










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.39 no.2 Santa Marta July/Dec. 2010
DINÁMICA OCÉANO-ATMÓSFERA Y SU INFLUENCIA EN LA BIOMASA FITOPLANCTÓNICA, EN LA ZONA COSTERA DEL DEPARTAMENTO DEL MAGDALENA, CARIBE COLOMBIANO
OCEAN-ATMOSPHERE DYNAMICS AND THEIR INFLUENCE ON THE PHYTOPLANKTON BIOMASS OFF THE MAGDALENA REGION COASTAL ZONE, COLOMBIAN CARIBBEAN
Lorena Marcela García-Hoyos, Andrés Franco-Herrera, Jeimmy Samanta Ramírez-Barón y Diego Alejandro López-Cerón
Universidad de Bogotá Jorge Tadeo Lozano, Facultad de Ciencias Naturales, Programa de Biología Marina, Carrera 2 No. 11-68. El Rodadero, Santa Marta, Magdalena, Colombia. lorenagarcia802@hotmail.com (L.G.H.), andresfranco@utadeo.edu.co (A.F.H.)
RESUMEN
Durante los meses de febrero a junio de 2008, en las aguas costeras del departamento del Magdalena, específicamente en las ensenadas de Gaira y Granate y en la bahía de Taganga, se realizaron seis muestreos que correspondieron a las épocas climáticas seca mayor y lluviosa menor, con el fin de determinar la influencia del acoplamiento océano-atmósfera sobre la intensidad de los eventos de surgencia, considerando la variabilidad fisicoquímica en la columna del agua comprendida entre los 0 y 60 m de profundidad y la respuesta de la comunidad fitoplanctónica medida en términos de clorofila a. Se evaluaron variables atmosféricas, oceanográficas y biológicas siguiendo el método Euleriano. De acuerdo a las condiciones océano-atmosféricas durante el período de estudio, se presentaron eventos de surgencia por transporte de Ekman y acción de vientos continentales durante los meses de febrero y abril, en respuesta a condiciones adecuadas para su desarrollo; velocidad promedio (>4.0 m/s) y dirección del viento (paralela y perpendicular a la costa). Se observaron alteraciones en las condiciones fisicoquímicas de la masa de agua, presentándose bajas temperaturas (25.34 ± 0.82 °C), altas salinidades (35.5 ± 0.30) y subsaturación de oxígeno (70.6 ± 7.8 %). El aporte de nutrientes inorgánicos fue mayor durante eventos de surgencia definiendo al sistema de carácter mesotrófico, lo que favoreció la magnitud de la biomasa fitoplanctónica con mayores concentraciones de hasta 1.60 µg/L. El evento La Niña 2007/2008, que cubrió los períodos en los que se realizó la investigación, no desencadenó ninguna alteración importante sobre las condiciones típicas de eventos de surgencia.
PALABRAS CLAVE: Surgencia, Vientos alisios, Nutrientes, Mesotrófico, Clorofila a.
ABSTRACT
From February to June 2008 at inshore sites of the Magdalena region, specifically in Gaira and Granate inlets and Taganga Bay, six field trips were done comparing the main dry and short rainy seasons The main aim was to determine the coupled ocean-atmosphere influence on the upwelling intensity, considering the physical and chemical variability within the water column between 0 and 60 m and the response of the phytoplankton community measured as chlorophyll a. Atmospheric, oceanographic, and biological conditions were evaluated following the Eulerian method. According to the oceanic and atmospheric conditions during the research period, Ekman transport and continental upwelling processes occurred through February and April, in response to adequate conditions: average wind speed (>4.0 m/s) and wind direction (parallel and perpendicular to the coast). Physical and chemical anomalies were detected in the water mass, such as low temperature (25.34 ± 0.82°C), high salinity (35.5 ± 0.30) and oxygen subsaturation (70.6 ± 7.8 %). The inorganic nutrient supply was higher during upwelling processes, consequently, the system was defined as mesotrophic, causing greater phytoplankton chlorophyll a biomass with levels up to 1.60 µg/L. "La Niña" 2007/2008 event, which covered the period of this research, did not incite any important disturbance over the typical upwelling conditions.
KEY WORDS: Upwelling, Trade winds, Nutrients, Mesotrophic, Chlorophyll a.
INTRODUCCIÓN
En los océanos, los centros de mayor productividad biológica se encuentran localizados a lo largo de los bordes orientales. En estas áreas, variaciones a pequeña escala desencadenan patrones de vientos periódicos y definidos los cuales al incidir sobre la superficie del mar generan corrientes superficiales y subsuperficiales que tienen un efecto directo sobre las propiedades fisicoquímicas de la columna de agua.
Al encontrarse el Caribe colombiano bajo la influencia de los desplazamientos norte-sur de la Zona de Convergencia Intertropical (ZCIT) y del movimiento meridional del sistema de monzones americanos, durante la estación de fuertes vientos (vientos alisios) la ZCIT permanece en el sur, alrededor de los 0-5° de latitud sur, permitiendo que el viento del este ejerza influencia sobre la región y en consecuencia ocurran procesos de surgencia (INVEMAR, 2004). Éstos importan cantidades de nutrientes a las capas superficiales, favoreciendo las comunidades fitoplanctónicas y muy probablemente la producción primaria nueva, estableciendo de esta forma una zona de alta oferta alimenticia para las comunidades planctívoras (Franco-Herrera, 2005).
A lo largo de la costa colombiana, es usual la ocurrencia de procesos de surgencia costera por transporte de Ekman y acción de vientos, continentales debido a la morfología de la costa y al régimen de vientos, donde sus direcciones prevalecen del noreste y sureste respectivamente, con velocidades superiores a 4.0 m/s durante la época seca mayor que comprende el período diciembre-abril (Franco-Herrera, 2005). Otra condición relevante para que se generen dichos procesos, es que la fuerza de los vientos sobre las capas superficiales del mar produzca niveles de estrés a profundidades de Ekman capaces de desplazar masas de agua hacia mar afuera y superar la profundidad de termoclina permanente que según Blanco et al. (1994) se ubica entre 40 y 50 m, lo cual ha sido confirmado para la zona por Fajardo (1979), Márquez (1982), Blanco (1988), Bula-Meyer (1990), Corredor (1992), Cabrera y Donoso (1993) y Arévalo-Martínez y Franco-Herrera (2008).
En contraparte, durante las estaciones húmedas, la surgencia es prácticamente nula debido a la ausencia de los vientos alisios (Ramírez, 1983; Bula-Meyer, 1990; Donoso, 1990). No obstante, otras vías de fertilización marina conocidas como corrientes salobres de mar adentro (ricas en elementos nutritivos), las cuales proceden de ríos como el Magdalena, el Sinú y el Atrato y de lagunas costeras como la Ciénaga Grande de Santa Marta producen una proliferación de microalgas planctónicas (Ramírez, 1983; Blanco, 1988; Bula-Meyer, 1990). La importancia de las dos vías de fertilización de las aguas marinas costeras (i.e. aportes continentales y surgencia) del departamento del Magdalena es que éstas mantienen altas concentraciones de fitoplancton a través del año desencadenando un constante proceso de flujo de materia y energía a lo largo de cada eslabón subsiguiente de la red trófica marina, que contribuye al desarrollo del ecosistema pelágico (Franco-Herrera, 2005).
Durante el período de estudio, la costa de Colombia estuvo bajo el dominio del evento océano-atmosférico La Niña 2007/2008. Eventos como éstos alteran el clima en diferentes regiones del planeta (Sánchez et al., 2001; Trenberth, 2003) y en consecuencia modifican la temperatura del aire, la humedad y el viento. De manera que el resultado de las variaciones en el calor latente y los flujos de agua dulce podrían inducir cambios en la temperatura superficial del mar (TSM), salinidad, capa de mezcla y corrientes oceánicas (Alexander et al., 2002).
Bajo la perspectiva anterior, el propósito de esta investigación fue evaluar la dinámica océano-atmósfera y su efecto sobre la variabilidad fisicoquímica de la columna de agua (0-60 m) en la zona costera del Magdalena durante las épocas seca mayor y lluviosa menor y determinar su influencia en la oferta de nutrientes inorgánicos y las concentraciones de clorofila a. Al mismo tiempo, se tuvo en cuenta el evento La Niña 2007/2008, evaluando su efecto sobre las condiciones típicas de eventos de surgencia, ampliando en espacio y tiempo las investigaciones realizadas por Arévalo-Martínez y Franco-Herrera (2008), quienes proponen que el sistema tropical del Caribe centro colombiano es de características mesotróficas y de alta oferta de alimento autotrófico.
ÁREA DE ESTUDIO
La región costera de Santa Marta comprende desde la desembocadura del río Magdalena hasta la desembocadura del río Piedras en el límite oriental del Parque Nacional Natural Tayrona. Sus rasgos morfológicos y paisajísticos están determinados por el macizo montañoso de la Sierra Nevada de Santa Marta, cuyas estribaciones septentrionales se adentran hacia el mar alternándose con bahías y playas arenosas en su interior y cabos rocosos con acantilados (Figura 1). La plataforma continental, es prácticamente ausente en esta área y los fondos se precipitan rápidamente a profundidades de más de 500 m a distancias relativamente cortas de la línea de costa (INVEMAR et al., 2004). Los períodos climáticos en el área están regidos por los patrones generales que influyen la costa atlántica colombiana. El desplazamiento norte-sur de la ZCIT define la época seca y lluviosa, siendo la primera de diciembre a abril y la segunda de mayo a noviembre. Cuando la ZCIT se desplaza hacia el sur, la región se encuentra bajo la influencia de los vientos alisios del noreste que son reforzados por diferentes eventos locales (Franco-Herrera, 2005); como consecuencia, se genera una corriente en sentido paralelo a la costa en la parte oriental del área y hacia mar afuera en la parte occidental, produciendo un evento local de surgencia (Bula-Meyer, 1985; Ramírez, 1990). Cuando el desplazamiento se da hacia el norte, se produce una disminución en la velocidad de los vientos alisios, además de favorecer las precipitaciones de la región (Franco-Herrera, 2005), y debido a la ausencia de surgencia las aguas superficiales tienden a ser más cálidas (Bula-Meyer, 1985; Ramírez, 1990).

MATERIALES Y MÉTODOS
Diseño experimental
Se efectuaron seis muestreos durante los meses de febrero a junio de 2008 en las bocas de las ensenadas de Gaira y Granate y en la bahía de Taganga, específicamente en estaciones bajo las coordenadas 11°12'44"N y 74°14'44"W, 11°16'57"N y 74°12'40''W y 11°15'54''N y 74°12'40"W, respectivamente siguiendo el método Euleriano, en el cual se realizan determinaciones de todas las variables a medir en campo en un solo punto por bahía o ensenada de muestreo (Cognetti y Magazzú, 2001; Fernández, 2003). Se tomaron muestras de agua mediante el uso de la botella Vand Dorn de 6 L de capacidad a 0, 5, 30 y 60 m de profundidad, teniendo en cuenta dos criterios básicos: a) que la profundidad de la capa fótica en esta área alcanza máximo los 25 m de profundidad (Franco-Herrera, 2005), por lo cual, en los primeros estratos de la columna de agua (0-5 m) se da la mayor actividad autotrófica y en consecuencia la dinámica de nutrientes y b) dada la importancia de la surgencia en el área, que incluye masas de agua por debajo de la termoclina permanente, se tuvo como fin tener muestras de agua por encima y una por debajo de la misma, basados en el ámbito de profundidad de esta discontinuidad térmica, que según Blanco et al. (1994), varía entre 40 y 50 m. En estas profundidades se realizaron las mediciones de las variables en estudio in situ (i.e. temperatura, salinidad, oxígeno disuelto y pH) mediante el uso de sondas multiparámetros WTW-330 (con un mínimo de detección > 0.1) y se recolectaron muestras de nutrientes (i.e. nitratos, nitritos, amonio, ortofosfatos) y clorofila a, siendo almacenadas en recipientes de polipropileno de 2 L de capacidad nominal, los cuales fueron previamente oscurecidos y refrigerados para evitar su degradación hasta su procesamiento en el laboratorio. Se tuvieron en cuenta las metodologías propuestas por Garay et al. (2003) para la cuantificación de las concentraciones de nutrientes inorgánicos y de Strickland y Parsons (1972), para determinar la concentración de pigmentos fitoplanctónicos, siguiendo el método espectrofotómetrico y las ecuaciones propuestas por Jeffrey y Humphey (Salamanca, 1996).
Con el fin de establecer el comportamiento de las variables atmosféricas, se determinó la dirección y velocidad del viento diaria a las 08:00, 14:30 y 17:30, una semana previa a cada muestreo a partir de la información obtenida de la estación meteorológica Davis-Wizard III, ubicada a 10 msnm, en la sede de Santa Marta de la Universidad de Bogotá Jorge Tadeo Lozano, y se consideraron las imágenes satelitales del Cooperative Institute for Meteorological Satellite Studies de la University of Wisconsin-Madison (CIMSS, 2008) para describir los campos de vientos y como consecuencia las condiciones atmosféricas imperantes de la zona durante el período en estudio. De acuerdo con la base de datos atmosférica, se evaluó el estrés del viento sobre la superficie del agua para la ensenada de Gaira y se consideró el mismo valor para la ensenada de Granate y bahía de Taganga ya que la extensión de la costa desde Gaira hasta Granate ≈ 11.30 km) no es significativa, y todas las estaciones de muestreo se posicionaron en las bocas de las ensenadas y bahías, lo que permite inferir que los campos de vientos actuan de manera homogénea en dichas estaciones.
A partir del cálculo del estrés del viento sobre la superficie del mar, se siguió el modelo matemático de Ekman para establecer la profundidad de Ekman (Pond y Pickard, 1983). Se tomaron imágenes satelitales en un intervalo de cuatro días durante todo el período de muestreo para establecer el inicio de la termoclina permanente y el patrón general de corrientes para el mar Caribe a partir del servicio SARPAR del Centro de Investigaciones Oceanográficas e Hidrográficas de la Armada Nacional de Colombia (CIOH, 2008).
Se construyeron perfiles de profundidad para cada variable y se observó su estructura en la columna del agua. Finalmente, se determinó la ocurrencia del evento El Niño-La Niña mediante el índice ONI (Oceanic Niño Index) a partir de la base de datos del Climate Prediction Center de la NOAA. Se tuvieron en cuenta valores de TSM correspondientes a las masas de agua de la ensenada de Gaira, bahía de Taganga y ensenada de Granate mediante mediciones in situ e imágenes satelitales de las mismas estaciones procesadas por el Institute for Marine Remote Sensing de la University of South Florida (USF, 2008) en el intervalo diciembre 2006-abril 2008. Así a partir de promedios mensuales y trimestrales se estableció la tendencia de esta variable junto con la velocidad del viento en dicha escala temporal y se determinó la relación entre ellas y el índice ONI mediante figuras de análisis comparativo. A continuación se hicieron pruebas de ámbitos de correlación Spearman (Zar, 1999) entre las variables velocidad del viento y TSM con el índice ONI para establecer los niveles de significación de la posible asociación entre ellas, teniendo como marco de referencia que la presencia del evento de surgencia en la zona costera del departamento del Magdalena presenta una isoterma superficial de 25 °C (Arévalo-Martínez y Franco-Herrera, 2008).
Análisis de las variables fisicoquímicas y biomasa fitoplanctónica
Las variables físicas y químicas medidas en la columna de agua comprendida entre 0 y 60 m de profundidad se analizaron con ayuda de estadística descriptiva cuantificando medias, máximos, mínimos y desviación estándar. Para determinar si existían diferencias estadísticamente significativas entre pares de estaciones para cada una de las variables medidas, se realizó un análisis pareado-dependiente efectuando pruebas de Wilcoxon (Zar, 1999); así mismo, se agrupó la información de cada una de las variables en dos niveles de contrastes (i.e. surgencia y no surgencia) y se repitió la prueba de Wilcoxon con el fin de establecer cambios significativos entre las dos condiciones oceanográficas. Adicionalmente para valorar si la profundidad tenía efecto sobre el comportamiento de las variables se empleó una prueba de Friedman (Zar, 1999) mediante el programa estadístico STATISTICA versión 8.0. Con el fin de mostrar las tendencias en el comportamiento de dichas variables en la columna del agua se realizaron perfiles de profundidad para los períodos correspondientes a eventos de surgencia y no surgencia. Se determinaron los valores mínimos, máximos y promedios de los nutrientes inorgánicos considerados, para establecer una base de comparación con trabajos relacionados (i.e. Ramírez, 1983; Arévalo-Martínez y Franco-Herrera, 2008) a razón de comprobar el grado de fertilización de las aguas costeras del departamento del Magdalena por el transporte de nutrientes cuantificado durante eventos de surgencia y no surgencia. Posteriormente, se procedió con un análisis de componentes principales (ACP) donde se tuvieron en cuenta las variables fisicoquímicas y la biomasa fitoplanctónica con el objetivo de dilucidar la incidencia de dichas variables en la masa de agua en los dos eventos en mención mediante el programa estadístico PRIMER® v.5. Finalmente, con el propósito de determinar la asociación entre las variables ambientales (variables atmosféricas y oceanográficas) y el componente biótico (biomasa fitoplanctónica) se realizaron correlaciones de Pearson con un 95 % de confianza, estandarizando las variables a correlacionar, volviéndolas adimensionales y en la misma escala (Zar, 1999) a partir del programa STATGRAPHICS® v.5.1.
RESULTADOS
Condiciones atmosféricas y oceanográficas ante eventos de surgencia y no surgencia
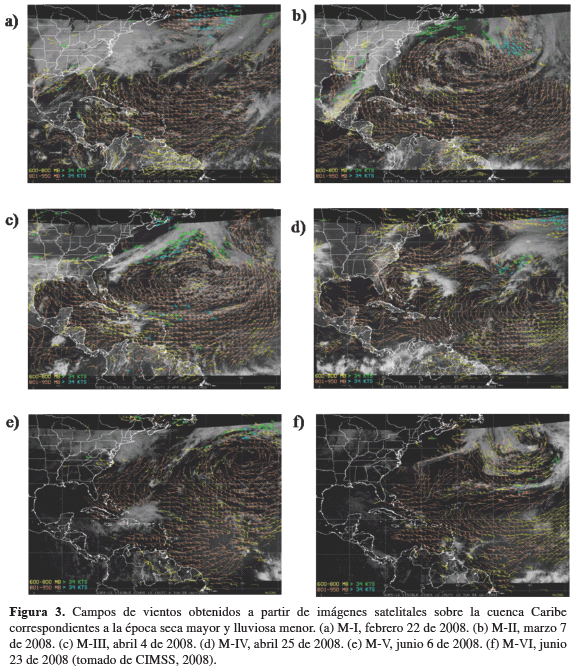
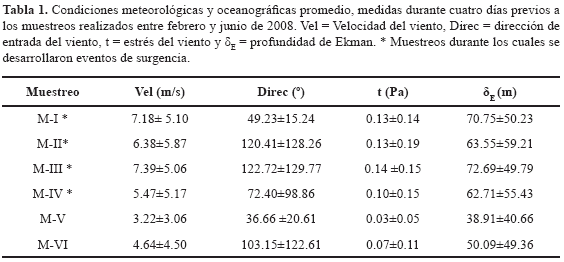
Velocidad y dirección del viento. En el primer semestre del año 2008, cuando se presentan las épocas climáticas seca mayor (diciembre a abril) y lluviosa menor (mayo a junio), la velocidad promedio del viento fue 3.7 ± 3.4 m/s y 2.1 ± 2.5 m/s, respectivamente, con ráfagas que oscilaron entre 4.5 y 21.0 m/s en las horas de la tarde y la madrugada, en respuesta a las mayores intensidades de los vientos alisios sobre la ensenada de Gaira. Los campos de vientos presentaron una dirección de entrada promedio de 70 ± 64 lo que implica una dirección predominante NE-SO (Figura 2). De igual manera, durante los muestreos efectuados, tanto la velocidad promedio (>4.0 m/s) como la dirección del viento (paralela y perpendicular a la costa) indicaron condiciones favorables para la ocurrencia de eventos de surgencia por transporte de Ekman y acción de vientos continentales para los muestreos (M-I) a (M-IV), los cuales se realizaron durante la primera época del año, a diferencia de los muestreos (M-V) y (M-VI) (muestreos de no surgencia) desarrollados en la segunda época, regidos por velocidades promedio más bajas (Tabla 1). Entre tanto, en las estaciones de Taganga y Granate, se presentaron vientos predominantes del noreste con velocidades que oscilaron entre 2.6 y 15.4 m/s para los primeros cuatro muestreos. Hacia el quinto y sexto, la incidencia de los vientos alisios sobre la región costera del Magdalena fue mínima (Figura 3).
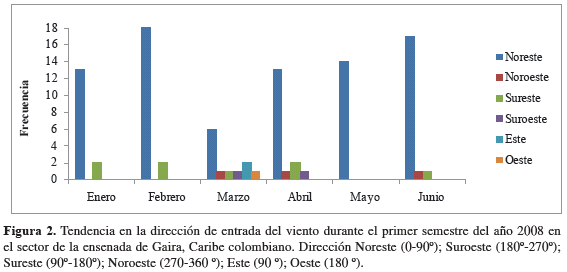
Stress del viento y profundidad de Ekman. Durante los días previos a los muestreos correspondientes a períodos de surgencia, la velocidad del viento generó niveles de estrés sobre la superficie del mar que oscilaron entre < 0.01 y 0.58 Pa, transmitiéndose su efecto hasta alcanzar una profundidad promedio de Ekman de hasta 72.70 ± 49.80 m (Tabla 1). Para los muestreos correspondientes a períodos de no surgencia, la disminución de la velocidad promedio del viento dio lugar a un estrés del viento fluctuante entre < 0.01 y 0.22 Pa, condición insuficiente para la generación de una profundidad de Ekman adecuada (i.e. mayor que la profundidad de la termoclina permanente) que promoviera la ocurrencia de eventos de surgencia (38.91 ± 40.66 m). Sin embargo, durante el último muestreo, en respuesta a la intensidad del viento, los niveles de estrés del viento oscilaron entre 0.003 y 0.372 Pa dando lugar a una profundidad promedio de Ekman de 50.09 ± 49.36 m, pero con dirección desfavorable para generar desplazamientos de agua mar adentro. Aún así, de acuerdo con las condiciones océano-atmosféricas, se consideraron en consecuencia a los muestreos (M-I) a (M- IV) correspondientes a condiciones de eventos de surgencia. Durante los muestreos uno y cuatro (M-I y M-IV) se presentó surgencia por transporte de Ekman a diferencia de los muestreos dos y tres (M-II y M-III) donde se presentó surgencia por acción de vientos continentales.
Corrientes superficiales. La corriente superficial del mar para el litoral norte y centro del Caribe colombiano durante los primeros cuatro muestreos correspondientes a la época climática seca mayor, estuvo dominada por la corriente costera del Caribe, la cual lleva aguas superficiales en dirección suroeste, presentando velocidades que fluctuaron entre 0.1 y 0.3 m/s. Para el quinto y sexto muestreo (M-V y M-VI), correspondientes a la época climática lluviosa menor, se evidenció la presencia de la corriente del Darién de dirección oeste-este con intensidades hasta de 0.7 m/s.
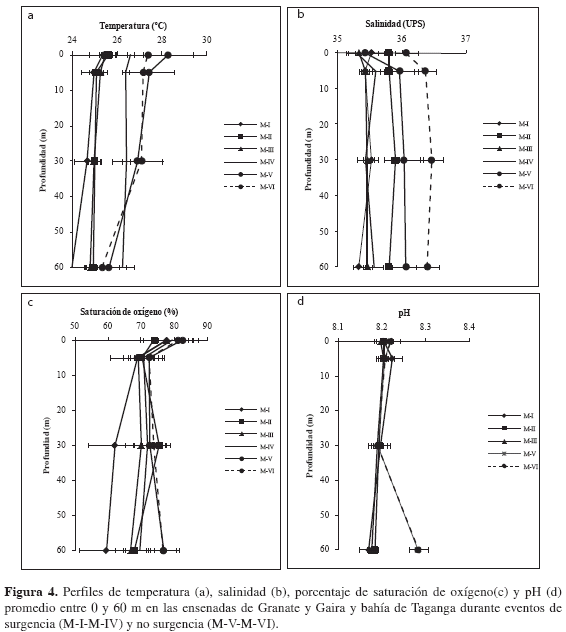
Temperatura, salinidad y estabilidad del agua. La temperatura promedio durante el período de muestreo en la columna de agua entre 0-60 m fue 25.86 ± 1.28 °C. No hubo diferencias significativamente estadísticas entre estaciones (Wilcoxon, p > 0.05). Aunque tampoco existieron diferencias entre las temperaturas del agua correspondientes a muestreos bajo condiciones de surgencia y de no surgencia (Wilcoxon, p >0.05). Los valores más altos se registraron para los muestreos quinto y sexto (M-V y M-VI) correspondientes a períodos de no surgencia y los más bajos para los muestreos correspondientes a períodos de surgencia (M-I, M-II, M-III y M-IV; Tabla 2 y Figura 4a). Se evidenció que las profundidades tuvieron efecto sobre dicha variable (Friedman, p < 0.05), presentando un gradiente de disminución en la temperatura a medida que se aumentó en profundidad.
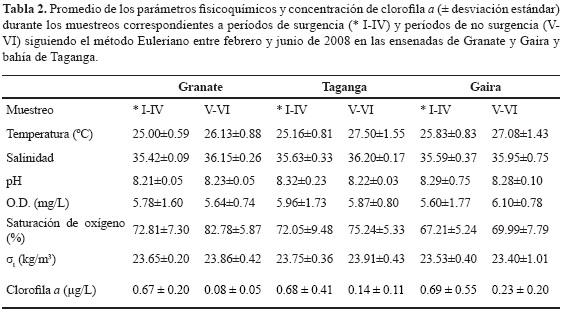
El comportamiento de la variable salinidad osciló entre 34.2 y 36.6 (promedio 35.7 ± 0.44). Los valores más altos se presentaron en períodos de no surgencia y los más bajos en períodos de surgencia (Tabla 2 y Figura 4b). No se encontraron diferencias estadísticamente significativas entre las dos épocas ni entre estaciones (Wilcoxon, p > 0.05). En cuanto a su distribución vertical la salinidad tuvo una tendencia leve a ser mayor en las capas superficiales y menor en las subsuperficiales a lo largo de los muestreos correspondiente a eventos de surgencia, de manera que se evidenció el efecto de la profundidad sobre esta variable (Friedman, p < 0.05).
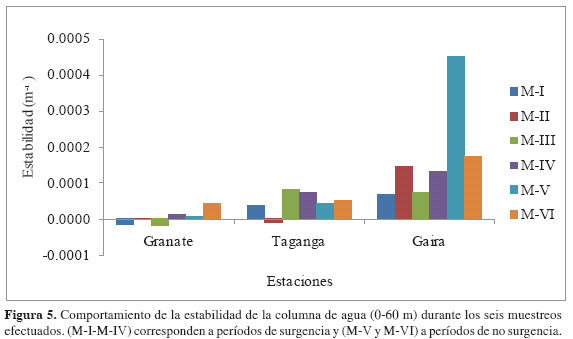
Los valores promedios de densidad del agua fueron relativamente homogéneos en la columna del agua y a través del tiempo (Tabla 2), sin embargo las diferentes profundidades tuvieron un efecto sobre esta variable (Friedman, p < 0.05), presentándose mayores valores en las capas superficiales hasta el muestreo IV y enseguida una notable homogenización en toda la columna del agua. No existieron diferencias estadísticamente representativas entre estaciones y tampoco entre épocas (Wilcoxon, p > 0.05). Como consecuencia, la estabilidad de la columna de agua se mantuvo en su mayoría positiva (Figura 5). Sin embargo, para los muestreos M-I, M-II y M-III se obtuvieron valores negativos, específicamente en Granate (-1.49 x 10-5m-1 y -1.82 x 10-5m-1) y Taganga (-6.77 x 10-6m-1), respectivamente.
Oxígeno disuelto, porcentaje de saturación de oxígeno y pH. El oxígeno disuelto osciló entre 3.8 y 9.14 mg/L (promedio 5.82 ± 1.43 mg/L). El valor máximo se presentó a 0 m durante el muestreo M-IV en Granate (9.14 mg/L) y el mínimo (3.82 mg/L) a 60 m durante el muestreo M-I en Taganga, lo que pone en evidencia el efecto de la profundidad sobre esta variable (Friedman, p < 0.05). En general, se evidenciaron aguas superficiales con un contendido bajo de oxígeno disuelto en el muestreo M-I, M-II y M-III A partir del muestreo M-IV, existió un aumento en la concentración que disminuyó hacia el muestreo M-VI, sin embargo el ámbito de variación no fue suficiente para generar diferencias estadísticamente representativas entre períodos de surgencia y no surgencia y tampoco entre estaciones (Wilcoxon, p > 0.05). El comportamiento de la variable porcentaje de saturación de oxígeno fue análogo a lo obtenido para el oxígeno disuelto, lo que se traduce en la predominancia de aguas subsaturadas a lo largo de la bahía y ensenadas muestreadas (Figura 4c). El pH, en contraste, se mantuvo alto durante todos los muestreos (promedio 8.26 ± 0.15; Tabla 2 y Figura 4d), indicando un contenido bajo de CO2, de tal forma que no existieron diferencias estadísticamente significativas entre estaciones y entre períodos de surgencia y no surgencia (Wilcoxon, p > 0.05) Las profundidades tuvieron efecto sobre el comportamiento de esta variable (Friedman, p < 0.05), presentándose en general mayores valores en las capas superficiales (0-50 m) y un leve decremento en las capas subsuperficiales (30-60 m).
Nutrientes. La concentración de NO3= osciló entre 1.92 y 30.00 µg-at-N- NO3=/L. En general, se presentó un gradiente en disminución de la concentración media de este nutriente a nivel espacial de norte a sur (Granate-Taganga-Gaira; Figura 6a). No existieron diferencias significativamente estadísticas entre períodos correspondientes a surgencia y no surgencia (Wilcoxon, p > 0.05), ni entre estaciones (Wilcoxon, p > 0.05). Sin embargo, es importante resaltar que las concentraciones medias más altas se dieron para los muestreos M-III y M-VI, que pueden representar la intensidad máxima del período de surgencia y el aporte inicial de aguas continentales durante el período correspondiente a la época seca y lluviosa menor, respectivamente (Figuras 6a y 7) Las diferentes profundidades tuvieron efecto sobre esta variable (Friedman, p < 0.05), presentándose en general un patrón irregular en la distribución.
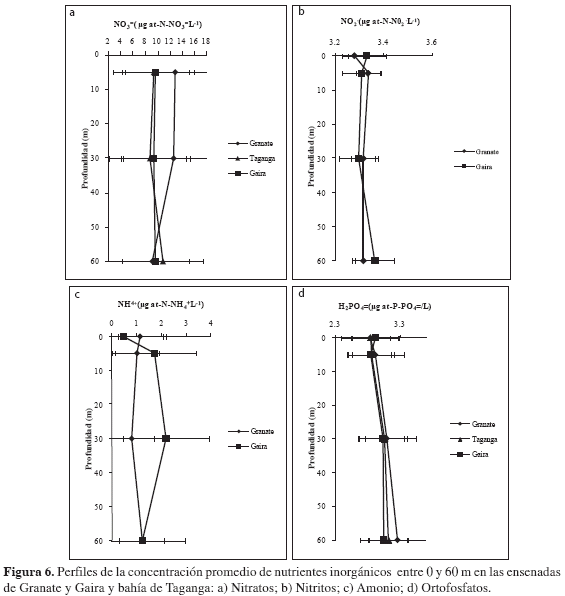
Por su parte, las concentraciones de NO2- fueron similares entre estaciones, oscilando entre 3.24 y 3.62 µg-at-N-NO2=/L. No existieron diferencias estadísticamente significativas entre estaciones (Wilcoxon, p > 0.05; Figura 6b) y tampoco se presentaron diferencias estadísticamente significativas entre periodos de surgencia y no surgencia (Wilcoxon, p > 0.05; Figura 7).
Las concentraciones medias de NH4+, oscilaron entre 0.04 y 5.88 µg-at-N-NH4+/L. No hubo diferencias estadísticas entre periodos de surgencia y no surgencia y tampoco entre estaciones (Wilcoxon, p > 0.05; Figura 6c y 7).
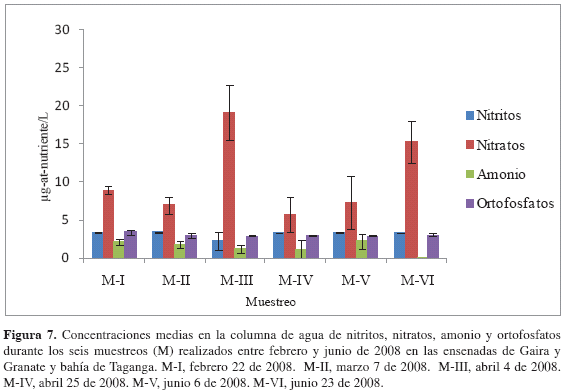
Finalmente, se encontró que las concentraciones promedio de ortofosfatos oscilaron entre 2.59 y 4.68 µg-at-P-PO4=/L. No existieron diferencias representativas entre estaciones (Wilcoxon, p > 0.05; Figura 6d). Se observó que las concentraciones fueron mayores en períodos de surgencia y menores en períodos de no surgencia (Figura 7), sin embargo, no existieron diferencias estadísticamente significativas entre dichos períodos (Wilcoxon, p > 0.05). Para los últimos tres nutrientes considerados (i.e. NO2=, NH4+ y H2PO4=), se encontró que la profundidad tuvo efecto sobre su comportamiento (Friedman p < 0.05), generando una tendencia a encontrar mayores concentraciones en las capas superficiales. Esto indicaría que corresponde a columnas de agua con comportamientos similares en el gradiente de profundidad, sin importar si hay la presencia de un evento de surgencia o ausencia de ella.
Biomasa fitoplanctónica. La biomasa fitoplanctónica osciló entre <0.01 y 1.610 µg/L durante el período febrero-junio de 2008. Los valores más altos se presentaron para los muestreos correspondientes a períodos de surgencia (I-IV) y los más bajos para aquellos de no surgencia (V-VI) (Figura 8a), sin embargo, no existieron diferencias estadísticamente significativas (Wilcoxon, p > 0.05; Tabla 2). Ya que las concentraciones fueron similares entre estaciones, no existieron diferencias estadísticamente significativas (Wilcoxon, p > 0.05; Figura 8b). Se observó mayor concentración del pigmento en las capas superficiales y subsuperficiales durante los tres primeros muestreos (MI, MII y MIII) y en los siguientes no se presentó un patrón claro de segregación vertical.
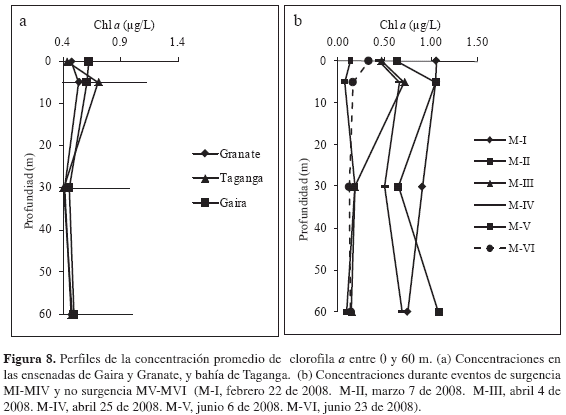
Ninguna de las variables estudiadas presentó diferencias contrastantes o estadísticamente significativas, lo que supone una masa de agua homogénea durante el primer semestre del año, inherente a la inestabilidad atmosférica que impidió la marcada estacionalidad de la época seca mayor y lluviosa menor en la zona costera del departamento del Magdalena. Sin embargo, las condiciones químicas de la columna de agua y la oferta de nutrientes inorgánicos en eventos de surgencia fueron indicadores primarios de condiciones favorables para el desarrollo de mayores concentraciones de clorofila a. En contraste, para el período en ausencia de eventos de surgencia las condiciones promovieron menores biomasas fitoplanctónicas.
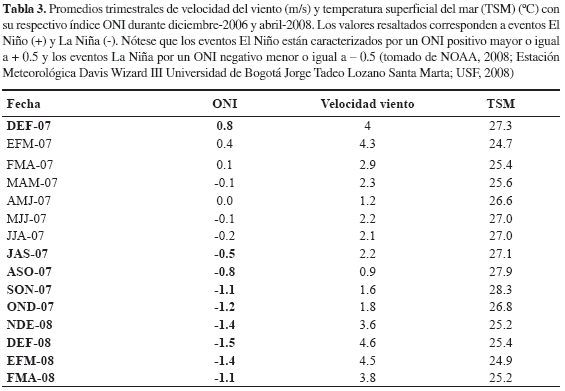
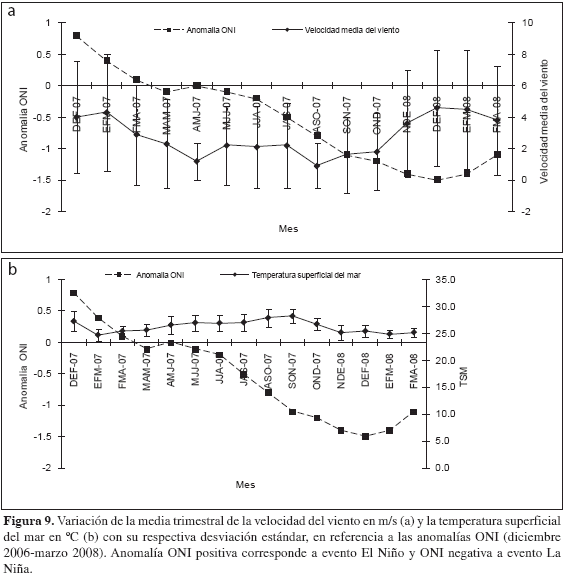
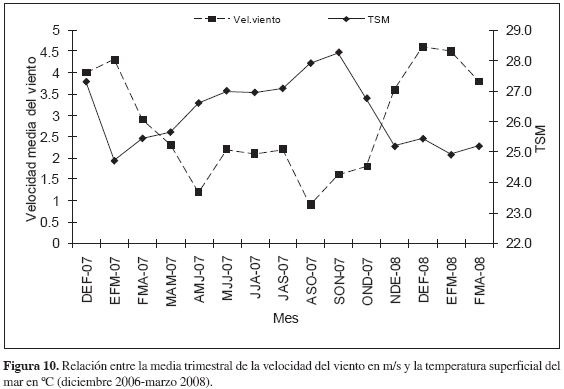
Efecto del evento atmosférico La Niña 2007/2008 sobre los eventos de surgencia. Los valores de TSM obtenidos a partir de imágenes satelitales provenientes de la base de datos de la University of South Florida (USF, 2008) no presentaron diferencias estadísticamente significativas con aquellos tomados in situ (t-pareada, p > 0.05) por lo cual, para establecer si los eventos El Niño-La Niña tuvieron algún impacto en intensificar o debilitar los eventos de surgencia en la región del Magdalena, se trabajó a partir de la imagen satelital. En el período interanual 2006-2008, se presentó un evento El Niño durante los meses de diciembre-2006 a febrero-2007 de acuerdo al índice ONI (0.8) y ocurrió un evento La Niña de larga duración en los meses de julio-2007 a abril-2008 con un índice ONI fluctuante entre -0.5 y -1.5 (Tabla 3) La variable velocidad del viento considerada de alta importancia ante el efecto de dichos eventos océano-atmosféricos, superó para el año 2008 en su mayoría los valores de velocidad promedio en comparación con el año 2007, es decir que durante eventos El Niño la velocidad del viento en la zona tendió a disminuir y durante eventos La Niña aumentó, sin embargo no presentó correlación significativa con el índice ONI (Spearman, r= -0.4. p > 0.05. Figura 9a). De igual forma, tampoco existió correlación significativa entre el índice ONI y TSM (Spearman, r= 0.64. p > 0.05; Figura 9b). Por otro lado, la TSM estuvo significativamente correlacionada con la velocidad del viento (Spearman, r= -0.68. p < 0.05; Figura 10). Lo anterior indica que el evento La Niña 2007/2008 no desencadenó ninguna alteración importante sobre las condiciones típicas de eventos de surgencia durante el período de estudio.
Dinámica biótica y abiótica (análisis de componentes principales). Se eligieron tres componentes que acumularon el 66 % de la varianza total de los datos, sin embargo, se contemplaron los dos primeros componentes que reunieron la mayor variabilidad con respecto al total (50.3 %). En el primer componente, que explicó el 31.9 % de la variación, la clorofila a y salinidad fueron variables que se correlacionaron negativamente, lo que en términos ecológicos se traduce en una mayor biomasa fitoplanctónica durante períodos de salinidad menores o viceversa. De manera similar, para el componente 2 que explicó el 18.4 % de la variación obtenida, el pH, la temperatura y NO3= fueron las variables que caracterizaron este factor (Figura 11a).
Así mismo, para el análisis de los grupos de muestreos, se observó una débil segregación de los muestreos realizados bajo condiciones de surgencia y no surgencia (Figura 11b). Para el primer componente, se evidenció la segregación de los muestreos pertenecientes a los diferentes periodos estudiados. Es así como los muestreos de surgencia (MI-MIV) presentaron una tendencia definida a agruparse entre sí mismos, sin la existencia de una discriminación entre profundidades al igual que los muestreos correspondientes a períodos de no surgencia (MV-MVI). Cabe resaltar que el muestreo M-IV, fue sobresaliente sobre el componente II y de esta forma se consideró como un muestreo de transición que respondió a la variabilidad fisicoquímica propia de la finalización del periodo de surgencia y comienzo del periodo de lluvias.
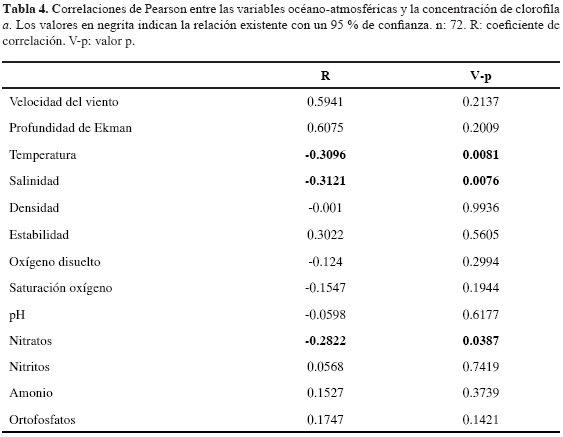
Correlaciones entre la biomasa fitoplanctónica y variables abióticas. Se presentaron asociaciones significativas entre la biomasa fitoplanctónica y las variables ambientales salinidad, temperatura y nitratos (Tabla 4). Las correlaciones inversas más alta y estadísticamente significativa se obtuvieron entre clorofila a y temperatura, seguida por clorofila a y nitratos.
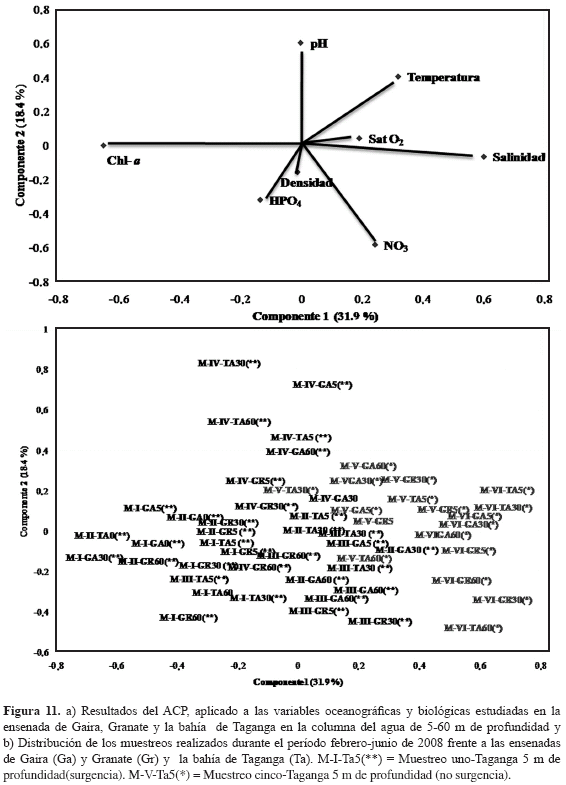
DISCUSIÓN
Los resultados encontrados se presentan en concordancia con trabajos previos que describen la dinámica océano-atmosférica en el área del Caribe colombiano (Fajardo, 1979; Márquez, 1982; Blanco, 1988; Bula-Meyer, 1990; Corredor, 1992; Cabrera y Donoso, 1993; Arévalo-Martínez y Franco-Herrera, 2008). Estos autores corroboran la ocurrencia de eventos de surgencia por transporte de Ekman (dirección del vector viento: 26°) y acción de vientos continentales (dirección del vector viento: 120°) durante los meses de febrero-abril correspondientes a la época de fuertes vientos (vientos alisios), donde el posicionamiento de la ZCIT se encuentra alrededor de los 0-5° S, permitiendo que el viento del este ejerza influencia sobre la región y de esta manera ocurran procesos de surgencia (INVEMAR, 2004). Como producto de la fuerza de estos vientos sobre las capas superficiales del mar, se obtuvieron niveles de estrés del viento y profundidades de Ekman capaces de desplazar masas de agua hacia mar afuera y superar la profundidad de la termoclina permanente que, según Blanco et al. (1994), se ubica entre 40 y 50 m.
Cabe resaltar, sin embargo, que las condiciones atmosféricas imperantes fueron afectadas también por sucesos de escala regional (e.g. frentes fríos, ondas tropicales, inestabilidad en el posicionamiento de la ZCIT) que representaron alteraciones en la tendencia general del comportamiento del viento (variando su dirección e intensidad) e indudablemente repercutieron sobre la dinámica oceanográfica siendo los frentes fríos muy frecuentes durante el período de investigación, los cuales actuaron como canal para que el viento se intensificara en dirección este-noreste, con velocidades que oscilaron entre 8 y 13 m/s y ráfagas de 18 m/s. De igual manera, ocasionaron un aumento significativo en la altura de la ola y de acuerdo a la permanencia de su actividad desencadenaron descensos en la TSM. Adicionalmente a la variabilidad atmosférica, se sumó el tránsito de la primera onda tropical del año que causó perturbaciones en los sistemas de presión atmosférica generando el desarrollo de núcleos conectivos incitadores de lluvia hacia la primera semana del mes de abril (CIOH, 2008), en la cual se efectuó el muestreo (M-IV), donde evidentemente en respuesta, se observó un comportamiento diferencial en la variabilidad fisicoquímica de la columna del agua y a su vez señaló la finalización de la época seca mayor.
El posicionamiento de la ZCIT influyó en la variabilidad estacional de las características en el campo de la presión atmosférica, ejerciendo un importante control en los campos de viento y precipitación. De manera que la ubicación de los sistemas de alta presión de las Azores y baja presión del Darién modularon los campos isobáricos y gradientes de presión actuando como los factores responsables del comportamiento del viento, así que cuando estuvo activo el primer sistema, el flujo sinóptico de los vientos siempre aumentó, a diferencia de la activación del segundo sistema, que conllevó a la disminución en la velocidad del viento y aumento de la cobertura nubosa. Lo anteriormente mencionado permitió inferir que la dinámica atmosférica del período febrero-junio de 2008, donde sobrevienen las épocas climáticas seca mayor y lluviosa menor, se presentó inestable con una tendencia de la estación seca mayor a ser perdurable durante todo el primer semestre del año en respuesta a los fenómenos atmosféricos ocasionales y no a acontecimientos de gran escala como el evento La Niña 2007/2008, que cubrió la durabilidad del presente estudio
El evento La Niña se presentó entre los meses de julio de 2007 y junio de 2008 (Figura 9a), partiendo del supuesto de que en el Atlántico norte tropical y en el mar Caribe, las anomalías positivas en la TSM durante períodos La Niña, se evidencian aproximadamente de tres a seis meses después de los picos en las anomalías de la TSM en el Pacífico tropical, según lo planteado por Covey y Hastenrath (1978) era de esperarse que para la costa Caribe colombiana algunas condiciones físicas de la columna del agua fueran afectadas (e.g. TSM); sin embargo, esto no ocurrió. Si bien existió un incremento en la velocidad del viento durante los meses de enero a abril de 2008 en comparación con el mismo período del año 2007, lo que repercute directamente sobre la TSM, el evento ENSO no explicó estas diferencias (Figura 9b), confirmando lo dilucidado anteriormente, es decir, que las variaciones climáticas regionales pueden ser una manifestación no necesariamente de los patrones de circulación atmosférica de larga escala, sino de los fenómenos océano-atmosféricos locales ocasionales (Bell y Chelliah, 2005).
De otro lado, el período de surgencia estuvo dominado por la corriente del Caribe de dirección noreste-suroeste, generando un giro ciclónico o divergente que tiende a arrastrar agua por debajo de la termoclina permanente hacia la superficie, lo que resulta en un aporte de nutrientes que contribuye a la productividad de la zona (Andrade, 2001). En contraparte, para el período mayo-junio, correspondiente a la época lluviosa menor, se evidenció la presencia de la contracorriente del Darién de dirección suroeste-noreste, que lleva aguas cálidas superficiales, en respuesta al patrón de vientos dominante del suroeste Bajo este contexto, las dos corrientes tienen influencia sobre las masas de agua costeras del departamento del Magdalena y regulan parcialmente los patrones de circulación local dentro de las ensenadas y bahías (Franco-Herrera, 2005).
Teniendo en cuenta que las masas de agua costeras del Magdalena (i.e. Granate, Taganga y Gaira) están influenciadas por el patrón climático general que rige la costa atlántica colombiana, durante el período febrero-abril, dichas aguas presentaron un comportamiento definido en respuesta a eventos de surgencia. La distribución vertical de los parámetros oceanográficos se caracterizó por el ascenso de aguas, las cuales exhiben valores bajos de temperatura (21-24 °C) y subsaturación de oxígeno (Müller, 1979; Ramírez, 1983, 1987; Bula-Meyer, 1990), permitiendo evidenciar el influjo de la masa de agua subtropical sumergida que se localiza entre los 100 y 200 m de profundidad en el mar Caribe (Bula-Meyer, 1985). En contraparte a los ámbitos de salinidad durante eventos de surgencia propuesto por los mismos autores (36.5-37.2), en esta investigación los valores altos correspondieron a períodos en ausencia de surgencia debido a la alta evaporación, el retraso y baja intensidad de las lluvias, y en consecuencia a las débiles descargas de origen continental y escorrentía (CIOH, 2008; IDEAM, 2008).
Las temperaturas medias de las aguas costeras en mención oscilaron entre 23.4 y 27.3 °C, presentándose los niveles más bajos para los tres primeros meses del año y de manera contraria la salinidad presentó valores altos que oscilaron entre 35 y 36.5 debido a la entrada marcada de la época seca, el incremento en la intensidad del viento y, en efecto, el evento de surgencia (Franco-Herrera, 2005).
Consecuentemente, las concentraciones de oxígeno disuelto en el agua, las cuales son dependientes de factores como la temperatura, salinidad (Pond y Pickard, 1983) y la acción del viento tendieron a producir valores promedio de subsaturación de 82.87 ± 25.04 %. Autores como Müller (1979), Ramírez (1983), Franco-Herrera (2001), Tigreros (2001), Vanegas (2002) y Arévalo-Martínez y Franco-Herrera (2008) confirman la variación estacional en la zona de estas propiedades oceanográficas, conforme períodos de surgencia y no surgencia. Para la época lluviosa, el aumento de la nubosidad, la disminución del viento y la llegada de las precipitaciones generaron un incremento en la temperatura del agua y la tendencia de las aguas a la saturación o sobresaturación de oxígeno disuelto (Franco-Herrera, 2005), encontrándose valores de temperatura durante la ausencia de períodos de surgencia, entre 24.9 y 30 °C y saturación de oxígeno en un ámbito de 67.05 y 132.85 %.
La evidente estratificación halina de las masas de agua del mar Caribe, genera como consecuencia un gradiente de densidad, cuyo principal resultado es la alta estabilidad de la columna de agua (Corredor, 1992). No obstante, como se mencionó anteriormente, existió la presencia de un proceso de desplazamiento vertical de agua durante los meses de febrero a abril que permitió el proceso de surgencia, lo cual se ajusta a lo postulado por Ramírez (1983), quien declara que la estabilidad disminuye cuando hay ascenso de aguas de fondo densas y aumenta cuando existe un gradiente creciente de la densidad con la profundidad. Los resultados de Ramírez (1983) en la bahía de Santa Marta, corroboran el comportamiento de la estabilidad del agua para el presente estudio, al encontrar mayores disminuciones entre diciembre y abril, asociadas a los valores más altos de la densidad superficial del agua, tipificando así la temporada de surgencias más intensa del año, y condiciones inversas, es decir, valores altos de la estabilidad asociados a la disminución de la densidad del agua para el resto del año (Ramírez, 1983; Salzweder y Müller, 1983).
Por otro lado, los resultados obtenidos para el pH, fueron homogéneos en la columna del agua y se mantuvieron estables a lo largo del primer semestre del año Aunque no existió un patrón de segregación vertical definido, según Millero (2006) el pH es mayor en las capas superficiales debido a la fotosíntesis. La pérdida de CO2 incrementa el pH, éste a su vez disminuye por la oxidación de la materia orgánica y va en decremento hasta 1 Km de profundidad.
En cuanto a los nutrientes, cuya concentración está condicionada por la acción de eventos físicos, químicos, geológicos y biológicos en las masas de agua costeras del departamento del Magdalena, no es evidente una fuerte fluctuación estacional como si lo es para otras variables fisicoquímicas (Franco-Herrera, 2005), sin embargo la fertilización de nutrientes por eventos de surgencia define el sistema como mesotrófico. Las concentraciones medias de nitratos y ortofosfatos se presentaron relativamente altas, a diferencia del comportamiento de los nitritos y amonio que mostraron concentraciones bajas durante todo el estudio. Ramírez (1990) registra una concentración máxima de nitratos en la surgencia de Santa Marta de 4 µg-at-N-NO3=/L a principios de febrero, siendo este valor comparable al máximo (2.4 µg-at-N-NO3=/L) encontrado en la surgencia de La Guajira (Corredor, 1979), es decir, niveles muy por debajo de lo encontrado en la presente investigación.
Considerando el proceso de nitrificación propuesto por Riley y Chester (1989) en el cual se establece que la oxidación del amonio en buenas concentraciones de oxígeno (como se presenta en las masas de agua en estudio), conduce a la formación de nitritos y un proceso posterior de oxidación origina los nitratos, se espera un proceso constante de nitrificación y una tendencia definida a inclinarse el nitrógeno a la forma de nitratos (Franco-Herrera, 2005). En respuesta a este precedente, la forma intermedia de nitritos se mantuvo en bajas concentraciones sin importar el período climático.
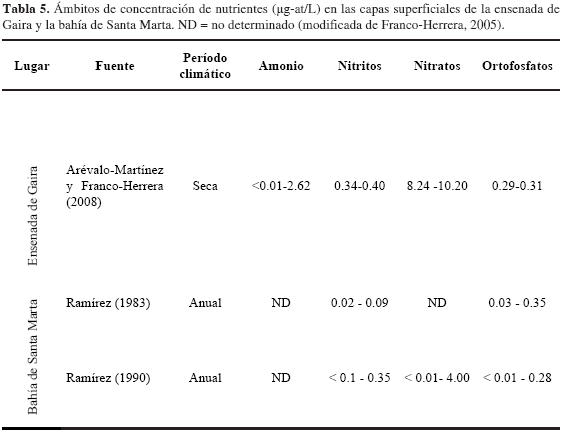
Por su parte, el amonio exhibió las concentraciones más bajas de todos los nutrientes en consideración (0.04-5.88 µg at NH4+/L), en concordancia con mediciones hechas por Arévalo-Martínez y Franco-Herrera (2008) para la época seca del año para la ensenada de Gaira (Tabla 5). Pese a que el amonio es la forma preferida de nitrógeno para el fitoplancton por no requerir una reducción química para ser usado en la síntesis de proteínas y demandar menor consumo de energía celular (Balech, 1977; Álvarez, 1981), el evento de surgencia no proporcionó una alta disponibilidad de este importante nutriente o en su defecto, como se indicó anteriormente, los procesos de nitrificación favorecidos por altas concentraciones de oxígeno disuelto en la columna de agua, llevan a una oxidación de este compuesto a la forma de nitratos.
De otro lado, las concentraciones de fosfatos fueron relativamente altas (> 0.3 µg at/L; Millero, 2006), reflejando a este nutriente para la condición ambiental evaluada como no biolimitante, por lo tanto el crecimiento de la comunidad autotrófica pelágica, sería independiente de la concentración de fósforo, o por el contrario, los muestreos se adelantaron durante fases tempranas de la surgencia, donde permanece alta la concentración de ortofosfatos y bajas la biomasa fitoplanctónica (Millero, 2006). En comparación con estudios realizados para la ensenada de Gaira y Santa Marta, los resultados actuales (2.59 y 4.68 µg-at-P-PO4=/L) se presentan en mayores concentraciones (Tabla 5). Lo anterior corrobora la intensidad del evento de surgencia comprendido entre los meses de febrero-abril de 2008 Entre tanto, para los meses de mayo y junio, correspondientes a la época lluviosa menor, los niveles de ortofosfatos fueron levemente menores, oscilando entre 2.59 y 3.79 µg-at-P-PO4=/L y superaron mediciones para la misma época (Tabla 5). Si bien, todo los parámetros anteriormente descritos, demuestran la variabilidad temporal de la dinámica física y química de la columna de agua, no se presentaron diferencias contrastantes entre los períodos de surgencia y no surgencia considerados inicialmente, ni tampoco entre los sitios muestreados, lo que pone en evidencia una masa de agua homogénea inherente a la inestabilidad atmosférica durante el período de investigación, que impidió la marcada estacionalidad de las épocas seca mayor y lluviosa menor.
Resulta importante notar que la disponibilidad de nutrientes fue un indicador primario de condiciones ideales que podrían influenciar la biomasa fitoplanctónica en eventos de surgencia a lo largo de zona costera del departamento del Magdalena. El aporte de la comunidad autotrófica estuvo entre los ámbitos registrados para la costa Caribe colombiana, por ejemplo comparados con sectores de la alta y media Guajira los valores fluctúan entre 0.76 y 1.52 µg/L (Duarte, 1996), o en el Parque Nacional Naturales Corales del Rosario, donde la biomasa fitoplanctónica está entre 0.1 y 0.8 µg/L (Franco-Herrera et al., 1992; Gualteros et al., 1992).
Las concentraciones medias de clorofila a a lo largo de las ensenadas y bahías fueron homogéneas, demostrando la ausencia de diferencias espaciales entre el sector sur y norte de las aguas costeras del departamento del Magdalena. Sin embargo, se presentaron concentraciones superiores en época de surgencia a diferencia de estudios realizados en la ensenada de Gaira y el golfo de Salamanca por Franco-Herrera (2001) y Tigreros (2001) quienes encuentran concentraciones mayores de este pigmento en época de lluvias debido al fuerte aporte continental, con valores que fluctúan entre 0.76 y 9.6 µg/L. Igualmente, Arévalo-Martínez y Franco-Herrera (2008) para el período septiembre-noviembre registran valores entre < 0.1 y 2.22 µg/L. En este sentido, se esperaría encontrar mayores biomasas en época de lluvias, sin embargo, el probable bajo aporte de aguas continentales analizado anteriormente y la mayor oferta de compuestos inorgánicos durante los eventos de surgencia favorecieron más las concentraciones de clorofila a durante esta última condición.
Entre tanto, se presentó una correlación negativa entre la biomasa fitoplanctónica y los nitratos (r= -0.28. p < 0.05), lo cual podría representar un desacople o respuesta de asimilación tardía por parte de la comunidad autotrófica frente a la fertilización y explicar el predominio de valores relativamente bajos de biomasa en el presente estudio, comparados con los registrados para otros sistemas de la gran cuenca Caribe (Franco-Herrera, 2005; Arévalo-Martínez y Franco-Herrera, 2008). Por su parte, la salinidad y temperatura se correlacionaron negativamente (r= -0.31, p < 0.05 y -0.32, p < 0.05, respectivamente) con la biomasa, reflejando parcialmente la interdependencia de la magnitud de clorofila a con los parámetros físicos.
En estudios realizados anteriormente para el mar Caribe occidental se corroboran algunas de las relaciones anteriormente descritas, como la influencia de las variables temperatura, salinidad y densidad sobre las fluctuaciones de clorofila a. Al respecto, Franco-Herrera et al. (2006) para el Parque Nacional Natural Tayrona y el golfo de Salamanca atribuyen los cambios en las concentraciones de clorofila a, composición y fisiología del fitoplancton a las mismas condiciones oceanográficas en mención. A estos se suman los nitratos, lo que pone en evidencia que los efectos de la surgencia estacional del área no son sólo físicos, si no también químicos y confirma de la misma manera la importancia de dichos parámetros sobre la biomasa fitoplanctónica en ausencia de pulsos de surgencia.
Todo lo anterior señala al área como un sistema tropical afectado por la variabilidad de los procesos oceanográficos típicos de sistemas costeros, con características mesotróficas y confirma en una escala espacial y temporal más amplia la nueva visión planteada por Arévalo-Martínez y Franco-Herrera (2008), frente a la alta dinámica oceanográfica de este sistema tropical, como su adecuada oferta de alimento autotrófico para las redes tróficas marinas. Así mismo, al parecer para el primer semestre de 2008, las condiciones atmosféricas y oceanográficas encontradas, indican una clara homogeneidad temporal como espacial, sin una clara diferenciación entre épocas climáticas, tal como se ha considerado históricamente para la región del departamento del Magdalena.
AGRADECIMIENTOS
Los autores agradecen a la Universidad de Bogotá Jorge Tadeo Lozano (UJTL), especialmente a la Dirección de Investigaciones, a la Facultad de Ciencias Naturales, Programa de Biología Marina, Bogotá-Santa Marta, por el apoyo logístico y financiero en la ejecución de esta investigación. La presente investigación fue financiada en un 100 % por la Dirección de Investigaciones de la UJTL, e hizo parte del Proyecto Código 96/03/07.
BIBLIOGRAFÍA
1 Alexander, M., I. Bladez, M. Mewman, J. Lanzante, N. Cheung y J. Scotti. 2002. The atmospheric bridge: the influence of ENSO teleconnections on air-sea interactions over the global oceans. J. Clim., 15: 1-30. [ Links ]
2 Álvarez, F. 1981. Introducción a la ecología marina. Fascículos de Ecología II. Mejoras, Barranquilla. 90 p. [ Links ]
3 Andrade, C. 2001. Las corrientes superficiales de la cuenca de Colombia observadas con boyas de deriva. Rev. Acad. Col. Cien., 25: 321-335. [ Links ]
4 Arévalo-Martínez, D. y A. Franco-Herrera. 2008. Características oceanográficas de la surgencia frente a la ensenada de Gaira, departamento del Magdalena, época seca menor de 2006. Bol. Invest. Mar. Cost., 37 (2): 131-162. [ Links ]
5 Balech, E. 1977. Introducción al fitoplancton marino. Universitaria, Buenos Aires. 194 p. [ Links ]
6 Bell, G. y M. Chelliah. 2005. Leading tropical modes associated with interannual and multidecadal fluctuations in north Atlantic hurricane activity. J. Clim., 19: 590-612. [ Links ]
7 Blanco, J. 1988. Las variaciones ambientales estacionales en las aguas costeras y su importancia para la pesca de la región de Santa Marta, Caribe colombiano. Tesis de Maestría Biol. Univ. Nacional de Colombia, Bogotá. 62 p. [ Links ]
8 Blanco, J., J. Díaz, G. Ramírez y M. Cortés. 1994. El banco de las Ánimas: una amplia formación arrecifal desarrollada sobre un antiguo delta del río Magdalena. Bol. Ecotrópica, 27: 10-18. [ Links ]
9 Bula-Meyer, G. 1985. Un núcleo nuevo de surgencia en el Caribe colombiano detectado en correlación con las macroalgas. Bol. Ecotrópica, 14: 3-20. [ Links ]
10 Bula-Meyer, G. 1990. Oceanografía. 100-113. En: Jimeno, M. C. (Ed.). Caribe Colombia. Primera edición, Fondo FEN-Colombia, Folio Ltda., Bogotá. 271 p. [ Links ]
11 Cabrera, E. y M. Donoso. 1993. Estudio de las características oceanográficas del Caribe colombiano, Región III, Zona 1. PDCTM. Bol. Cient. Cent. Invest. Oceanogr. Hidrogr., 13: 19-32. [ Links ]
12 CIMSS. 2008. Tropical cyclones. Cooperative Institute for Meteorological Satellite Studies. http://www.cmiss.ssec.wisc.edu/tropic/. 06/30/08. [ Links ]
13 CIOH. 2008. Programa SARPAR. Sistema Automatizado de Recepción y Pronóstico del CIOH. http://www.sarpar.cioh.org.co/SARPAR/goes.php. 06/30/08. [ Links ]
14 Cognetti, G. M. y G. Magazzú. 2001. Biología marina. Editorial Ariel S.A., Barcelona. 619 p. [ Links ]
15 Corredor, J. E. 1979. Phytoplankton response to low level nutrients enrichment through upwelling in the Colombian Caribbean Basin. Deep-Sea Res., 26: 731-741. [ Links ]
16 Corredor, J. E. 1992. Dinámica espacio-temporal de la surgencia de La Guajira. 206-221. En: CCO (Ed.). Memorias VIII Seminario Nacional de Ciencias y Tecnologías del Mar. (CCO), Santa Marta. 1144 p. [ Links ]
17 Covey, D. y S. Hastenrath. 1978. The Pacific El Niño phenomenon in the Atlantic sector. Mon. Weather Rev., 106: 1280-1287. [ Links ]
18 Donoso, M. 1990. Circulación de las aguas en el mar Caribe. 345-354. En: CCO (Ed.). Memorias VII Seminario Nacional de Ciencias y Tecnologías del Mar. Cali. 792 p. [ Links ]
19 Duarte, G. 1996. Estructura del fitoplancton al noreste de La Guajira, durante el crucero Caribe 1-93. Trabajo de grado Biol. Mar. Univ. Jorge Tadeo Lozano, Bogotá. 98 p. [ Links ]
20 Fajardo, G. 1979. Surgencia costera en las proximidades de la península colombiana de La Guajira. Bol. Cient. Cent. Invest. Oceanogr. Hidrogr., 2: 7-19. [ Links ]
21 Fernández, A. 2003. Tecnología marina (Gran enciclopedia del mar). Carrogio editores, Barcelona. 128 p. [ Links ]
22 Franco-Herrera, A. 2001. Estrategias de ingestión y flujo de carbono entre el fitoplancton, seston total y Eucalanus spp. (Copepoda: Calanoidea) en dos ambientes oceanográficos del Caribe central colombiano. Informe final, Univ. Jorge Tadeo Lozano, Santa Marta. 67 p. [ Links ]
23 Franco-Herrera, A. 2005. Oceanografía de la ensenada de Gaira: El Rodadero, más que un centro turístico en el Caribe. Univ. Jorge Tadeo Lozano, Bogotá. 58 p. [ Links ]
24 Franco-Herrera, A., D. Avendaño, W. Bernal y M. Uribe. 1992. Contribución al conocimiento del plancton de isla Tesoro, Parque Nacional Natural Corales del Rosario, Caribe colombiano. 754-762. En: CCO (Ed.). Memorias VIII Seminario Nacional de Ciencias y Tecnologías del Mar y 1er. Congreso Centroamericano y del Caribe de Ciencias del Mar, Santa Marta. 1144 p. [ Links ]
25 Franco-Herrera, A., L. Castro y P. C. Tigreros. 2006. Plankton dynamics in the south-central Caribbean sea: strong seasonal changes in a coastal tropical system. Caribb. J. Sci., 42 (1): 24-38. [ Links ]
26 Garay, T. J., G. Ramírez, J. Betancourt, B. Marín, B. Cadavid, L. Panizzo, L. Lesmes, J. Sánchez, H. Lozano y A. Franco (Eds.). 2003. Manual de técnicas analíticas para la determinación de parámetros físico-químicos y contaminantes marinos: aguas, sedimentos y organismos. Serie de documentos generales No. 13, Instituto de Investigaciones Marinas y Costeras-INVEMAR, Santa Marta. 177 p. [ Links ]
27 Google Earth. 2008. Beta 4.1. http://www.google.com. 22/01/08. [ Links ]
28 Gualteros, W., C. Hernández, A. Castillo y A. Fresneda. 1992. Clorofila y seston en la laguna costera de Cocoliso, Isla Grande, Parque Nacional Natural Corales del Rosario. 512-521. En: CCO (Ed.). Memorias VIII Seminario Nacional de Ciencias y Tecnologías del Mar y Congreso Centroamericano y del Caribe de Ciencias del Mar, Santa Marta. 1144 p. [ Links ]
29 IDEAM. 2008. Pronóstico de pleamares y bajamares costa Caribe colombiana. Informe técnico, (IDEAM), Bogotá. 123 p. [ Links ]
30 INVEMAR. 2004. Informe del estado de los ambientes marinos y costeros en Colombia. Serie de Publicaciones Periódicas No.8, Santa Marta. 327 p. [ Links ]
31 INVEMAR, The Nature Conservancy, Environmental Defense, UAESPNN, UNEPCAR/ECU, ICRAN y Caribbean Environment. 2004. Primer curso nacional de áreas marinas protegidas en Colombia. Informe final, Santa Marta. 48 p. [ Links ]
32 Márquez, G. 1982. Los sistemas ecológicos marinos del sector adyacente a Santa Marta, Caribe colombiano I: Generalidades. Ecol. Trop., 2 (1): 5-18. [ Links ]
33 Millero, F. 2006. Chemical oceanography. Tercera edición. CRC Press LLC, Boca Ratón, EE. UU. 496 p. [ Links ]
34 Müller, K. 1979. Interrelaciones entre salinidad y temperatura en la bahía de Santa Marta. An. Inst. Invest. Mar. Punta de Betín, 11: 219-226. [ Links ]
35 NOAA (National Oceanography and Atmosphere Agency). 2008. Changes to the Oceanic Niño Index. National Weather Service. Climate Prediction Center. http://www.cpc.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.shtml. 01/12/06 a 01/04/08. [ Links ]
36 Pond, S. y G. L. Pickard. 1983. Introductory dynamical oceanography. Butterwotrh Heinemann, Oxford. 329 p. [ Links ]
37 Ramírez, G. 1983. Características fisicoquímicas de la bahía de Santa Marta (agosto, 1980-julio, 1981). An. Inst. Invest. Mar. Punta de Betín, 13: 111-121. [ Links ]
38 Ramírez, G. 1987. Características hidrodinámicas y composición química de los sedimentos de la bahía de Neguanje, Caribe colombiano. Bol. Invest. Mar. Cost., 17: 15-26. [ Links ]
39 Ramírez, G. 1990. Distribución de los nutrientes inorgánicos en las aguas costeras de la región de Santa Marta, Caribe colombiano. 244-254. En: CCO (Ed.). Memorias VII Seminario Nacional de Ciencias y Tecnologías del Mar, Cali. 792 p. [ Links ]
40 Riley, J. P. y R. Chester. 1989. Introducción a la química marina. AGT editor, México, D. F. 459 p. [ Links ]
41 Salamanca, M. 1996. Curso de química marina, guía de trabajos prácticos. Laboratorio de química Marina. Univ. Concepción, Concepción, Chile. 68 p. [ Links ]
42 Salzwedel, H. y K. Müller. 1983. A summary of meteorological and hydrological data from the Bay of Santa Marta, Colombian Caribbean. An. Inst. Invest. Mar. Punta de Betín, 13: 67-83. [ Links ]
43 Sánchez, R., G. Vargas, H. González y D. Pabón. 2001. Los fenómenos cálido del Pacífico (El Niño) y frío del Pacífico (La Niña) y su incidencia en la estabilidad de laderas en Colombia. III Simposio Panamericano de Deslizamientos. Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM), Bogotá D.C. 12 p. [ Links ]
44 Strickland, D. H. y T. Parsons. 1972. A practical handbook of sea water analysis. Bulletin Fisheries Research Board of Canada, Ottawa, Canadá. 310 p. [ Links ]
45 Tigreros, P. C. 2001. Biodiversidad y valoración bioquímica del fitoplancton marino en ambientes costeros mesotróficos y oligotróficos tropicales, Caribe colombiano. Trabajo de grado Biol. Mar. Univ. Jorge Tadeo Lozano, Santa Marta. 173 p. [ Links ]
46 Trenberth, K. 2003. The El Niño-Southern Oscillation (ENSO) System: 59-63. En: Potter, T. D. y B. R. Colman (Eds.). Handbook of weather, climate and water: dynamics, climate, physical meteorology, weather systems, and measurements. John Wiley and Sons, Chicago. 973 p. [ Links ]
47 USF (University of South Florida). 2008. Sea surface temperature daily. Institute for Marine Remote Sensing (IMARS). http://www.imars.usf.edu. 20/06/08. [ Links ]
48 Vanegas, T. 2002. Riqueza de órdenes y dinámica del mesozooplancton marino en ambientes costeros del Golfo de Salamanca y el Parque Tayrona, Caribe Colombiano. Trabajo de grado Biol. Mar., Univ. Jorge Tadeo Lozano, Santa Marta. 110 p. [ Links ]
49 Zar, H. 1999. Biostatisticial analysis. Cuarta edición. Prentice Hall, New Jersey. 665 p. [ Links ]
FECHA DE RECEPCIÓN: 01/12/2008 FECHA DE ACEPTACIÓN: 10/08/2010
