










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.39 no.2 Santa Marta July/Dec. 2010
NOTA:
LA ALIMENTACIÓN DE TIBURONES MARTILLO JOVENES (SPHYRNA LEWINI) CAPTURADOS EN EL GOLFO DE NICOYA, COSTA RICA
THE FEEDING HABITS OF JUVENILE SCALLOPED HAMMERHEAD SHARKS (SPHYRNA LEWINI) CAUGHT IN THE GULF OF NICOYA, COSTA RICA
Ilena Zanella Cesarotto1, Andrés López Garro2 y Randall Arauz Vargas2
1 Universidad Nacional de Costa Rica, Instituto Internacional en Conservación y Manejo de Vida Silvestre. Apartado 1350-3000 Heredia, Costa Rica. izanella@misiontiburon.org
2 Programa Restauración de Tortugas Marinas (PRETOMA). Apartado 1203-1100 San José, Costa Rica. alopez@misiontiburon.org (A.L.G.); rarauz@pretoma.org (R.A.V.)
ABSTRACT
Sharks present strong intraspecific changes in feeding habits, which may vary according to size, geographic location and season. This study analyzed the stomach contents of 52 juvenile scalloped hammerhead sharks (25 females and 27 males) caught by artisanal vessels in the Gulf of Nicoya. Regarding the organisms found in the stomachs, 41.07 % were teleost fish, 30.36 % mollusks and 28.57 % crustaceans. Mollusks presented the highest Relative Importance Index (RII) (43.05 %), followed by teleost fish (41.37 %) and crustaceans (15.58 %). Squids (Loliginidae) were the items with the greatest RII (32.57 %). By means of the Pianka index, a relatively low overlap between sexes was identified, suggesting a spatial sexual segregation and, in consequence, an ecological niche separation between male and female hammerhead sharks.
KEY WORDS: Scalloped hammerhead shark, Stomach contents, Gulf of Nicoya, Relative importance index.
Los tiburones son los máximos depredadores de los ambientes acuáticos y por ende realizan un papel primordial en el paso de la energía entre los niveles tróficos superiores de la cadena alimentaria marina (Wetherbee y Cortés, 2004). A pesar de esto, existe un vacío de información sobre su biología alimentaria, debido principalmente a las dificultades del muestreo (preservación y transporte de muestras obtenidas en alta mar) y a los fuertes cambios intraespecíficos en la dieta, según la talla de los individuos, la localización geográfica y la época del año. Los cambios ontogénicos resultan evidentes para especies de tiburones pelágicas y migratorias, que conforme van creciendo, cambian de hábitat y el aumento del tamaño de sus mandíbulas les permite alimentarse de presas cada vez más grandes (Wetherbee y Cortés, 2004). Entre estas especies, se encuentra el tiburón martillo Sphyrna lewini, que se alimenta principalmente de peces e invertebrados (moluscos y crustáceos, entre otros) (Klimley, 1987; Anislado, 2000; Torres, 2004). En Costa Rica no se han realizados estudios acerca de la biología alimentaria de esta especie por lo cual el presente estudio analiza la dieta de jóvenes de tiburón martillo (S. lewini) en el golfo de Nicoya, Costa Rica.
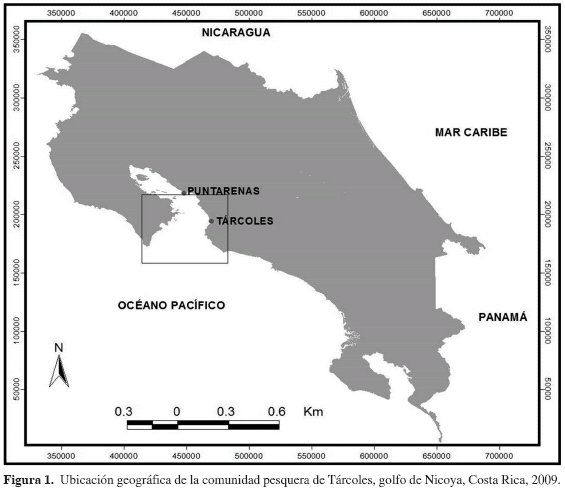
Los muestreos se realizaron entre febrero de 2006 y marzo de 2007, en la comunidad pesquera de Tárcoles, en el Pacífico central de Costa Rica (Figura 1). Dicha localidad está ubicada a 37 Km al sureste de la ciudad de Puntarenas, en el extremo oriental de la desembocadura del golfo de Nicoya. Los pescadores de Tárcoles, organizados en la Cooperativa de Pescadores Artesanales de Tárcoles (CoopeTárcoles R. L.), utilizan embarcaciones con una autonomía para faenar un máximo de cinco millas náuticas, suelen operar en la parte externa del golfo de Nicoya, y por lo general utilizan línea de fondo o planera y trasmallos (redes de enmalle). Durante los desembarques pesqueros, se determinó la longitud total (LT), como la distancia entre la punta del hocico del tiburón hasta el extremo distal de la aleta caudal, el peso total (PT) y el sexo, mediante la observación de la presencia o ausencia de mixopeterigios o cláspers de los tiburones martillo descargados. A continuación se realizó una incisión en la región ventral y se extrajo el estómago a cada tiburón, el cual luego fue almacenado en una bolsa plástica con formalina al 10 % por un período de 2 a 8 meses. Todos los estómagos fueron trasladados al Laboratorio de Recursos Naturales y Vida Silvestre (LARNAVISI) de la Escuela de Ciencias Biológicas de la Universidad Nacional de Costa Rica, donde fueron diseccionados y analizados. Los contenidos estomacales se lavaron y se clasificaron hasta la categoría taxonómica más baja posible y, cuando fue necesario, se utilizó un estereoscopio para su debida clasificación. La ictiofauna se clasificó mediante características anatómicas externas (Bussing y López, 1993; Fischer et al., 1995), mientras que los crustáceos y los moluscos se identificaron según la guía de Fischer et al. (1995). Se excluyó del análisis cualquier vestigio de carnada dentro de los estómagos. Para cada taxón identificado como alimento, se calculó la frecuencia de presencia (% F), la proporción respecto al número (% N), y la proporción en peso (% W) (Wetherbee y Cortés, 2004), utilizando las siguientes fórmulas:
% F = (número de estómagos con una presa específica/total de estómagos analizados)*100
% N = (número total de una presa específica/número total de las presas)*100
% W = (peso total de una presa/peso total de todas las presas)*100
Se calculó el índice de importancia relativa (% IRI): IRI = (% N + % W) x % F para determinar la presa más significativa para el tiburón martillo. Además se utilizó el índice de Pianka (O) (Krebs, 1986) para determinar el grado de solapamiento del nicho trófico entre los ítems alimentarios de machos y hembras.
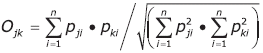
Donde Pji y Pki son las proporciones de un recurso i, utilizado por dos grupos, en este caso por machos y hembras. El índice oscila entre 0 y 1; cuando el valor supera 0.60 el solapamiento se considera significativo (Zares y Rand, 1971). Como este índice no provee una medida de dispersión del cálculo, se realizó un análisis de remuestreo de los datos mediante las técnicas de randomización de Jackknife (InfoStat/L).
Durante el período de muestreo se analizaron los contenidos estomacales de 52 tiburones martillo, de los cuales 25 fueron hembras y 27 machos. En comparación con otros estudios sobre alimentación de tiburones martillo, la presente investigación tuvo acceso a un número menor de estómagos (Anislado, 2000; Aguilar, 2003; Torres, 2004). Según Torres (2004), el número mínimo de estómagos necesarios para caracterizar las presas de S. lewini, aplicando el índice de Hoffman, es 47. Por lo tanto, la información brindada en este estudio puede ser utilizada como referencia para otras investigaciones sobre la alimentación de S. lewini en su etapa juvenil.
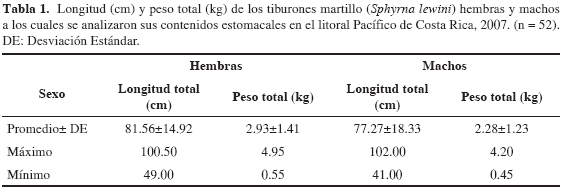
La longitud y peso total promedio de los tiburones hembra analizados fue de 81.56 cm (± 14.92 de desviación estándar) y 2.93 kg (± 1.41) respectivamente. Para el caso de los machos fue de 77.27 cm (± 18.33) y 2.28 kg (± 1.23), respectivamente (Tabla 1). Según Anislado (2000), en el litoral de Michoacán de México las hembras alcanzan la madurez sexual a los 190 cm y a una edad de 4.4 años, mientras que los machos alcanzan la madurez sexual a los 170 cm y a una edad de 3.6 años. De esta forma se confirma que los individuos analizados en este estudio fueron todos tiburones jóvenes, por lo tanto, los resultados deben ser referidos exclusivamente a este estado de desarrollo. Según Klimley (1987), Anislado (2000) y Aguilar (2003) existen fuertes diferencias en la dieta de tiburones martillo de distintas tallas y edades, así comprobando cambios alimentarios ontogénicos. Dichas diferencias son consecuencia de las migraciones horizontales y verticales que realizan los tiburones martillo, tanto los jóvenes en aguas costeras como los adultos en aguas oceánicas (Klimley et al., 1993). De los 52 estómagos analizados ocho estaban vacíos. La presencia de estómagos vacíos ha sido registrada también por otras investigaciones sobre biología alimentaria de tiburones (Wetherbee y Gruber, 1990; Lowe et al., 1996; Wetherbee y Cortés, 2004). Algunos autores relacionan este aspecto con regurgitaciones provocadas por fuertes cambios de presión durante el levantamiento de la línea de pesca (Wetherbee et al., 1990) mientras que Wetherbee y Cortés (2004) sugieren que algunos tiburones se alimentan en forma intermitente por lo cual existen períodos donde los tiburones no tendrían alimento en su cavidad estomacal.
De los organismos encontrados en los estómagos de los tiburones, 41.07 % fueron teleósteos, 30.36 % moluscos y 28.57 % crustáceos (Tabla 2). Los moluscos tuvieron el mayor índice de importancia relativa (43.05 %), seguidos por los teleósteos (41.37 %) y por último los crustáceos (15.58 %). La importancia de los moluscos se concentró en los representantes de la familia Loliginidae, la cual aportó el mayor % IRI del total (32.57 %) (Tabla 2). De acuerdo con Anislado (2000), Torres (2004) y Aguilar (2003), entre las principales presas de los jóvenes de tiburón martillo figuran los teleósteos, los moluscos (en particular los cefalópodos) y los crustáceos. Al igual que lo registrado por Torres (2004), en el presente estudio los teleósteos fueron el taxón más importante en cuanto a frecuencia de presencia. Sin embargo, los moluscos fueron los que aportaron mayor peso y mayor índice de importancia relativa. Dentro de este grupo, sobresalió la familia Loliginidae, representada por especies de calamares demersales de talla pequeña, que generalmente se encuentran en aguas costeras y someras (Fischer et al., 1995). Dichas aguas son también las favoritas por los jóvenes de tiburón martillo, los cuales debido a su temprana edad, poseen poca destreza y capacidad en cuanto a búsqueda de alimento (Aguilar, 2003). Como consecuencia, canalizan su energía en la captura de especies bentónicas y demersales de talla pequeña (Aguilar, 2003; Torres, 2004), siendo los loligínidos presas ideales. De la misma forma Aguilar (2003), quién analizó la ecología trófica del tiburón martillo en el golfo de California, determinó que los cefalópodos tuvieron el mayor índice de importancia relativa, siendo los jóvenes del calamar gigante (Dosidicus gigas) la categoría más importante en la dieta. Smale y Cliff (1998), quienes también realizaron un estudio sobre la dieta de S. lewini en Sudáfrica, indicaron que sus presas favoritas eran los cefalópodos, entre las cuales figuraban especies de la familia Loliginidae.
En cuanto al grado de solapamiento entre la dieta de machos y hembras, el valor del índice de Pianka promedio para este estudio fue de 0.38, con una desviación estándar de ±0.1. Por lo tanto, se determinó un solapamiento entre sexos relativamente bajo, lo cual sugiere que existe una separación de nicho ecológico entre machos y hembras del tiburón martillo. Klimley (1987) describió que las hembras y machos de tiburón martillo en el golfo de California se alimentan de presas diferentes debido a la segregación espacial entre los dos sexos. En contraste, Aguilar (2003) documentó un traslape significativo entre machos y hembras en el golfo de California, sugiriendo que se alimentan de presas similares en proporciones semejantes, lo que sugiere que coexisten temporal y espacialmente en una misma área. Por lo tanto, el bajo grado de solapamiento entre sexos registrado por el estudio, sugiere la presencia de segregación espacial como la descrita por Klimley (1987) y Klimley et al. (1993); sin embargo, solamente la realización de estudios futuros podrán afirmar o no esta tendencia.
AGRADECIMIENTOS
Se agradece el apoyo financiero de Conservación Internacional y Whitley Found for Nature, así como la valiosa ayuda brindada por la Cooperativa de Pescadores Artesanales de Tárcoles R.L. (CoopeTárcoles R.L.) y por los voluntarios Loic Le Foulgoc y Benoit Figarede de la Universidad La Rochelle de Francia. Además, agradecemos a Joel Sáenz del Instituto Internacional en Conservación y Manejo de Vida Silvestre; a Julio Lamilla de la Universidad Austral de Chile; a Maurizio Protti de la Escuela de Biología de la Universidad Nacional de Costa Rica y a José Rodrigo Rojas del Instituto Costarricense de Electricidad.
BIBLIOGRAFÍA
1 Aguilar, C. A. 2003. Ecología trófica de juveniles del tiburón martillo Sphyrna lewini (Griffith and Smith, 1834) en el golfo de California. Tesis Maestría Cien., Cicimar, La Paz, México. 119 p. [ Links ]
2 Anislado, T. V. 2000. Ecología pesquera del tiburón martillo, en el litoral del estado de Michoacán, México. Tesis Maestría Cien., UNA, México D. F. 145 p. [ Links ]
3 Bussing, W. y M. López. 1993. Peces demersales y pelágicos costeros del Pacífico de Centro América Meridional. Rev. Biol Trop. (edición especial), 164 p. [ Links ]
4 Fischer, W., F. Kupp, W. Schneider, C. Sommer, K. E. Carpenter y V. H. Niem. 1995. Guía FAO para la identificación de especies para los fines de la pesca. Pacífico Centro-Oriental. Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO), Roma. 1813 p. [ Links ]
5 Klimley, A. P. 1987. The determinants of sexual segregation in the scalloped hammerhead shark, Sphyrna lewini. Environ Biol Fish, 18 (1): 27-40. [ Links ]
6 Klimley, A. P., M. I. Cabrera y G. Castillo. 1993. Descripción de los movimientos horizontales y verticales del tiburón martillo, Sphyrna lewini, del sur del golfo de California de México. Ciencias Marinas, 19 (1): 95-115. [ Links ]
7 Krebs, C. J. 1986. Ecología: análisis experimental de la distribución y abundancia. Edición 3. Ediciones Pirámides, Madrid. 782 p. [ Links ]
8 Lowe, C. G., B. M. Wetherbee, G. L. Crow y A. L. Tester. 1996. Ontogenetic dietary shifts and feeding behavior of the tiger shark, Galeocerdo cuvier, in Hawaiian waters. Environ Biol Fish, 47: 203-211. [ Links ]
9 Smale, M. J. y G. Cliff. 1998. Cephalopods in the diets of four shark species (Galeocerdo cuvier, Sphyrna lewini, S. zygaena and S. mokarran) from Kwazulu-Natal, South Africa. S. Afr. J. Mar. Sci, 20: 241-253. [ Links ]
10 Torres, H. A. M. 2004. Distribución, abundancia y hábitos alimentarios de juveniles del tiburón martillo Sphyrna lewini Griffith y Smith (SPHYRNIDAE) en la costa de Sinaloa, México durante el evento El Niño 1997-98. Tesis Maestría Cienc., Univ. Mar, Oaxaca, México. 91 p. [ Links ]
11 Wetherbee, B. M. y E. Cortés. 2004. Food consumption and feeding habits. 225-246. En: Carrier, J. C., J. A. Musick y M. R. Heithaus (Eds.). Biology of sharks and their relatives. CRC Press. Boca Ratón, EE. UU. 596 p. [ Links ]
12 Wetherbee, B. M. y S. H. Gruber. 1990. The effect of ration level on food retention time in juvenile lemon sharks, Negaprion brevirostris. Environ Biol Fish, 29: 59-65. [ Links ]
13 Wetherbee, B. M., S. H. Gruber y E. Cortés. 1990. Diet feeding habits, digestion, and consumption in sharks, with special reference to the lemon shark, Negaprion brevirostris. 29-47. En: Pradtt, H. L., S. H. Gruber y T. Taniuchi (Eds.). Elasmobranchs as living resources: advances in the biology, ecology, systematics, and the status of the fisheries. Reporte técnico, NOAA-NMFS. 90 p. [ Links ]
14 Zares, T. M. y A. S. Rand. 1971. Competition in tropical stream fishes: support for the competitive exclusion principle. Ecology, 52: 336-342. [ Links ]
FECHA DE RECEPCIÓN: 15/03/2010 FECHA DE ACEPTACIÓN: 28/06/2010
