










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.39 no.2 Santa Marta July/Dec. 2010
CHRYSOGORGIIDAE (OCTOCORALLIA) RECOLECTADOS DURANTE LAS EXPEDICIONES MACROFAUNA I Y II EN EL CARIBE COLOMBIANO*
CHRYSOGORGIIDAE (OCTOCORALLIA) COLLECTED DURING THE MACROFAUNA I Y II EXPEDITIONS TO THE COLOMBIAN CARIBBEAN
Isabel Cristina Chacón-Gómez1, Nadiezhda Santodomingo1, Javier Reyes1 y Camilo Ernesto Andrade Sossa2
1 Instituto de Investigaciones Marinas y Costeras José Benito Vives De Andreis-INVEMAR, Cerro Punta de Betín, Santa Marta, Colombia. icchacongomez@gmail.com (I.C.C.G.)
2 Natural History Museum, Cromwell Road, SW7 5BD, Londres, Reino Unido.
RESUMEN
Se describen ejemplares de cuatro especies de octocorales de la familia Chrysogorgiidae (Chrysogorgia desbonni, C. elegans, C. thyrsiformis y Trichogorgia lyra) recolectados en el Caribe colombiano durante las exploraciones INVEMAR-Macrofauna I y II. Las tres especies de Chrysogorgia se encontraron entre 160 y 516 m de profundidad, mientras que T. lyra se recolectó en aguas someras entre 20 y 150 m de profundidad, ámbitos batimétricos anteriormente poco estudiados en el país, por lo que es un estudio de particular interés. Además se han identificado alrededor de 50 especies de diferentes octocorales a partir de este tipo de estudios. Chrysogorgia thyrsiformis se registra por primera vez en el Caribe colombiano. Por otro lado, se comenta la distribución geográfica y el ámbito batimétrico de todas estas especies en el Caribe y particularmente para Colombia.
PALABRAS CLAVE: Octocorales, Chrysogorgiidae, Taxonomía, Caribe, Colombia.
ABSTRACT
Four samples of octocorals in the family Chrysogorgiidae are described (Chrysogorgia desbonni, C. elegans and C. thyrsiformis and Trichogorgia lyra), collected in the Colombian Caribbean during the explorations INVEMAR-Macrofauna I and II. The three Chrysogorgia species were found between depths of 160 and 516 m, while T. lyra was collected in shallow waters between depths of 20 and 150 m, both bathymetric ranges which had not been widely studied in the country before, making this a particularly interesting study. Furthermore, nearly 50 species of various octocorals have been identified based on studies of this type. Chrysogorgia thyrsiformis is recorded for the first time for Colombian Caribbean. In the other hand, geographic distribution and bathymetric ranges are indicated for all these species in the Caribbean and particularly Colombia.
KEY WORDS: Octocorals, Chrysogorgiidae, Taxonomy, Caribbean, Colombia.
INTRODUCCIÓN
Los octocorales (Anthozoa: Octocorallia), conocidos comúnmente como corales blandos, abanicos de mar y plumas de mar, cuentan más de 2000 especies en el mundo, constituyendo la subclase más diversa de antozoos. Están presentes en casi todos los ambientes marinos, cumplen un importante papel ecológico (Sánchez et al., 1998; Krieger, 2001; Witherell y Coon, 2001) y tienen un amplio potencial económico en materia de bioprospección de productos naturales marinos (Pérez-Gil et al., 1999). En Colombia los estudios morfológicos, ecológicos y taxonómicos han sido realizados principalmente sobre especies arrecifales de aguas someras, destacándose los trabajos de Botero (1987), Sánchez (1994, 1998, 1999) y Sánchez et al. (1998), mientras que las especies de ambientes más profundos han sido poco estudiadas (Lattig-Matiz, 2000; Reyes et al., 2005).
Con el fin de caracterizar la macrofauna de la plataforma y talud continental a lo largo del Caribe colombiano (20-500 m), el Instituto de Investigaciones Marinas y Costeras-INVEMAR, en asocio con la Universidad Nacional de Colombia, llevó a cabo dos importantes proyectos entre 1998 y 2000, Macrofauna I y II. En las colectas realizadas hasta entonces, se identificaron 45 especies de octocorales siendo la familia Chrysogorgiidae Verrill, 1883 de particular interés; todas las especies de Chrysogorgia, Duchassaing y Michelotti, 1864 encontradas en este estudio se recolectaron entre 160 y 516 m de profundidad, ámbitos batimétricos anteriormente poco estudiados en el país. Adicionalmente, esta familia incluye algunas de las especies de octocorales más llamativas por su particular iridiscencia y sus patrones de ramificación, que son matemáticamente precisos (Cairns, 2001). De los 12 géneros que la conforman (Williams, 2001), sólo Chrysogorgia y Trichogorgia Hickson, 1904 se encuentran en el Caribe colombiano.
El conocimiento previo que se tiene de la distribución de la familia para el Caribe colombiano se basa en el trabajo de Cairns (2001), quien registra la presencia de C. elegans y C. fewkesii de acuerdo con material recolectado durante las expediciones del "Pillsbury", depositado en el National Museum of Natural History (NMNH) del Instituto Smithsonian en Washington. Reyes et al. (2005) registran la presencia de C. desbonni frente a Santa Marta (200 m de profundidad), asociada a comunidades de corales azooxantelados. En cuanto al género Trichogorgia, Bayer y Muzik (1976) describieron a T. lyra con material proveniente de Cabo Tiburón y el golfo de Morrosquillo.
En este trabajo se describen especímenes de C. desbonni, C. thyrsiformis, C. elegans y T. lyra, recolectados en las expediciones INVEMAR-Macrofauna I y II; C. thyrsiformis se registra por primera vez en el Caribe colombiano. Además, se comenta la distribución geográfica y el ámbito batimétrico de estas especies en el Caribe y particularmente para Colombia.
MATERIALES Y MÉTODOS
Área de estudio
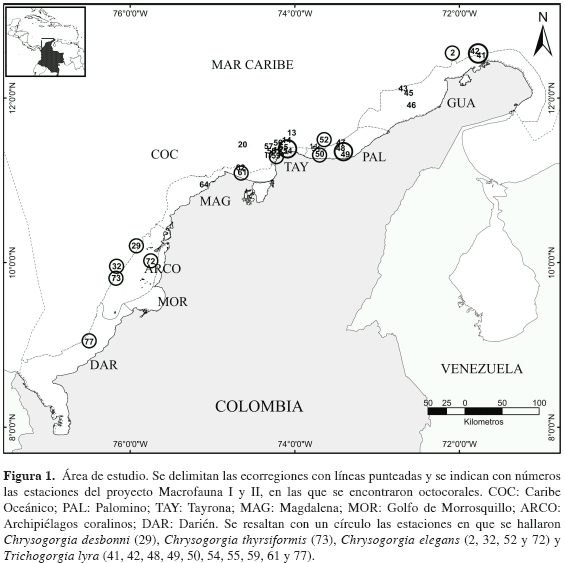
El material se recolectó durante las expediciones INVEMAR-Macrofauna I y II, en ocho de las nueve ecorregiones (Figura 1) que hacen parte de la sectorización del Caribe colombiano (Díaz y Gómez, 2000). Las ecorregiones se definen según los siguientes criterios: grado de influencia continental; amplitud, rasgos geomorfológicos y tipos de sedimentos de la plataforma continental; rasgos geomorfológicos de la costa; ocurrencia estacional o permanente de afloramientos de masas de agua subsuperficiales; grado de exposición o nivel de energía del oleaje; presencia de unidades ecológicas particulares o mosaicos de éstas; productividad biológica de la columna de agua y de los ecosistemas costeros (Díaz y Gómez, 2000).
Muestreo
Se diseñó un muestreo sistemático seleccionando puntos sobre las cartas de navegación (CIOH, 1991; Defense Mapping Agency Hydrographic/Topographic Center, 1990, 1995a, 1995b) distribuyendo las estaciones frente a cada ecoregión costera a 20, 70, 150, 300 y 500 m de profundidad (Navas et al., 2002; Reyes et al., 2005). Las campañas se desarrollaron en el buque de investigaciones "B/I Ancón", perteneciente al Instituto de Investigaciones Marinas y Costeras-INVEMAR. Los muestreos se realizaron con una red de arrastre demersal de cuatro tapas, tipo semibalón, con dos puertas metálicas tipo "V", diseñada y construida por Marinovich Trawl Co., Inc. (Navas et al., 2002; Reyes et al., 2005). El material fue fijado y preservado en etanol al 70 y 96 %.
Identificación y descripción
El material se identificó y describió mediante disecciones de los diferentes fragmentos de las muestras y el montaje de escleritos. Para la observación de escleritos se tomaron fragmentos por separado de cenénquima de los tallos, cálices, ejes y pólipos. Para limpiar los escleritos calcáreos del tejido blando se montó cada fragmento en una lámina portaobjetos con el suficiente hipoclorito (NaClO) comercial para cubrir el material (Bayer, 1961). Se aplicó glicerina para mantener los escleritos en un medio líquido y facilitar su observación desde diferentes ángulos al microscopio.
Las imágenes fueron obtenidas con una cámara Nikon D-1 adaptada a un microscopio (para observar los escleritos) o al estereoscopio (para los detalles de los cálices). Las imágenes de las colonias se tomaron en acuarios de vidrio con fondo negro y se editaron usando Corel Photopaint 10.0. A través del programa Scion Image se midieron los escleritos y sus ornamentaciones. La identificación se realizó con base en los trabajos de Deichmann (1936), Bayer (1961, 1981), Bayer y Muzik (1976), Bayer y Stefani (1988) y Cairns (2001), y mediante la comparación con especímenes donados de C. elegans y el material tipo de T. lyra depositados en el NMNH.
RESULTADOS Y DISCUSIÓN
Se examinaron 45 especies de octocorales pertenecientes a nueve familias. Para Chrysogorgiidae se describen ejemplares de Chrysogorgia desbonni, C. thyrsiformis, C. elegans y Trichogorgia lyra. Las especies del género Chrysogorgia se distribuyen en aguas profundas, a partir de 160 m, mientras que T. lyra se encontró principalmente en aguas someras (entre 20 y 150 m). Adicionalmente, C. desbonni y C. thyrsiformis se recolectaron únicamente en Santa Marta y el archipiélago de San Bernardo, respectivamente, haciendo parte de la diversa fauna asociada a las comunidades de corales azooxantelados registradas para estas dos localidades (Reyes et al., 2005). Chrysogorgia elegans y T. lyra presentaron un ámbito más amplio a lo largo del Caribe colombiano, distribuyéndose principalmente en fondos blandos desde La Guajira hasta Darién.
Taxonomía
Subclase OCTOCORALLIA Haeckel, 1866
Orden ALCYONACEA Lamoroux, 1894
Suborden CALCAXONIA Grasshoff, 1999
Familia CHRYSOGORGIIDAE Verrill, 1883
Género Chrysogorgia Duchassaing y Michelotti, 1864
Especie tipo: Chrysogorgia desbonni Duchassaing y Michelotti, 1864
Diagnosis: Ramificación simpodial. Las ramas se dividen repetidas veces de manera dicotómica; se localizan formando dos abanicos paralelos entre sí sobre un eje principal corto, o se originan alrededor de un tallo principal cuyo eje tiene forma de espiral regular e individual. Los pólipos son grandes con relación a las ramas, poco abundantes y dispuestos dispersamente. Los escleritos son escamas, bastones y husos. El eje presenta un brillo metálico, comúnmente color ámbar, amarillo o dorado (Deichmann, 1936; Bayer, 1956; Bayer y Stefani, 1988; Williams, 1992; Cairns, 2001).
Distribución: Las especies de este género se distribuyen en aguas profundas de los océanos Atlántico e Indo-Pacífico (Williams, 1992).
Chrysogorgia desbonni Duchassaing y Michelotti, 1864
(Figuras 2 y 3)
Chrysogorgia desbonni: Cairns 2001: 772.
Sinonimias: Chrysogorgia occidentalis Versluys 1902; Chrysogorgia desbonhi Grygier 1984 (Deichmann, 1936: 228; Cairns, 2001: 772).
Referencias: Deichmann, 1936: 222, 228-229; Cairns, 2001: 772, 784.
Material examinado: INV CNI1412 (10°9'12'' N, 76°0'24'' W).
Diagnosis: Esta especie se reconoce fácilmente porque presenta forma de crecimiento flabelada, ramificada a manera de dos abanicos paralelos y ramas anastomosadas en la mayoría de los casos.
Descripción: Se examinaron una colonia y varios fragmentos de hasta 8.5 cm de altura por 6 cm de ancho. Cada división dicotómica genera dos ramificaciones de grosor diferente, presentando una dicotomía irregular (Figura 2A, B, C). Ramas con forma circular en sección transversal, diámetro promedio de 0.643 mm, que disminuye gradualmente a medida que las ramificaciones se subdividen. Eje dorado iridiscente, con un diámetro promedio de 0.431 mm, que disminuye gradualmente hacia el ápice de la colonia.
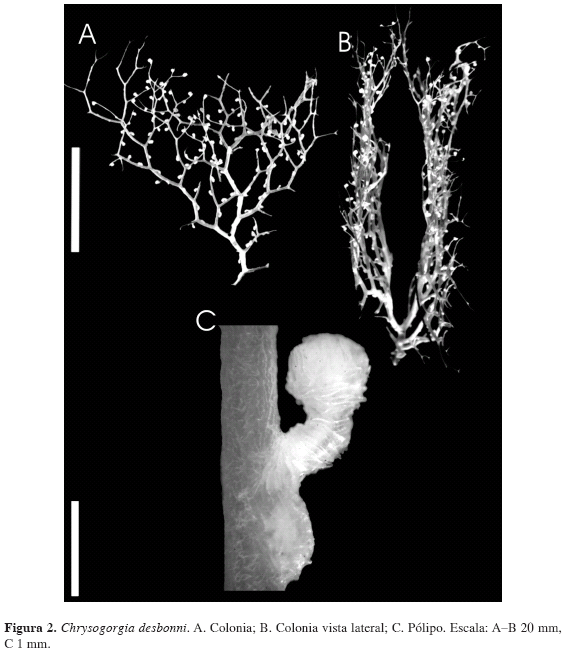
El cenénquima de toda la colonia está compuesto por escamas con formas irregulares, por lo general alargadas y con protuberancias laterales que pueden llegar a parecer, en algunos casos, ramificaciones de la escama (Figura 3A); escamas con longitud promedio de 0.278 mm y ancho mayor promedio de 0.056 mm, con ornamentaciones de 30 μm de altura promedio. El cenénquima en los cálices se compone principalmente de husos curvos a manera de "C", algunos con formas irregulares y otros ramificados (Figura 3B); en promedio, presentan 0.508 mm de longitud y 0.069 mm de ancho mayor, con ornamentaciones de 9 μm de altura. En todos los escleritos mencionados se presenta ornamentación espinosa y/o rugosa.
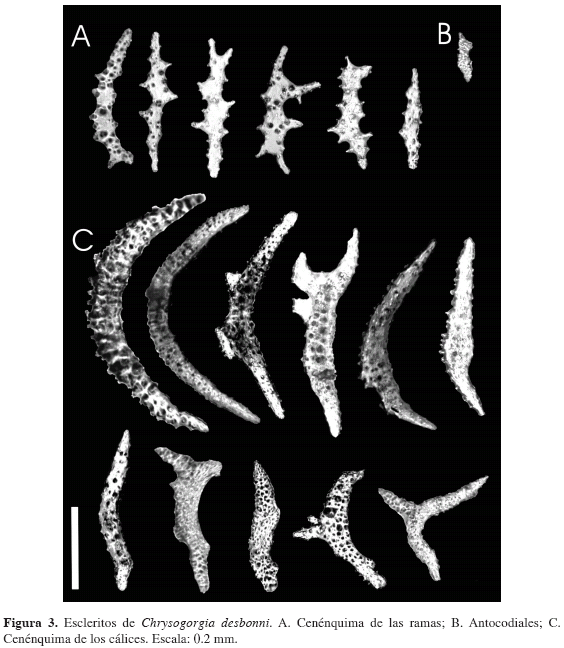
Cálices cilíndricos con ápice engrosado (Figura 2C), dispuestos uniforme e individualmente alrededor de todas las ramas, con uno o dos pólipos por subdivisión de las ramificaciones; en promedio los cálices tienen altura de 0.9 mm, diámetro del área basal de 0.358 mm y diámetro del ápice de 0.599 mm. Los escleritos antocodiales (porción blanda del pólipo) son principalmente husos pequeños (Figura 3C), en promedio de 0.371 mm de longitud y 0.066 mm de ancho mayor, con ornamentaciones espinosas y/o rugosas de 0.013 mm de altura mayor. Color café claro a crema en alcohol al 70 %; los cálices son más claros que el resto de la colonia.
Discusión: El material colombiano es de menor tamaño en comparación con especímenes del resto del Atlántico occidental, donde las colonias pueden alcanzar hasta 16 cm de alto y 12 cm de ancho, con un diámetro en los tallos de 1.6 mm (Cairns, 2001). No obstante, se identifican como C. desbonni por su inconfundible y característica forma flabelada en dos abanicos paralelos (Figura 2A); adicionalmente, la distribución geográfica y el ámbito batimétrico se encuentran dentro de los conocidos para la especie. Aparentemente todos los fragmentos pertenecen a la misma colonia de un tamaño un poco mayor al descrito (>8.5 cm de altura), pero no tan grandes como las descritas por Cairns (2001). Se presentaron dificultades en confirmar que el eje basal se ramifica bajo la fórmula 2/5 R (Cairns, 2001) debido a que la base de la colonia se encontró incompleta.
Distribución geográfica: Distribución continua desde el atolón de Turneffe, Belice, en dirección de las manecillas del reloj, a través de las Antillas hasta la isla de Granada, incluyendo las Bahamas. En el Caribe colombiano se encontró frente al Archipiélago de San Bernardo en la ecoregión ARCO.
Ámbito batimétrico: Chrysogorgia desbonni se encuentra entre 153 y 595 m de profundidad (Deichmann, 1936; Cairns, 2001). En el área de estudio se encontró entre 290 y 296 m.
Chrysogorgia thyrsiformis Deichmann, 1936
(Figuras 4 y 5)
Chrysogorgia thyrsiformis Deichmann, 1936: Cairns, 2001: 776.
Sinonimias: Chrysogorgia desbonni var. thyrsiformis Deichmann, 1936; Chrysogorgia elisabethae Bayer 1951 (Deichmann, 1936: 230; Cairns, 2001:776).
Referencias: Deichmann, 1936: 230; Bayer, 1951: 269-273; Cairns, 2001: 776-784, figs. 13-14.
Material examinado: INV CNI1739 (9°47'12''N, 75°13'45''W).
Diagnosis: Chrysogorgia thyrsiformis se reconoce por la forma arbustiva de las colonias, con un patrón de crecimiento de las ramas muy denso, más denso que en otras especies del género, pues mientras en C. thyrsiformis la distancia entre las ramas alrededor del eje principal es de máximo 0.7 mm, en las otras especies pueden estar separadas hasta por 6 mm entre sí. Así mismo, el intervalo entre las ramas alineadas no supera los 5 mm, mientras que en las otras especies este intervalo varía entre 5 y 25 mm; las ramificaciones pueden presentar hasta 12 divisiones dicotómicas y los escleritos en los cálices se disponen transversalmente.
Descripción: una colonia de 10 cm de alto y 9 cm de ancho de forma arbustiva; ramificaciones separadas por 0.5 a 0.625 mm, alrededor de un único tallo principal, robusto, con apariencia de un espiral por su crecimiento simpodial, con una a doce divisiones dicotómicas, no siempre simétricas (Figura 4A). El espacio entre las ramas que están alineadas es de 3.5 a 3.875 mm. Fórmula de ramificación 2/5 R. La colonia estaba firmemente adherida al coral escleractinio Madracis myriaster, recubriéndolo mediante un disco sólido y gris (Figura 4B). Tallo y ramas con forma circular en sección transversal (Figura 4A, B, C).
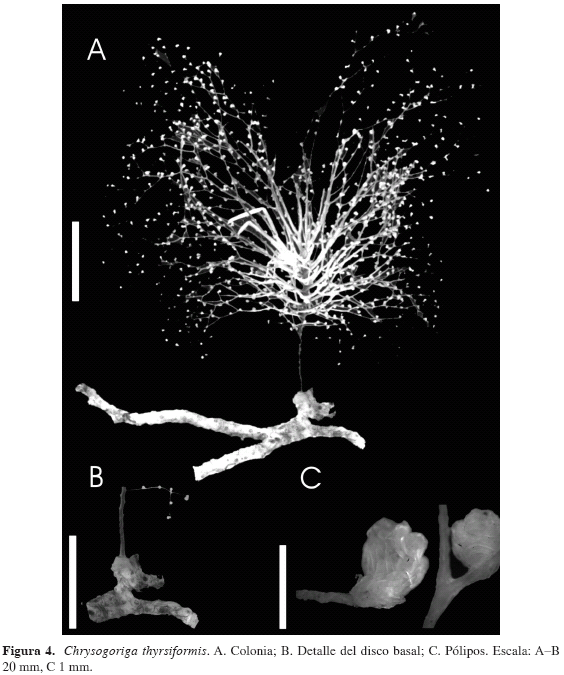
El eje del tallo principal presenta un diámetro promedio de 1.083 mm, disminuyendo gradualmente hacia la región apical de la colonia. El diámetro promedio de las ramas, incluyendo las terminales, es de 0.375 mm.
El cenénquima de la colonia está compuesto por escamas de 0.146 mm de longitud promedio y 0.054 mm de ancho mayor promedio, con formas variables, irregulares, por lo general alargadas, con bordes sutilmente aserrados y algunas ornamentaciones redondeadas poco notorias (Figura 5A). El cenénquima alrededor de los cálices se compone de escamas con formas más similares entre sí, igualmente las escamas antocodiales (Figura 5B), aunque también irregulares, con ornamentaciones tanto redondeadas como espinosas y en promedio de 0.356 mm de longitud y 0.113 mm de ancho mayor (Figura 5C).
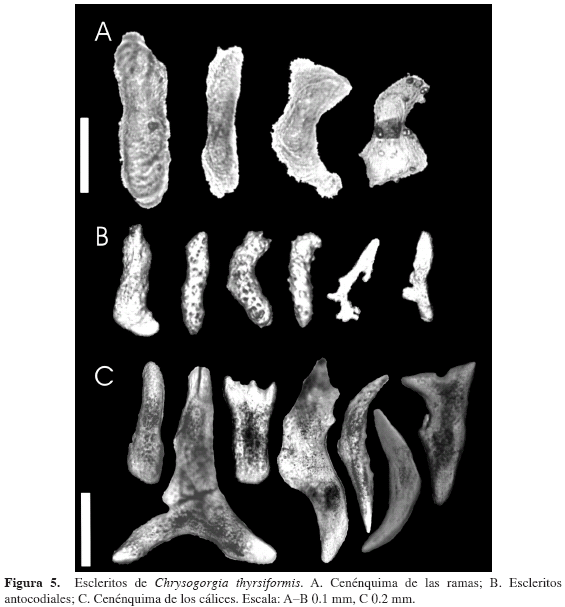
Cálices en forma de copa (Figura 4C) con escleritos organizados transversalmente. Pólipos individuales localizados alrededor de las ramas, entre uno y tres por subdivisión de las ramificaciones, relativamente equidistantes entre sí. En promedio, los cálices presentan una altura de 1.166 mm y un diámetro de 0.493 mm en la base y de 0.848 mm en el ápice. Por encima de los pólipos contraídos se observan ocho lóbulos correspondientes a los ocho tentáculos.
Los escleritos antocodiales son principalmente escamas alargadas, algunas con formas irregulares (Figura 5B), con un promedio de 0.117 mm de longitud y 0.032 mm de ancho mayor. Presentan ornamentaciones poco conspicuas, espinosas y/o rugosas.
El color de la colonia preservada en etanol al 96 % es bronce muy oscuro con visos iridiscentes en el tallo principal, dorado iridiscente claro en las ramas primarias el cual se acentúa hasta tornarse más oscuro y brillante en las ramas secundarias y terminales. Los cálices presentan un color que varía de crema claro a blanco.
Discusión: Los caracteres distintivos están claramente presentes en la colonia, tales como la distancia entre las ramas muy corta y entre las ramas alineadas, esta distancia no supera los 5 mm en ninguna parte de la colonia. Aunque según la descripción de Cairns (2001), las colonias de esta especie pocas veces alcanzan tamaños mayores a 8 cm de altura y 8 a 9 cm de ancho y la colonia del Caribe colombiano presenta mayores dimensiones (10x9 cm), al igual que en C. desbonni, esto puede atribuirse a un crecimiento diferencial determinado por las condiciones ambientales de la localidad, o por diferencias en la edad y grado de desarrollo que hayan alcanzado los ejemplares al momento del muestreo.
Chrysogorgia thyrsiformis fue descrita como una variación de C. desbonni (Deichmann, 1936). Cairns (2001) la eleva a nivel de especie debido a que la forma de la colonia, número de divisiones de las ramas, número de pólipos por división, formas de los escleritos de los cálices, distancias e intervalos entre las ramas y color del eje en el tallo principal se diferencian claramente de C. desbonni.
Distribución geográfica: Distribución continua desde el banco de Grand Bahama a través de las Antillas hasta la isla Tortuga, Venezuela, incluyendo la península de Yucatán y Bahamas. En Colombia se distribuye en la ecorregión Archipiélagos Coralinos-ARCO.
Ámbito batimétrico: Chrysogorgia thyrsiformis se encuentra entre 146 y 526 m de profundidad (Cairns, 2001). Dentro del área de estudio se encontró a 160 m de profundidad.
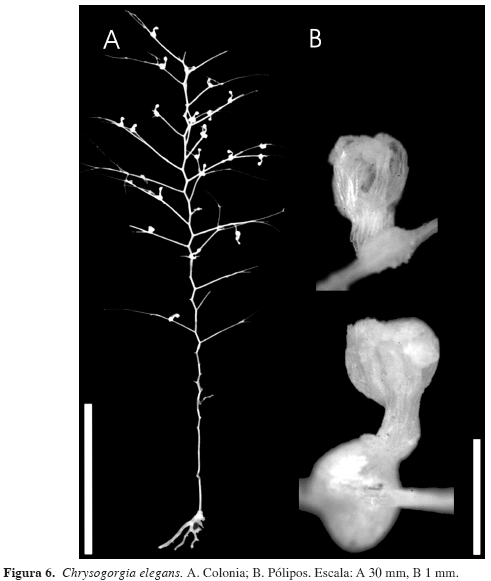
Chrysogorgia elegans Verrill
(Figuras 6 y 7)
Sinonimias: Dasygorgia elegans Verrill 1883; Dasygorgia spiculosa Verrill 1883; Chrysogorgia affinis Versluys 1902; Chrysogorgia flexilis Thomson 1927; Chrysogorgia agassizii Deichmann 1936 (Deichmann, 1936: 231; Cairns, 2001:760-762).
Referencias: Deichmann, 1936: 222, 228, 231; Bayer, 1951: 269-273; Cairns, 2001: 760, 784.
Material examinado: INV CNI1733 (9°48'58''N, 76°16'27''W). INV CNI1734 (9°48'58''N, 76°16'27''W). INV CNI1736 (11°27'64''N, 73°40'14''W). INV CNI1737 (9°48'58''N, 76°16'27''W). INV CNI1738 (9°48'58''N, 76°16'27''W). INV CNI1836 (11°27'06''N, 74°1'08''W). INV CNI1837 (12°31'48''N, 72°7'58''W). USNM 88109. USNM 89090. USNM 100884. USNM 52908. USNM 55919.
Diagnosis: Esta especie se caracteriza por su crecimiento simpodial que le da apariencia de espiral, las ramas presentan de una a tres divisiones dicotómicas y se fija al sustrato a través de un fragmento de aspecto rizoidal. El espacio entre ramas alineadas ("orthostiche interval") varía entre 7 y 7.5 mm. La longitud máxima alcanzada por las ramas es de alrededor de 25 mm, siendo siempre rígidas y quebradizas.
Descripción: Colonias de poca flexibilidad, hasta 12.5 cm de altura por 5 cm de ancho; arbustivas con ramificaciones simpodiales cada 1.74 a 2 mm, alrededor de un único tallo principal, dispuestas a manera de espiral, con divisiones (entre dos y cinco) dicotómicas, no siempre simétricas (Figura 6). El espacio entre las ramas alineadas ("orthostiche interval") es de 7.169 a 11.000 mm. Fórmula de ramificación 2/5 R. El fragmento de la colonia para fijación al sustrato es sólido, blanco y ramificado rizoidalmente. Tallo y ramas con forma circular en sección transversal, con un diámetro promedio de 0.423 mm, disminuyendo gradualmente hacia la región apical de las colonias y ramas terminales, las cuales presentan un diámetro promedio de 0.073 mm. Diámetro promedio del eje de 0.261 mm, disminuyendo gradualmente hacia la región apical de las colonias y ramas terminales. El cenénquima del cuerpo de la colonia está compuesto por escleritos a manera de escamas, con formas irregulares, por lo general alargadas, con bordes aserrados de 0.262 mm de longitud promedio, ancho mayor promedio de 0.046 mm y algunas ornamentaciones redondeadas difícilmente visibles (Figura 7).
El cenénquima en los cálices se compone principalmente de husos aplanados, en promedio de 0.262 mm de largo y 0.038 mm de ancho mayor, con ornamentaciones tanto redondeadas como espinosas de altura de 0.01 mm (Figura 7).
Cálices cilíndricos, constreñidos centralmente y con un notorio ensanchamiento en el área basal que se adhiere a las ramas; los escleritos en los cálices se encuentran organizados longitudinalmente (Figura 7). Cálices dispuestos alrededor de todos los tallos de las ramas, separados entre sí. Se observan únicamente uno o dos pólipos por subdivisión de las ramificaciones. En promedio los cálices presentan 1.503 mm de altura y 0.875 mm de diámetro de área basal, siendo menor en el ápice de los pólipos: 0.773 mm. En los cálices cerrados se observan ocho lóbulos correspondientes a los ocho tentáculos.
Los escleritos antocodiales son principalmente escamas alargadas, también se encuentran algunas con formas irregulares (Figura 7), con longitud promedio de 0.15 mm y ancho mayor promedio de 0.03 mm; en algunos escleritos se observan ornamentaciones un poco espinosas y/o rugosas de altura promedio de 0.011 mm.
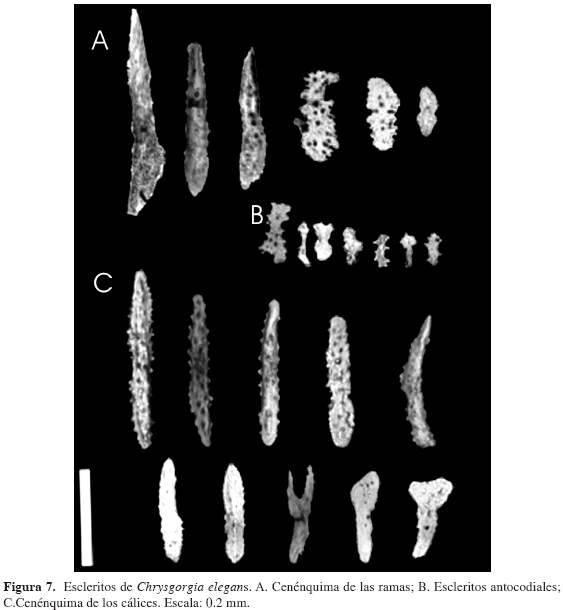
El color de las colonias preservadas en etanol al 70 y 96 % es entre café claro y crema.
Discusión: El material revisado se identificó como C. elegans. Sin embargo, presentaba algunos caracteres similares a C. agassizii (ambas especies presentan los siguientes caracteres en común: fórmula de ramificación, orientación de los escleritos en los cálices, forma de la colonia, coloración y el fragmento de fijación al sustrato son similares). Los especímenes colombianos presentan algunas similitudes con C. agassizii, tales como las distancias ("orthostiche interval" de 10-12 mm) e intervalos entre 10-12 mm ) e intervalos entre ramas (1.5-2 mm), el número de divisiones dicotómicas de las ramas (2 a 5), el diámetro promedio de las ramas terminales (0.1 mm). Por su parte, en C. elegans estos mismos caracteres presentan dimensiones diferentes, así las ramas están cada 1-1.5 mm, el "orthostiche interval" cada 7-7.5 mm, hay de una a tres divisiones dicotómicas y el diámetro promedio en las ramas es de 1.1 mm. Sin embargo, la longitud de las ramas y la dureza de las mismas, coinciden con las de C. elegans, la cual presenta ramas rígidas y quebradizas de 25 mm de longitud, mientras que C. agassizii presenta ramas flexibles y más largas que las observadas en el material colombiano con 65 mm de longitud. A pesar de las similitudes con ambas especies, fue posible identificar los especímenes colombianos como C. elegans, con base en lo mencionado anteriormente y además los tipos de escleritos presentes en las diferentes estructuras. Según Cairns (2001), una característica definitiva de C. agassizii es la presencia de escamas y ausencia de husos o bastones en los cálices, la cual es diferente en el material revisado donde sí se observan bastones y husos aplanados en los cálices (Figura 7). Un dato adicional que corrobora la identidad del material colombiano como perteneciente a C. elegans, son los registros realizados por Cairns (2001) con base en muestras recolectadas en la frontera entre Panamá y Colombia, La Guajira (531-537 m) y el Cabo San Juan de Guía en el Parque Tayrona (567-713 m). Adicional a todo lo anterior, esta especie se comparó con muestras provenientes del NMNH, siendo posible confirmar que la muestra revisada es C. elegans.
Distribución geográfica: Distribución discontinua en el norte del golfo de México a Florida, El "Little Bahama Bank", el Caribe suroriental y Antillas Menores hasta la frontera de Panamá y Colombia. En Colombia se distribuye en las ecorregiones Caribe Oceánico-COC frente a Archipiélagos Coralinos-ARCO, Palomino-PAL, Tayrona-TAY y Guajira-GUA.
Ámbito batimétrico: Chrysogorgia elegans se encuentra entre 128 y 1716 m de profundidad, siendo muy común entre 500 y 800 m (Cairns, 2001). En el área de estudio se encontró entre 484 y 516 m de profundidad.
Género Trichogorgia Hickson, 1904
Especie tipo: Trichogorgia flexilis Hickson, 1904
Diagnosis: Colonias ramificadas en un plano, flabeladas, con forma de lira, con ramas terminales largas. Base discoidal. Eje iridiscente, delgado y flexible en las ramas terminales, puede presentar o no depósitos calcáreos, cálices en dos filas a cada lado del eje, o alrededor de todas las ramas terminales y subterminales, dispersos o agrupados en las regiones apicales. Escleritos presentes o ausentes; si están presentes son en forma de discos dobles delgados o en forma de remos dobles con bordes finamente aserrados (Deichmann, 1936; Bayer y Muzik, 1976).
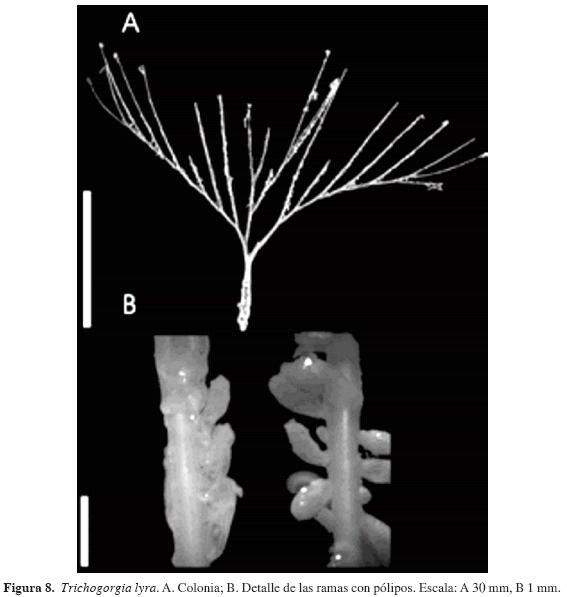
Trichogorgia lyra Bayer y Muzik, 1976
(Figura 8)
Material examinado: USNM 52868 (Paratipo, P370). USNM 52870 (Paratipo, P403). INV CNI1740 (12°29'18''N, 71°43'52''W). INV CNI1741 (11°17'32''N, 73°27'6''W). INV CNI1742 (12°29'18''N, 71°43'52''W). INV CNI1743 (11°23'83''N, 73°27'78''W). INV CNI1744 (11°19'86''N, 74° 5'28''W). INV CNI1745 (11°'18'28''N, 73°46'50''W). INV CNI1746 (11°5'46''N, 74°40''35'W). INV CNI1747 (11°18'29''N, 74°9'70''W). INV CNI1748 (11°18'3''N, 74°9'36''W). INV CNI1749 (11°5'58''N, 74°40'37''W). INV CNI1842. (12°29'18''N, 71°43'52''W). INV CNI1843 (11º23'83''N, 73°27'78''W). INV CNI1844 (11°23'83''N, 73°27'78''W). INV CNI1845 (11°24'4''N, 73°27'32''W). INV CNI1846 (11°17'41''N, 73°27'58''W). INV CNI1848 (12°29'93''N, 71°45'21''W). INV CNI1849 (11°21'29''N, 74°6'17''W).
Diagnosis: Colonias flabeladas en un solo plano; forma de lira; pólipos dispersos pero dispuestos en dos filas a lado y lado de las ramas, no se agrupan en los ápices. Eje con depósitos calcáreos. Ausencia absoluta de escleritos en toda la colonia. Base rizoidal (Deichmann, 1936; Bayer y Muzik, 1976).
Descripción: Colonias hasta de 10.5 cm de ancho por 11.5 cm de alto (Figura 8A). Eje principal con forma circular en sección transversal, diámetro promedio de 0.916 mm (0.5-1.45 mm), que disminuye hacia los dos extremos de la colonia. Diámetro promedio de las ramas de 0.539 mm (0.232-0.812 mm) siendo menor hacia los ápices.
El cenénquima, completamente desprovisto de escleritos, rodea el tallo principal con una capa gruesa de color café con tonalidad grisácea; las ramas presentan hendiduras longitudinales visibles, a manera de canales, desde la base de las ramas. De estos canales se originan los pólipos a lado y lado de las ramas dispuestos muy cerca entre sí, pero no contiguos. Los pólipos presentan forma de copa y un color café más oscuro que el resto de la colonia (Figura 8B).
Discusión: Trichogorgia lyra presenta características inconfundibles como: forma de las colonias, ausencia de escleritos, manera en que el cenénquima cubre los ejes de las ramas (formando canales) y modo en que se originan los pólipos a partir de dichos canales longitudinales.
Distribución geográfica: Yucatán, Honduras, Nicaragua, Panamá, y Colombia: Islas de San Bernardo, Punta Caribaná y entrada del golfo de Urabá (Bayer y Muzik, 1976). En el área de estudio se encontró además en las ecorregiones Guajira-GUA, Palomino-PAL, Tayrona-TAY, Magdalena-MAG y Darién-DAR.
Ámbito batimétrico: Trichogorgia lyra se encuentra entre 23 y 183 m de profundidad (Bayer y Muzik, 1976). Dentro del área de estudio se encontró entre 20 y 150 m de profundidad.
AGRADECIMIENTOS
Al Instituto de Investigaciones Marinas y Costeras "José Benito Vives De Andréis"-INVEMAR, y en particular al Museo de Historia Natural Marina de Colombia (MHNMC), por el apoyo permanente en cuanto a información, equipos y espacio adecuado para el desarrollo del proyecto dentro del cual se enmarca este trabajo. Al Dr. Stephen Cairns, por su colaboración y asesoría para la identificación del material y facilitar bibliografía, durante las diferentes pasantías realizadas en el National Museum of Natural History (Smithsonian Institution). A Patricia Lattig, por su ayuda con la identificación de algunas muestras. A los proyectos INVEMAR-Macrofauna (No. 2015-09-16649). A los editores y evaluadores por sus valiosos aportes.
BIBLIOGRAFÍA
1 Bayer, F. M. 1951. A new Caribbean coral of the genus Chrysogorgia. Proc. U. S. National Mus., 101 (3276): 269-273. [ Links ]
2 Bayer, F. M. 1956. Octocorallia. 166-231. En: Moore, R. C. (Ed.). Treatise on invertebrate paleontology. Part F. Coelenterata. Primera Edición. Geological Society of America and University of Kansas Press, Kansas, EE. UU. 508 p. [ Links ]
3 Bayer, F. M. 1961. The shallow water Octocorallia of the West Indian region. A manual for marine biologists. Smithsonian Institute. Washington, EE. UU. 373 p. [ Links ]
4 Bayer, F. M. 1981. Key to the genera of Octocorallia exclusive of Pennatulacea (Coelenterata: Anthozoa), with diagnoses of new taxa. Proc. Biol. Soc. Wash. 94 (3): 902-947. [ Links ]
5 Bayer, F. M. y K. Muzik. 1976. New genera and species of the holaxonian family Chrysogorgiidae (Octocorallia: Gorgonacea). Zool. Med. Leiden, 50 (5): 65-90. [ Links ]
6 Bayer, F. M. y J. Stefani. 1988. A new species of Chrysogorgia (Octocorallia: Gorgonacea) from New Caledonia, with descriptions of some other species from the Western Pacific. Proc. Biol. Soc. Wash. 101 (2): 257-279. [ Links ]
7 Botero, L. 1987. Zonación de octocorales gorgonáceos en el área de Santa Marta y Parque Nacional Natural Tayrona, Costa Caribe colombiana. An. Inst. Invest. Mar. Punta de Betín, 17: 61-80. [ Links ]
8 Cairns, S. D. 2001. Studies on western Atlantic Octocorallia (Coelenterata: Anthozoa) Part 1: The genus Chrysogorgia Duchassaing and Michelotti, 1864. Proc. Biol. Soc. Wash., 114 (3): 746-787. [ Links ]
9 CIOH. 1991. Carta de navegación No. COL 407. Santa Marta a Puerto Colombia. Escala 1:100.000.10º35'- 11º5' N: 75º40'-74º50' W. Centro de Investigaciones Oceanográficas e Hidrográficas, Cartagena. [ Links ]
10 Defense Mapping Agency Hydrografic/Topographic Center. 1990. Carta de navegación No. 24480. Cabo Tiburón to Barranquilla. Scale 1:300.000; 7º50'-9º50' N; 76º50'-74º50' W. Washington D. C. [ Links ]
11 Defense Mapping Agency Hydrografic/Topographic Center. 1995a. Carta de navegación No. 24490. Río Magdalena to Cabo de la Vela. Escala 1:300.000;10º50'-12º40 N; 72º-75º W. Washington D.C. [ Links ]
12 Defense Mapping Agency Hydrografic/Topographic Center. 1995b. Carta de navegación No. 24501. Río Magdalena to Punta Canoas. Escala 1:80.000;10º35'-11º5 N; 75º40-74º50 W. Washington D.C. [ Links ]
13 Deichmann, E. 1936. The Alcyonaria of the western part of the Atlantic Ocean. Memoirs of the Museum of Comparative Zoology at Harvard College, Vol. LIII. Cambridge Mass, EE. UU. 315 p. [ Links ]
14 Díaz, J. M. y D. I. Gómez. 2000. Programa nacional de investigaciones en biodiversidad marina. INVEMAR y Ministerio del Medio Ambiente, Bogotá, D. C. 83 p. [ Links ]
15 Krieger, K. 2001. Coral (Primnoa) impacted by fishing gear in the Gulf of Alaska. 106-116. En: Proceedings of the First International Symposium on Deep-Sea Corals. Ecology Action Center. Halifax, Nueva Escocia. 231p. [ Links ]
16 Lattig-Matiz, P. 2000. Porifera, Cnidaria y Annelida de la franja superior del talud continental (200-500 m) del Caribe colombiano. Trabajo de grado Biol. Pontificia Univ. Javeriana. Bogotá, D.C. 191 p. [ Links ]
17 Navas, G., L. S. Mejía-Mantilla, N. Ardila, J. Reyes-Forero, A. Gracia, N. Santodomingo, A. Bermúdez- Tobón, M. Benavides-Serrato, G. Borrero-Pérez, N. Cruz-Casta y L. M. Mejía-Ladino. 2002. Fauna marina colombiana de la plataforma continental del mar Caribe colombiano, estado del conocimiento de la macrofauna de la plataforma continental del océano Pacífico colombiano y transferencia tecnológica y de información al SINA. Informe técnico final, UAESPNN, CORPOGUAJIRA y CVC, Santa Marta. [ Links ]
18 Pérez-Gil, R. M., A. D. Ávila-Cabrera, J. L. Bello-Gárciga, M. Cruz-Nurque, M. Calderón-Marín y M. Montalvo-Duquesne. 1999. Efecto producido sobre la actividad antitumoral del coralán por el tratamiento con una enzima proteolítica. Rev. Cub. Oncol., 15 (1): 30-35. [ Links ]
19 Reyes, J., N. Santodomingo, A. Gracia, G. Borrero-Pérez, G. Navas, L. M. Mejía-Ladino, A. Bermúdez y M. Benavides. 2005. Southern Caribbean azooxanthellate coral communities off Colombia. 309- 330. En: Freiwald, A y J. M. Roberts (Ed.). Cold-water corals and ecosystems. Springer-Verlag. Berlin. 1243 p. [ Links ]
20 Sánchez, J. A. 1994. Presencia de los octocorales Stylatula diadema Bayer (Pennatulacea) y Carijoa riisei (Duchassaing y Michelotti)/Telestacea) en la costa Caribe colombiana. An. Inst. Invest. Mar. Punta de Betín, 23: 137-147. [ Links ]
21 Sánchez, J. A. 1998. Sistemática filogenética del género Eunicea Lamoroux, 1816 (Octocorallia: Gorgonacea: Plexauridae) con aspectos sobre la historia natural de algunas especies en el Caribe colombiano. Tesis de Maestría, Univ. Nacional de Colombia, Bogotá. 134 p. [ Links ]
22 Sánchez, J. A. 1999. Black coral-octocoral distribution patterns on a deep-water reef, Imelda bank, Caribbean sea, Colombia. Bull. Mar. Sci. 65 (1): 215-225. [ Links ]
23 Sánchez, J. A., S. Zea y J. M. Díaz. 1998. Patterns of octocoral and black coral distribution in the oceanic barrier reef-complex of Providencia Island, Southwestern. Carib. J. Sci. 34 (3-4): 250-264. [ Links ]
24 Williams, G. C. 1992. The Alcyonacea of Southern Africa. Stoloniferous octocorals and soft corals (Coelenterata, Anthozoa). Ann. S. Afr. Mus., 100 (3): 249-358. [ Links ]
25 Williams, G. C. 2001. Systematic list of valid octocoral genera. http://www.calacademy.org/research/izg/OCTOCLASS.htm#alcyo. 10/10/10. [ Links ]
26 Witherell, D. y C. Coon. 2001. Protecting gorgonian corals off Alaska from fishing impacts. Report to NPFMC. 117-125. En: Willison, J. H. M., J. Hall, S. E. Gass, E. L. R. Kenchington, M. Butler y P. Doherty (Eds) Proceedings of the First International Symposium on Deep-Sea Corals. Ecology Action Centre, Nova Scotia Museum, Halifax, Nueva Escocia. 231 p. [ Links ]
FECHA DE RECEPCIÓN: 23/12/2008 FECHA DE ACEPTACIÓN: 14/10/2010
*Contribución No. 1064 del Instituto de Investigaciones Marinas y Costeras-INVEMAR.
