. incilis, fluctuando entre 374 y 1098 t y un promedio de 750 (±213) t por año. Le siguen en captura el boliche (165-729 t) con un promedio de 361 (±212) t por año y los trasmallos (30-555 t) con 232 (±178) t por año (Figura 8).
En todo el período de estudio se determinó una TMC de 24.4 (±1. 2) cm, la cual varió entre años (Kruskal-Wallis: H' = 37.08; p<0.05), registrándose el valor más alto en 2004 y el más bajo en 2007. En lo referente a las artes de pesca, la TMC también varió (Kruskal-Wallis: H' = 197.25; n = 712; p<0.05) y las UEP de atarraya capturaron los peces más pequeños (60 %), mientras los trasmallos capturaron los más grandes (Mann-Whitney: W = 57893; n = 565; p<0. 05), con el 40 % de los individuos muestreados (Figura 9). Con relación a los sitios, también fue evidente la variación espacial (Kruskal-Wallis: H' = 61.05; n = 712; p<0.05). En isla del Rosario y Tasajera se capturaron los peces más pequeños y en Nueva Venecia los de mayor talla (Mann-Whitney: W = 46206; n = 528; p<0.05) En la mayoría de los casos, M. incilis fue capturada por debajo de la TMM En forma consistente, los valores más altos del IGS se observaron al final y principio de año, coincidiendo con los mayores valores de CPUE y TMC por tipo de UEP (Figura 6).
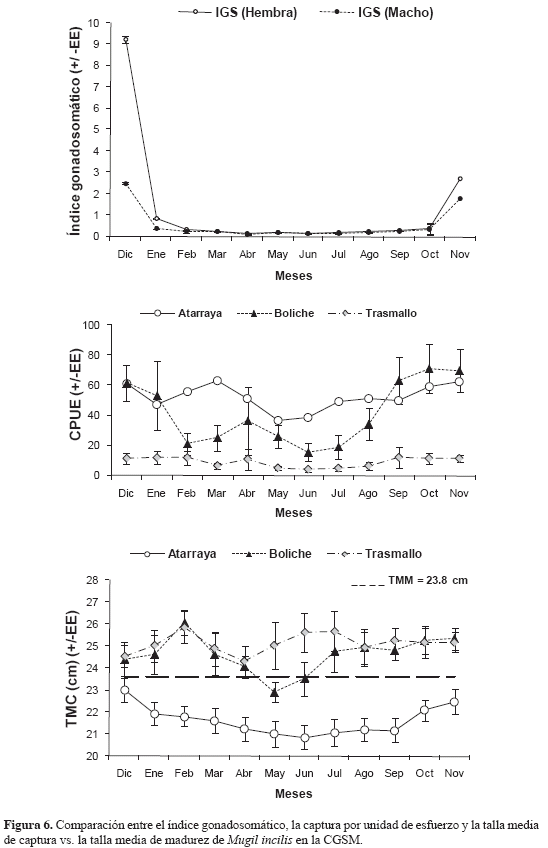
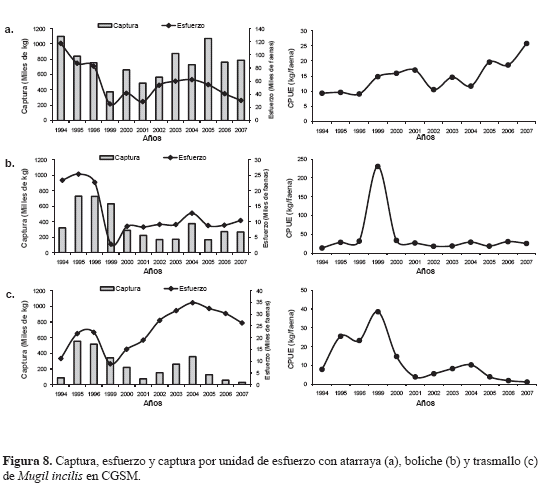
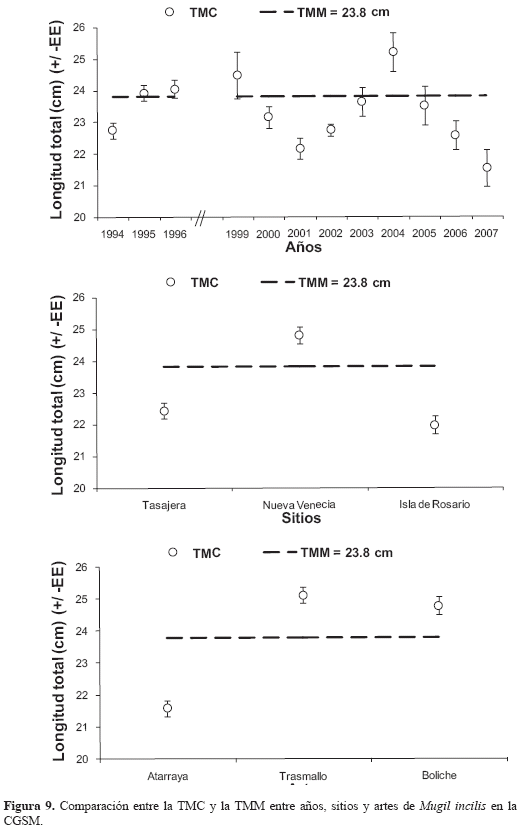
DISCUSIÓN
Este trabajo analiza algunas características reproductivas de M. incilis para la CGSM. En cuanto a la determinación de la proporción de sexos, ésta fue de 1: 2.6 (machos por hembra respectivamente) a lo largo del periodo muestreado. Aunque Nikolsky (1963) señala que la proporción de sexos puede variar considerablemente dentro de la misma especie en diferentes cuerpos de agua, menciona que esta proporción tiende a ser de 1:1. Margalef (1977) señaló que la relación entre el número de machos y hembras está influida por mecanismos genéticos y de regulación ambiental. Autores como Thompson (1966), refiriéndose a las lisas, mencionan que en el desove es usual ver que varios machos asedian a cada hembra, lo cual es poco probable en este caso y eventualmente pudiera representar un riesgo para esta población, al influir sobre el éxito de la fecundación.
La presencia de hembras maduras y desovadas, durante diciembre de 2006, y en enero y febrero de 2007, indica que esta especie se reproduce sólo una vez al año, siendo consistente con otros estudios que argumentan que M. incilis se reproduce a final del año (Perdomo, 1973; Blanco, 1980; Velasco y Villegas, 1985; Sánchez et al., 1998). Si se atiende a los valores del índice gonadosomático de la población, puede inferirse que la etapa final de maduración suele ocurrir en diciembre, aunque se observó un aumento del índice a partir de septiembre, cuando se supone que los peces se han recuperado físicamente de su pasada migración de desove al mar y aprovechan la energía extra para madurar sus gónadas (Nikolsky, 1963).
Por ser una proporción del peso y la talla, el factor de condición depende de la escasez o abundancia del alimento y de la madurez sexual del pez (Albertini-Berhaut, 1975), asociadas probablemente a la condición ambiental, en este caso indicada por la variación de la salinidad media en la CGSM Se reafirma lo mencionado por Blanco (1983) respecto a que la condición física de las lisas varía inversamente con la salinidad Tratándose de una especie eurihalina, es poco probable que el cambio de salinidad le afecte Sin embargo, asociado al cambio de salinidad ocurre un cambio en las condiciones tróficas, demostrado por Osorio (1985) para la lisa de la CGSM, cuando dice que a finales del año y con el régimen de bajas salinidades, se produce un aumento en la oferta de alimento bentónico, como son los foraminíferos, en el sedimento aportado por los ríos tributarios y que les sirven como alimento a M. incilis (Blanco, 1983). Lo anterior puede determinar un incremento en el factor de condición y del desarrollo gonadal en los meses de diciembre y enero cuando la salinidad baja gracias al agua dulce que entra al sistema desde sus tributarios crecidos (Blanco et al , 2006). La coincidencia de los valores relativamente altos del IGS indica que la especie comienza la migración de su ciclo reproductivo para esa época Alvarez-Lajonchere (1976) observó este mismo comportamiento con Mugil curema en aguas cubanas.
Comparando los valores de la TMM estimada en este trabajo (23 8 cm de LT) con los documentados por Perdomo (1973) (34 cm) y Sánchez et al. (1998) (25 7 cm), se observa una disminución en las estimaciones debido a la presión pesquera ejercida en los últimos años, principalmente sobre el tamaño de los peces, dado que la mayoría son capturados muy jóvenes (Narváez et al., 2008). Así, la abundancia de peces de tallas pequeñas y una menor frecuencia de capturas de los individuos de mayor tamaño en la muestra, hace disminuir el promedio de tallas de las lisas maduras Parece ser una explicación razonable a la disminución de la TMM observada. Ricker (1979) menciona un efecto similar causado cuando el mayor esfuerzo se ejerce sobre el estrato más joven de la población. Aunque Nikolsky (1963) menciona una aceleración de la maduración de los individuos por contar con mayor disponibilidad de alimento al disminuir su población, no indica que esto necesariamente ocurra en los individuos más jóvenes No se puede atribuir dicho decrecimiento en TMM a alguna estrategia fisiológica de la especie, por carecer al momento de evidencia científica que permita corroborar la hipótesis de un efecto compensatorio ante la disminución de la población producto de la presión pesquera y de los cambios ambientales en la CGSM.
Los resultados de la relación talla-peso concuerdan con los de Blanco (1980) para la misma especie en la CGSM, y no se encontraron diferencias significativas entre machos y hembras En este estudio se evidencia que las UEP atarrayeras son las que más afectan negativamente la población de M. incilis, debido a que capturan los individuos más pequeños y la mayor parte de ellos por debajo de la TMM Teniendo en cuenta que el 14 % de las capturas están incluso por debajo de la talla de primera madurez (TPM) y que el 56 % se encuentra por debajo de la TMM, se comprueba que M. incilis está siendo sobreexplotada En este contexto debe tenerse en cuenta que, en general, las artes de pesca en la CGSM (atarrayas, trasmallos, boliches), han mostrado una disminución gradual en los tamaños de malla usados de 2000 a 2004 (Narváez, et al., 2008), lo cual facilitaría la captura de ejemplares cada vez de menores tallas, aún por debajo de la TMM, con el consecuente impacto sobre las poblaciones de recursos pesqueros en general
Aunque indudablemente existen otras variables pesqueras que pueden explicar el estado de un recurso, su estudio rebasa los alcances de este trabajo, cuyo aporte se circunscribe al análisis del esfuerzo, la variabilidad en las capturas y al probable impacto de un cambio en la talla de reclutamiento que influyan en la sostenibilidad del recurso, afectando las probabilidades de reproducción de la población Este enfoque se dirigió a comparar aspectos como el IGS con K y la CPUE, así como la TMC y la TMM por arte de pesca para explicar el tipo de uso actual del recurso M. incilis en la CGSM. En este caso se demostró que cuando la especie inicia y finaliza su ciclo reproductivo es cuando más se captura porque entonces es más apetecida comercialmente por su alta condición, su contenido de ovarios maduros y por su vulnerabilidad debido a su comportamiento migratorio y de cardúmenes. En esta situación es cuando debe ejercerse mayor control y vigilancia en la explotación de este recurso.
AGRADECIMIENTOS
Fue financiado por el INVEMAR a través del Banco de Proyectos de Inversión de la Nación (BPIN), en el marco del proyecto "Monitoreo de las condiciones ambientales y los cambios estructurales y funcionales de las comunidades vegetales y de los recursos pesqueros durante la rehabilitación de la Ciénaga Grande de Santa Marta". Agradecemos al INVEMAR por su apoyo, al grupo de trabajo de la línea Uso y Producción Sostenible del Programa Valoración y Aprovechamiento de Recursos Marinos y Costeros del INVEMAR, coordinado por el Dr. Mario Rueda, y al M. Sc. Juan Carlos Narváez Barandica por sus valiosos comentarios.
BIBLIOGRAFÍA
1 Albertini-Berhaut, J. 1975. Biologie des stades juvéniles de Teleosteens Mugilidae Mugil auratus Risso 1810, Mugil capito Civier 1829 et Mugil saliens Risso 1810. Partie III: Croissance lineaire et ponderale de Mugil capito dans le Golfe de Marseille. Acuacultura, 5: 179-197. [ Links ]
2 Álvarez-Lajonchere, L. 1976. Contribución al estudio del ciclo de vida de Mugil curema Valenciennes in Cuvier et Valenciennes, 1836 (Pisces: Mugilidae). Invest. Mar., 28: 3-130. [ Links ]
3 Anderson, R. O. y S. J. Gutreuter. 1983. Length, weight, and associated structural indices. 283-300. En: Nielsen L.A. y D. L. Johnson (Eds.). Fisheries techniques. American Fisheries Society, Bethesda, EE.UU. [ Links ]
4 Blanco, J. 1980. Algunos aspectos ecológicos y biológico-pesqueros de la lisa, Mugil incilis Hancock 1830 en la Ciénaga Grande de Santa Marta "Colombia" Pisces Mugilidae. Tesis Biol., Univ. Nacional Colombia, Bogotá. 69 p. [ Links ]
5 Blanco, J. 1983. The condition factor of Mugil incilis Hancock (Pisces: Mugilidae) and its seasonal changes in the Ciénaga Grande de Santa Marta (Colombia). An. Inst. Invest. Mar. Punta Betín, 13: 133-142. [ Links ]
6 Blanco, J. A., E. A. Viloria y J. C. Narváez B. 2006. ENSO and salinity changes in the Ciénaga Grande de Santa Marta coastal lagoon system, Colombian Caribbean. Estuar. Coast. Shelf Sci., 66: 157-167. [ Links ]
7 Botero, L. y J. Mancera. 1996. Síntesis de los cambios de origen antrópico ocurridos en los últimos 40 años en la Ciénaga Grande de Santa Marta (Colombia). Rev. Acad. Colomb. Cienc., 20 (78): 465-474. [ Links ]
8 Botero, L. y H. Salzwedel. 1999. Rehabilitation of the Ciénaga Grande de Santa Marta, a mangroveestuarine system in the Caribbean coast of Colombia. Ocean Coast. Manag., 42: 243-256. [ Links ]
9 Caddy, J. F. y R. Mahon. 1995. Reference points for fishery management. FAO Fish. Techn. Paper, 347. 83 p. [ Links ]
10 DANE. 2005. Resultados censo general 2005: Total de hogares, viviendas y personas. Departamento Administrativo Nacional de Estadísticas. Colombia. http://www.dane.gov.co/censo/files/consulta/2005_compensada.xls. [ Links ]
11 Fulton, T. 1902. Rate of growth of sea fishes. Sci. Invest., Fish Div. Scot. Report, 20: 20 p. [ Links ]
12 Holden, M. y D. Raitt. 1975. Manual de ciencias pesqueras. Parte II: Métodos para la investigar los recursos pesqueros y su aplicación. Doc. Técn. Pesca FAO, 115: 1-211. [ Links ]
13 IGAC. 1993. Monografía del departamento del Magdalena. Instituto Geográfico Agustín Codazzi, Bogotá. 147 p. [ Links ]
14 INVEMAR. 2007. Monitoreo de las condiciones ambientales y los cambios estructurales y funcionales de las comunidades vegetales y de los recursos durante la rehabilitación de la Ciénaga Grande de Santa Marta. Informe final, Santa Marta. 105 p. [ Links ]
15 Kaufmann, R. y H. Hevert. 1973. El régimen fluviométrico del río Magdalena y su importancia para la Cienaga Grande de Santa Marta. Mitt. Inst. Col. Alemán Invest. Cient., 7: 121-137. [ Links ]
16 Mancera, J. E. y L. Botero. 1993. Estudio ecológico de la Ciénaga Grande de Santa Marta - Delta exterior del río Magdalena- Etapa III. Primer informe de avance presentado a COLCIENCIAS. Programa lagunas costeras. Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta. 128 p. [ Links ]
17 Margalef, R. 1977. Ecología. Omega S. A., Barcelona. 600 p. [ Links ]
18 Narváez, J., M. Rueda, E. Viloria, J. Blanco, J. Romero y F. Newmark. 2005. Manual del sistema de información pesquera del INVEMAR (SIPEIN Ver.3.0): Una herramienta para el diseño de sistemas de manejo. Serie de documentos generales No. 18, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta. 128 p. [ Links ]
19 Narváez, J., F. Herrera y J. Blanco. 2008. Efecto de los artes de pesca sobre el tamaño de los peces en una pesquería artesanal del Caribe colombiano. Bol. Invest. Mar. Cost., 37 (2): 163-187. [ Links ]
20 Nikolsky, G. 1963. The ecology of fishes. Academic Press, Nueva York. 312 p. [ Links ]
21 Osorio, D. 1985. Ecología trófica de Mugil curema Valenciennes, 1836, Mugil incilis Hancock, 1830 y Mugil liza Valenciennes, 1836 (Pisces: Mugilidae) en la Ciénaga Grande de Santa Marta, Caribe colombiano. Tesis Biol., Univ. Nacional Colombia, Bogotá. 65 p. [ Links ]
22 Perdomo, J. M. 1973. Contribución al estudio biológico-pesquero de la lisa Mugil incilis Hancock. Tesis Lic. Cien. Mar, Fundación Univ. Bogotá Jorge Tadeo Lozano, Bogotá. 93 p. [ Links ]
23 Ricker, W. 1979. Methods for assessment of fish production in fresh water. International Biological Programme. Handbook N° 3, Blackwell Scientific Publications, Oxford. 348 p. [ Links ]
24 Rueda, M. y O. Defeo. 2003. Spatial structure of fish assemblages in a tropical estuarine lagoon: combining multivariate and geostatistical techniques. J. Exp. Mar. Biol. Ecol., 296: 93-112. [ Links ]
25 Sánchez, C., M. Rueda y A. Santos. 1998. Dinámica poblacional y pesquería de la lisa, Mugil incilis en la Ciénaga Grande de Santa Marta, Caribe colombiano. Rev. Acad. Colomb. Cienc., 22 (85): 507-517. [ Links ]
26 Santos-Martínez, A. 1989. Estudio biológico y ecológico de la ictiofauna de la Ciénaga Grande de Santa Marta, Caribe colombiano. Tesis M.Sc., Univ. Nacional Colombia, Bogotá. 178 p. [ Links ]
27 Santos-Martínez, A. y A. Acero P. 1991. Fish community of the Ciénaga Grande de Santa Marta (Colombia). Composition and zoogeography. Ichthyol. Explor. Freshwaters, 2 (3): 247-263. [ Links ]
28 Santos-Martínez, A., E. Viloria, C. Sánchez, M. Rueda, R. Tíjaro, M. Grijalba y J. C. Narváez. 1998. Evaluación de los principales recursos pesqueros de la Ciénaga Grande de Santa Marta y Complejo Pajarales, Caribe colombiano. Informe final, Colciencias, INVEMAR y GTZ-Prociénaga, Santa Marta. 205 p. [ Links ]
29 Sparre, P. y S. C. Venema. 1995. Introducción a la evaluación de stocks pesqueros tropicales. Parte 1. FAO Doc. Técn. Pesca, 306, 376 p. [ Links ]
30 Thompson, J. M. 1966. The grey mullets. Oceanog. Mar. Biol. Ann. Rev., 4: 301-335. [ Links ]
31 Tíjaro, R., M. Rueda y A. Santos-Martínez. 1998. Dinámica poblacional del chivo mapalé Cathorops spixi en la Ciénaga Grande de Santa Marta y Complejo de Pajarales, Caribe colombiano. Bol. Invest. Mar. Cost., 27: 87-102. [ Links ]
32 Velasco, A. y P. Villegas. 1985. Aportes al conocimiento de la historia de la vida de Mugil incilis (Hancock, 1830) Mugil liza (Valenciennes, 1836) y Mugil curema (Valenciennes, 1836) en la Ciénaga de Tesca (Bolívar, Colombia). Tesis Biol. Mar., Fundación Univ. Bogotá Jorge Tadeo Lozano, Bogotá. 55 p. [ Links ]
33 Wiedemann, H. 1973. Reconnaissance of the Ciénaga Grande de Santa Marta, Colombia: physical parameters and geological history. Mitt. Inst. Col. Alemán Invest. Cient., 7: 85-119. [ Links ]
34 Zamora, A. 2005. Evaluación de impacto socioeconómico asociado a cambios ambientales del ecosistema Ciénaga Grande de Santa Marta en el periodo 1999-2003. Tesis Economía Internacional, Univ. Magdalena, Santa Marta. 124 p. [ Links ]
FECHA DE RECEPCIÓN: 16/06/2008 FECHA DE ACEPTACIÓN: 22/04/2010
*Contribución No. 1058 del Instituto de Investigaciones Marinas y Costeras-INVEMAR.
