










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.40 supl.1 Santa Marta Dec. 2011
VARIATION IN THE SURFACE CURRENTS IN THE PANAMA BIGHT DURING EL NIÑO AND LA NIÑA EVENTS FROM 1993 TO 2007
VARIACIÓN EN LAS CORRIENTES SUPERFICIALES EN EL PANAMA BIGHT DURANTE EVENTOS EL NIÑO Y LA NIÑA OCURRIDOS ENTRE 1993 Y 2007
Andrea Corredor-Acosta1, Alberto Acosta1, Phillipe Gaspar2 and Beatriz Calmettes2
1 Pontificia Universidad Javeriana, Facultad de Ciencias, Departamento de Biología, Unidad de Ecología y Sistemática (UNESIS). Carrera 7 No 40-62, Bogotá, Colombia. andreaacosta28@hotmail.com (A.C.A.); laacosta@javeriana.edu.co (A.A.).
2 Collecte Localisation Satellites (CLS), Direction Océanographie Spatiale, 8-10 rue Hermes, 31520 Ramonville, France. philippe.gaspar@cls.fr (P.G.); bcalmettes@cls.fr (B.C.).
ABSTRACT
Climatic anomalies have changed the ocean circulation pattern and thus the demographic connectivity. However, in many geographical regions there is insufficient evidence of this change. Therefore, comparisons were made between neutral years and years of El Niño and La Niña with moderate intensity, for the North Equatorial Counter Current (NECC), the South Equatorial Current (SEC), the Coastal Current (CoaC) and the main anticyclonic eddy in the Panama Bight. Daily dynamics topography data of the Maps of Absolute Dynamic Topography (MADT) provided by AVISO and daily wind stress data provide by the European Centre for Medium Range Weather (ECMWF) were used to calculate the speed of surface currents (multi-year, quarterly average), during months with the highest number of eggs and larvae released by the species with a pelagic phase (Sept-Nov). It was found that the speed magnitude for the three oceanic currents was statistically different among the compared events, except for the anticyclonic eddy; obtaining higher values of speed for neutral years in relation to years with El Niño or La Niña for the NECC, for the SEC higher values for La Niña years, followed by neutral years and a moderate El Niño years; for the CoaC higher velocity for neutral and La Niña years but the lowest for El Niño years; and a tendency of higher values in La Niña years for the anticyclonic eddy. Additionally, the number of eddies increased in moderate El Niño years. The results suggest that the decreased velocity of the NECC and the potential barriers created by the cyclonic eddies and the anticyclonic eddy near the South American coast could diminish the passive dispersal of larvae and the potential functional connectivity between the Western, Central and Eastern Tropical Pacific. Therefore, there are implications at the evolutionary, biogeographic, and ecologic levels (dispersion rates and population rescue effect). In contrast, during La Niña the SEC could favor teleplanktonic larval transport to the Central Pacific, material which is exported from the South American coast by CoaC, aided by the anticyclonic eddy. In conclusion, anomalous climatic events alter the velocity of oceanic currents in the Panama Bight; consequently these could change the functional potential connectivity from September to November.
KEY WORDS: Ocean Currents, ENSO, Climatic Anomalies, Panama Bight.
RESUMEN
Las anomalías climáticas modifican el patrón de circulación oceánica y con ello la conectividad demográfica. Sin embargo, en muchas regiones geográficas no hay suficiente evidencia de este cambio. Por ello, en el Panama Bight se realizó la comparación entre años neutrales, años El Niño y La Niña de moderada intensidad, para la Contracorriente Norecuatorial (CCNE), la Corriente Surecuatorial (CSE), la Corriente Costera (CCos) y para el remolino anticiclónico principal. Datos diarios de la topografía dinámica proporcionados por AVISO usando el producto MADT y del estrés del viento proporcionados por el Centro Europeo de Meteorología a Medio Plazo (ECMWF) se usaron para calcular la velocidad de las corrientes superficiales (promedio trimestral multianual) para los meses con mayor cantidad de huevos y larvas liberados por las especies con fase pelágica (sep-nov). Se encontró que la magnitud de la velocidad para las tres corrientes oceánicas fue estadísticamente diferente entre los distintos eventos comparados, excepto para el anticiclón. Obteniendo que los valores de velocidad fueron mayores en años neutrales en relación a años Niño y Niña para la CCNE; mayores en años Niña, seguido por neutrales y Niño para la CSE; mayor en años neutrales y Niña pero menor para años Niño en la CCos; y tendencia de mayores valores en años La Niña para el remolino anticiclónico. Adicionalmente, se observó un aumento en el número de remolinos en años Niño moderado. Los resultados sugieren que la disminución en la velocidad de circulación de la CCNE y las posibles barreras creadas por remolinos ciclónicos y el remolino anticiclónico presentes cerca a la costa Suramericana podrían disminuir la dispersión pasiva de larvas y la conectividad funcional potencial entre el Pacífico Occidental, Central y Tropical Oriental, lo cual tiene implicaciones a nivel evolutivo, biogeográfico y ecológico (tasa de dispersión y efecto de rescate poblacional). Contrariamente, durante La Niña la CSE podría favorecer el transporte de larvas teleplantónicas hacia el Pacífico Central, material exportado desde la costa Suramericana mediante la CCos, ayudado por el remolino anticiclónico. Se concluye que los eventos climáticos anómalos alteran la velocidad de las corrientes oceánicas en el Panama Bight, lo cual podría afectar la conectividad funcional potencial entre septiembre y noviembre.
PALABRAS CLAVE: Corrientes Oceánicas, ENOS, Anomalías Climáticas, Panama Bight.
INTRODUCTION
There are widespread concerns about regional marine biodiversity loss as a consequence of human overexploitation and global change. Management strategies such as Marine Protected Areas (MPAs) and conservation corridors (e.g. Eastern Tropical Pacific marine corridor), has been touted as a method for both conserving biodiversity and managing fisheries (Cowen and Sponaugle, 2009). However, there is a need for models that capture the spatial and temporal dynamics of patchy (or isolated) marine populations and its viability, especially with respect to biophysical processes such as larval dispersal (movement away from a spawning location) and passive transport due to oceanic currents (Sale et al., 2010). Determining how the oceanic current, which changes seasonally and annually, affects the patterns of planktonic larval dispersal is important to understand demographic (the exchange of individuals among local populations, that can influence population demographics and dynamics), and evolutionary connectivity (the amount of gene flow occurring among populations, which determine the extent of genetic differences among populations) as well as its relevance for the design and management of marine reserve networks and conservation (Sale et al., 2010).
It is common knowledge that marine currents are one of the mechanisms which promote transport of gametes, larvae, propagules or individuals among islands or between them and the continent (DiBacco et al., 2006). Hence, the viability and evolution of several marine populations or sub-populations depends on the gamete-larvae subsidy from distant populations (Hanski and Simberloff, 1997). The subsidy of downstream or sink populations (consistently receives more immigrants than it sends emigrants) is determined by the direction and velocity of the currents passively transporting the larvae, which for the most part have a limited time of development in the water column (Cowen et al., 2006). Thus, there is no certainty in which spatial scale (tens, hundreds, or thousands of kilometers) the connectivity is possible, given that in practice it is impossible to actually monitor the mobility of this reproductive material in the ocean currents and the success of the settlement (dispersal kernel), due to the lack of technical tools (Crooks and Sanjayan, 2006; Cowen and Sponaugle, 2009). Thereby, at least from the theoretical point of view, understanding the circulation of the ocean surface in terms of direction and velocity is an indirect mechanism employed to infer the potential functional connectivity in different regions (organism's behavioral response to landscape elements, e.g. patches, barriers), caused by the dispersion and response of marine organisms (Hanski and Simberloff, 1997; Werner et al., 2007). Data of larvae distribution suggest that the Eastern Tropical Pacific (ETP) could act as a barrier for the dispersion of several larval forms of invertebrates between the continental coast and the Central Pacific (Scheltema, 1988).
Knowledge pertaining to currents in the ETP (30° N-20° S; 75°-140° W) included studies from Wooster (1959) and Wyrtki (1961), these studies compiled and defined oceanography and the ocean circulation patterns (Figure 1a), then Wyrtki (1967) discussed the cyclonic/anticyclonic eddy present in the Panama Bight, and Fiedler (2002) examined the monthly climatic effect on the structure and evolution of the Costa Rican Dome (uniquely productive habitat), among others. Recently, Kessler (2006) summarized the patterns of oceanic circulation and described the main currents in the ETP: the California Current (CC), the North Equatorial Current (NEC), the South Equatorial Current (SEC), which flows towards the West; the North Equatorial Counter Current (NECC) which flows East; the Humboldt Current (HC) and the Coastal Current of Costa Rica (CCCR), which runs parallel to the coast of Peru/Chile and Costa Rica, respectively.
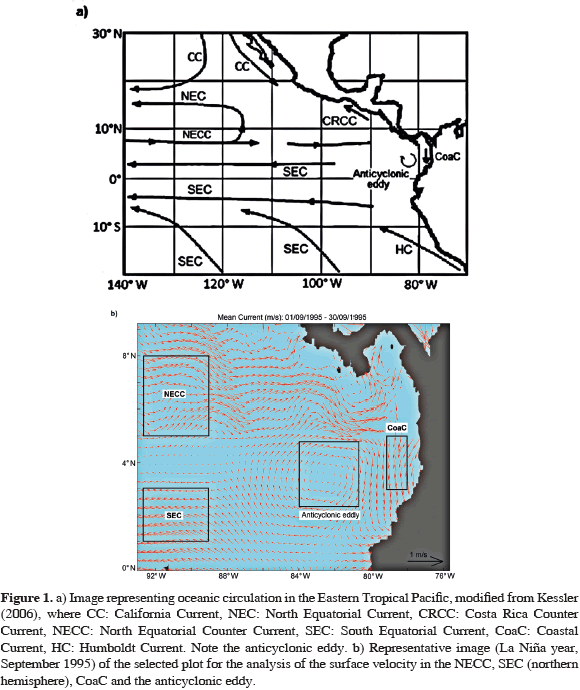
The eco-region of the Panama Bight (9° N-2° S; 75°-94° W) within the ETP Province, has three main oceanic currents the NECC, SEC (Glynn and Ault, 2000; Lukas, 2001; Chaigneau et al., 2006; Kessler, 2006) and the Coastal Current (CoaC; Chaigneau et al., 2006; Devis-Morales et al., 2008) with general velocities nearing ~22-32 cm/s. The NECC characteristically flows towards the East and ends near the coast of Panama, it flows in opposite direction to the trade winds and is confined within 4°N and 10°N (Glynn and Ault, 2000; Kessler, 2002; Rodríguez-Rubio et al., 2003, 2007; Rojas, 2005; Kessler, 2006; D'Croz and O'Dea, 2007; Devis-Morales et al., 2008). Lukas (2001) and Kessler (2006) have described the SEC as an equatorial current flowing in western direction between 3° N and 20° S, even though the SEC is divided in the southern hemisphere by the South Equatorial Counter Current (SECC). The SEC presents a greater velocity to the North than to the South of the equator as well as to the East than to the West; and is generated by the advection of the NECC, the equatorial upwelling, and the deviation towards the West of the Peruvian Current.
The circulation in the Panama Bight indicates a seasonal variation influenced by Trade winds (Devis-Morales et al., 2008). Therefore, in winter in the northern hemisphere (January-March), when the Intertropical Convergence Zone (ITCZ; Tomczak and Godfrey, 1994) moves South (~1° N), it generates an area of low atmospheric pressure, which together with the influence of the Panama Jet and the effect of wind stress produces a cyclonic eddy which dominates the Coastal Current (CoaC) flow from South to North, parallel to the South American coast (>50 km). The opposite event occurs in summer (July-September) when the ITCZ moves North (~10° N), the southeast trade winds are strengthened creating an area of high atmospheric pressure, and the influence of the Choco Jet is perceived (Poveda and Mesa, 2000) generating an anticyclonic eddy (mean velocities of ~40 cm/s) that dominate the Coastal Current flow from North to South along the South American coast (Rodríguez-Rubio et al., 2003; Devis-Morales et al., 2008). The Coastal Current is formed by the mixture of the Colombian (mass of low-salinity water due to intense precipitation and river discharge) and Panama Currents (Rodríguez-Rubio et al., 2003).
The Panama Bight is part of the Eastern Tropical Pacific marine corridor (CMAR, 2004) in which Ecuador (Galapagos), Colombia (Malpelo and Gorgona), Panama (Coiba), and Costa Rica (Cocos) implemented regional strategies for the preservation of marine biodiversity. This agreement recognizes that the viability of populations and preservation of marine biodiversity depend on physicaloceanographic processes occurring at great spatial (Western, Central, and Eastern Pacific Basin) and temporal scales (climate change: inter-annual anomalies such as events like El Niño and La Niña) and consequently, the local and regional management initiatives will not be effective if the adequate climatic and oceanographic scales are not considered (Roberts, 1997; Palumbi, 2004). Because of the latter, it is important to recognize the pattern of velocity of the main currents in the Panama Bight and understand how these could be affected by climatic events.
According to Lavin et al. (2006), the lack of evidence regarding the multiannual variability and climate change in the Eastern Tropical Pacific region (~12x106 km2, which includes the Panama Bight) may be due to the lack of attention to this temporal scale. According to Trenberth (1997), El Niño South Oscillation (ENSO; Grodsky and Carton, 2001) occurs 31 % of the time, La Niña 23 % of the time and the remaining 56 % the circulation takes place under neutral conditions; this generates a large dynamic in the hydrology of the Pacific Basin.
Chaigneau et al. (2006) and Kessler (2006) stated that the ETP is very dynamic because of the intense interaction between the ocean and the atmosphere (jets and local winds), particularly under the influence of interannual events such as El Niño (being the biggest climate cycle). The nature of the process has been broadly discussed and has been associated with the extreme alteration of the temperature field (Philander, 1983; Trenberth and Hoar, 1996). It is known that El Niño generates intense climatic changes and effects in the oceanic conditions along the Tropical Pacific, where these anomalies originate, extending even to the Antarctic (Turner, 2004). El Niño affects the biogeochemistry including the enrichment of nutrients, which supports the fishing industry and the economy in different countries. It also has effects in the flow of CO2 between the ocean and the atmosphere (Chávez et al., 1999). According to Willet (1996), the number of anticyclonic eddies increased in number during El Niño years; as an example of this, González-Silvera et al. (2004) have identified up to 18 cyclonic and anticyclonic eddies near the coastal region of the ETP.
Climate change models predict the accelerated mass extinction of local species, even when there are no mechanisms to explain it from an oceanographic point of view, for example, changes in the pattern of currents and their effect on dispersal pathways (Glynn and Ault, 2000; Reyes-Bonilla et al., 2002). Likewise, the models predict that in the following decades El Niño will be of moderate intensity and will occur more frequently (Thompson et al., 2002).
For the Tropical Pacific, it has been indicated that the currents change velocity in the presence of climatic events like El Niño or La Niña (Glynn and Ault, 2000; Grodsky and Carton, 2001; Johnson et al., 2002). In the Panama Bight it has been statistically corroborated that there is a seasonal change in the velocity of the main currents in response to local winds (Rodríguez-Rubio et al., 2003) or in the presence or absence of climatic events (La Niña/El Niño; Chaigneau et al., 2006). However, upon an analysis of the information published by Chaigneau et al. (2006) which included 25 years of data, we noticed an inadequacy in the statistical analyses due to the accumulated variance produced by merging different, independent variables in the analysis, such as the intensity of El Niño/La Niña, months presenting different climatic events and seasons which were grouped rather than analyzed individually. In addition, the region was irregularly sampled on an interannual scale, and with high sampling error in the calculation of the u and v components (buoy position errors and the wind drag effect on them). Likewise, Fiedler (2002) compared climatic disturbances of different intensities, a year of extreme El Niño and two moderate La Niña years, to derive biological data related with the surface circulation. The latter makes it difficult to determine the actual dynamics of the currents in the Panama Bight, during the major reproductive period of marine species (Sep-Nov, i.e. ictioplancton, Escarria et al., 2006 and corals, Baird et al., 2009), and to understand how the main currents respond to specific climatic disturbance intensity (moderate), making it necessary to follow an experimental design.
Different studies hypothesize that during El Niño there will be an increase in the velocity and volume of transport of the NECC (Wyrtki, 1985; Kessler and Taft, 1987; Glynn et al., 1996; Glynn and Ault, 2000) and a decrease in the velocity of the SEC (Kessler and Taft, 1987; Kessler and McPhaden, 1995; Lukas, 2001; Kessler, 2006). On the other hand, during La Niña years, an increase in the velocity of the SEC is expected (Lukas, 2001). Theories indicate a variation in the velocity of the CoaC and the anticyclonic eddy during El Niño or La Niña but they do not explain how it will vary (Chaigneau et al., 2006).
Consequently, this study seeks to determine whether statistical differences exist in the magnitude of the velocity of the main ocean surface currents (NECC, SEC-northern hemisphere only, and CoaC) and the anticyclonic eddy within the Panama Bight. The comparison will be for particular climatic disturbance intensity: moderate El Niño and La Niña years versus neutral years, for a season (Sep-Nov, reproductive season in autumn). Understanding the spatial-temporal behavior and variability of the ocean surface circulation is important given that it affects the regional connectivity of organisms from different strategic marine ecosystems (Fiedler, 2002; Sale et al., 2010).
MATERIALS AND METHODS
Using the Oceanic Niño Index (ONI) we selected four years (1994, 2002, 2004 and 2006) with moderate El Niño (0.5 to 1.5 °C), three years (1995, 1998 and 2007) with moderate La Niña (-0.5 to -1.3 °C), and four years (1993, 1996, 2001 and 2005) which are neutral (-0.4 to 0.4 °C; http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.shtml). During these years, data was selected exclusively for September to November (biased towards autumn; see Johnson et al., 2002) given its ecological importance, i.e., high reproductive activity displayed by marine organisms with a pelagic phase (Lalli and Parsons, 1993; Escarria et al., 2006; Baird et al., 2009).
The Collecte Localisation Satellites (CLS, France) provided the surface current velocity from satellite data. The total current (Vt) was estimated as the sum of the surface geostrophic current (Vg) and the Ekman current (Ve):
Vt = Vg + Ve
Using the geostrophic relation, Vg was directly obtained from altimeterderived absolute dynamic topography (η) data:
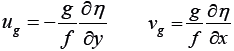
Where, g refers to gravity and f to the Coriolis parameter.
The dynamic topography data was based on the daily values of the Maps of Absolute Dynamic Topography (MADT) provided by AVISO (http://www.aviso.oceanobs.com/index.php?id=1271), in delayed time, on a 1/3° x 1/3° Mercator grid.
The Ekman component of the current (Ve) was computed as a function of the surface wind stress using the Río and Hernández model (2003). Daily wind stress data was positioned in the same grid and scale of the geostrophic current estimates (on a 1/3° x 1/3° Mercator grid), they were derived from 6-hour wind stress value provided by the European Centre for Medium Range Weather (ECMWF), with a 6 hour temporal resolution and 0.5° spatial resolution (longitude/latitude).
Velocity calculations provided by CLS already include tide and wave corrections. Coastal areas (under 50 km) were excluded, where significant deviations of the geostrophic balance occur, as well as errors due to the diffraction effect of the coast on the radar signal. Similarly, we excluded data from the equatorial band between 0-1º N, following the theory of geostrophic balance, where the Coriolis force and the Ekman transport are weakened generating errors in the calculation of the velocity; other sources of error may be the equatorial upwelling (Pascual et al., 2006), the amplitude of internal tides (Niwa and Hibiya, 2001), and the wind flow energy generated by the ocean's inertial motion (Jiang et al., 2005).
Therefore, we conducted daily and subsequently monthly calculations of the velocity of the total surface current, for each of the three main currents and the anticyclonic eddy present in the Panama Bight. From this data, a quarterly average per year was estimated (September-November; every month of each year considered as a repetition) and the vectors graphed in each grid for the region studied (1/3 x 1/3 of degree; 2° S-9° N, 75°-94° W). The average monthly and quarterly estimation of the total surface current intensity was obtained by averaging the zonal (u) and the meridional velocity (v) components (30 to 90 days, respectively).
For the statistical analysis and interannual comparison, geographical coordinates were defined for each current and for the anticyclonic eddy (plots, Figure 1b). The selection of the geographic coordinates for the NECC was based on the study carried out by Chaigneau et al. (2006) where for 25 years, the NECC has presented three entry trajectories into the Panama Bight domain, between 5-6° N and 90° W, which justifies the selection of the coordinates (Table 1). Therefore, three criteria were established to select a plot: 1) display the general circulation behavior, 2) contain the greatest quantity of reliable velocity data, and 3) prevent interference between neighboring currents.
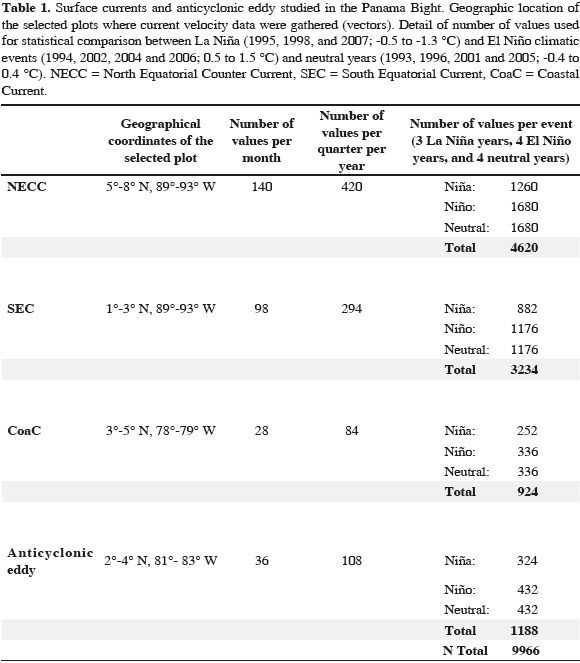
We used a total of 9966 values of average velocity distributed according to current and climatic event for statistical comparisons (Table 1). The number of vectors used per repetition for the analysis differed because of the number of years and the plot size. Only the vectors near its center were considered for the anticyclonic eddy and given its westward movement over time, according to the climatic event, the geographic coordinates used for the statistical comparison varied temporally.
Then, the null hypothesis of no differences in current velocity was tested by comparing moderate El Niño, moderate La Niña, and neutral years. To do this we reviewed the normality assumptions with and without transformation of the current velocity variable (Normality, Kolmogorov-Smirnov; Homogeneity of variance, Levene or Barlett; Independence, X2, using STATISTICA 8.0). Upon noncompliance of the assumptions, the Kruskal-Wallis nonparametric test was employed (more than two independent groups), as well as the Mann Whitney test (two independent groups). The results are represented as multi-year, quarterly average velocity figures per climatic event and per current. Changes in the direction of the circulation and the number of cyclonic and anticyclonic eddies were registered under climatic events.
RESULTS
The magnitude of the NECC surface velocity was significantly different between events (Kruskal-Wallis test, H2= 188.61, n= 4620, p= 0.000; Figure 2a); this was greater for neutral years, followed by years of moderate El Niño and La Niña events (Mann Whitney U test; Niña-Niño U= 801211.0, N1= 1260, N2= 1680, p= 0.000; Niña-neutral U= 767653.5, N1= 1260, N2= 1680, p= 0.000; Niño-neutral U= 1346350, N1= 1680, N2= 1680, p= 0.021).
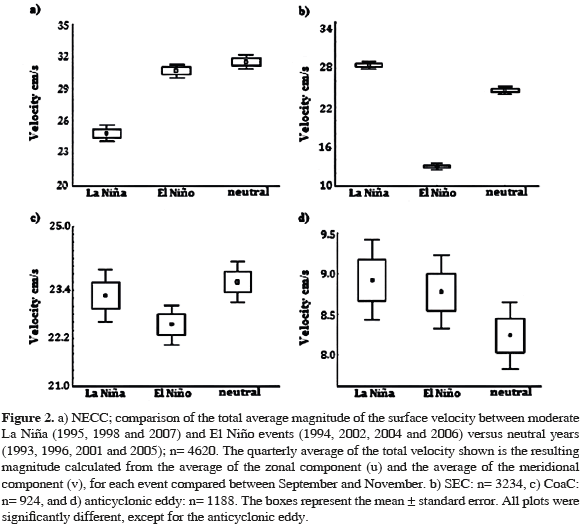
Differences were also found in the velocity magnitude for the SEC (northern hemisphere) among the events compared (Kruskal-Wallis test, H2= 1254.73, n= 3234, p= 0.000; Figure 2b). The magnitude was greater during La Niña years, followed by neutral and El Niño years (Mann Whitney test; Niña-Niño U= 977422.0, N1= 882, N2= 1176, p= 0.000; Niña-neutral U= 392095.5, N1= 882, N2= 1176, p= 0.000; Niñoneutral U= 231584.0, N1= 1176, N2= 1176, p= 0.000).
The magnitude of the Coastal Current speed (CoaC) was also different among the events compared (Kruskal-Wallis test, H2= 7.68, n= 924, p= 0.021; Figure 2c), being lower during El Niño years and equal on average for La Niña and neutral years (Mann Whitney, Niña-Niño U= 39061.5, N1= 252, N2= 336, p= 0.108; Niña-neutral U= 40727.0, N1= 252, N2= 336, p= 0.429; Niño-neutral U= 49431.0, N1= 336, N2= 336, p= 0.005).
Lastly, the velocity magnitude of the anticyclonic eddy was not significantly different between moderate El Niño, La Niña, and neutral years (Kruskal-Wallis test, H2= 5.38, n= 1188, p= 0.067; Figure 2d); however, it was slightly higher for La Niña years and lower for neutral years. Thus, the velocity analysis of the three surface currents (Figure 2a-c) revealed that during moderate El Niño conditions their magnitudes diminished with respect to neutral conditions. In this sense, the SEC velocity showed the greatest difference when comparing the averages between El Niño and neutral years (Figure 2b), followed by the NECC with a mean velocity difference of ~4.4 cm/s (Figure 2a). The lowest differences in mean velocity were found in the CoaC with ~1.1 cm/s (Figure 2c). During La Niña, the velocity magnitude with respect to neutral years depends on the current; greater for the SEC and lower for the other currents.
At the qualitative level, observing the interannual variation of the zonal and meridional component of the currents, it could be generalized for the quarter analyzed that the direction of the NECC during neutral and La Niña years was eastward, between 4° and 9° N, while during El Niño years it changed direction slightly, first towards the Southeast and later towards the Northeast, surrounding a cyclonic eddy at 7-9º N, 87-93º W (Figure 3a). According to Chaigneau et al. (2006) this current may feed the Costa Rican Coastal Current. The SEC did not modify its direction, always flowing westward between 1° and 3° N during moderate El Niño, La Niña, and neutral years (Figure 3b). The CoaC flowed southward during the three events compared (Figure 3c). The main anticyclonic eddy was found at 2-4º N, 80- 83º W for neutral years, moving westward during La Niña years (81-85º W) and El Niño years (81-83º W; Figure 3d).
Furthermore, the study area presented two cyclonic eddies in front of the coasts of Panama and Costa Rica during the three events compared and three additional eddies (two cyclonic and one anticyclonic) during moderate El Niño years, all between 5-9º N, 79-87º W and associated to the NECC (Figure 4).
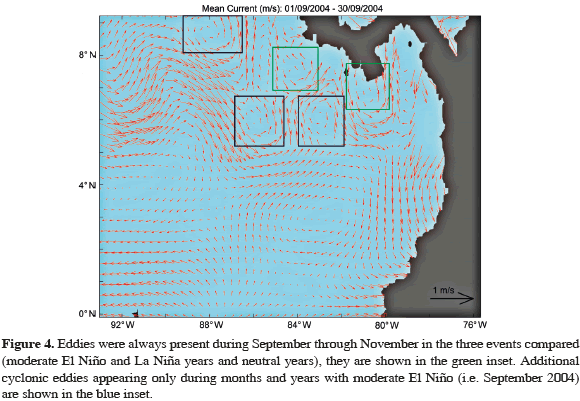
DISCUSSION
The NECC presented decreased mean velocity magnitude during moderate El Niño years (11.7 %) and moderate La Niña years (25.8 %) when compared to neutral years. This is contrary to that predicted by Glynn and Ault (2000), Johnson et al., (2002), Kessler (2002, 2006), and Chaigneau et al. (2006) where it is expected during El Niño years that the NECC should increase the volume of transport. Differences could be attributed, in part, to the fact that studies like the one performed by Chaigneau et al. (2006) do not limit the independent variable to a type of disturbance intensity or to a particular climatic season. The increment of the NECC velocity in other studies would generally be because of weakened trade winds and the presence of strong winds from the West propagating to the East, generating a series of equatorial Kelvin Waves which displace abnormally warm masses from the Western Pacific to the American coast (Seidel and Giese, 1999; IDEAM, 2002; Chen and Cane, 2008).
An explanation of the diminished NECC mean velocity during moderate El Niño years, compared to neutral years, is the influence exerted over this current by the cyclonic eddy originating between coordinates 7°-9° N and 87°-93° W during the months sampled in the area studied (Sep-Nov), which affects NECC velocity and direction. This eddy (Kessler, 2002) is originated by local changes in the direction of the predominant wind (trade winds from the North rather than from the South, according to Devis-Morales et al., 2008), causing a displacement of the surface water mass in a counter-clockwise direction (Figure 3a), which could explain the pattern found.
The predictions or generalizations indicated by several authors regarding NECC velocity during El Niño years (Glynn and Ault, 2000; Kessler, 2006; Chen and Cane, 2008) consider a large geographical area, climatic events with different intensities and different temporal scales, annual or seasonal (lower NECC velocity in Spring and towards the East; Wyrtki, 1961); however, they have not mentioned the effect of the local eddies, which could completely alter the interpretation of connectivity in the ETP, being lower than expected. Biophysical circulation models developed for the Western Pacific predict strong connectivity among islands in the Western Pacific during El Niño (independent of the season; Treml et al., 2008). Similarly, it is possible to infer the presence of a potential barrier or filter preventing the dispersion of larvae from the Central Pacific towards the East during the most important reproductive period, caused by the cyclonic eddy (increasing the genetic differentiation between populations, and potential speciation). This would support the study by Combosch et al. (2008), with respect to unidirectional and restricted gene flow between the Central and Eastern Pacific, and the statement by Scheltema (1988), on the reduction of larvae occurrence from different invertebrate groups from the Central to the Eastern Pacific (gastropods, polychaetes, sipunculids, decapod crustaceans, echinoderms, and coelenterates, among others).
Therefore, the eddy could retain planktonic larvae for a longer time than their period of maturity (Pelagic Larvae Duration-PLD), eliminating biological groups with short larval duration: a few days (i.e. Amphiprion fish, PLD of 9-12 days; Almany et al., 2007) or weeks (i.e. fish of genus Pomacentrus with PLD 16- 24 days; James et al., 2002; Patterson et al., 2005), diminishing potential functional connectivity (Grantham et al., 2003; Kinlan and Gaines, 2003; Sale et al., 2010) and increasing the probability of extinction of isolated and smaller populations (Glynn and Ault, 2000). Ekman (1953) was the first to suggest the Eastern Pacific as the most effective barrier against larval dispersal; however, Glynn and Ault (2000) refer to it as a filter bridge for larvae dispersion between the Indo-West Pacific and the East Pacific.
Likewise, it is known that El Niño generates negative physical effects on the pelagic ecosystem by diminishing the primary and secondary productivity (thermocline and the nutricline at greater depths), necessary for the survival of some larvae species (Ballance et al., 2006). This supports the recent theory which states the importance of self-seeding for the maintenance of local populations (Clarke, 1995; Figueira, 2009; Munday et al., 2009) when they are not able to receive immigrants (rescue effect for declining populations).
Nevertheless, if there were no physical connectivity barriers, for example in another season of the year, some larvae species would have the potential to enter the Panama Bight (90° W) and within ~20 days reach the isle of Cocos (5° N, 87° W) or within ~40 days reach Coiba (7° N, 81° W), assuming that the NECC flows at an average (non-linear) velocity of 30 cm/s and connects to the Panama Bight cyclonic eddy (only present in the first months of the year) where the larvae would remain trapped for two more months according to findings by Chaigneau et al. (2006). In that scenario, only larvae with long pelagic duration (long-distance dispersal) or rafting of settled stages (propagules) will survive the journey to reach the habitat for settling [e.g., coral larvae of Pocillopora damicornis that remain competent for >100 d (Richmond, 1989), and lobster larvae, which spend over six months in the plankton (Goldstein et al., 2008)]. This potential dispersal route via the NECC was proposed by Glynn and Wellington (1983) and Glynn and Ault (2000) to explain the recent colonization of modern reef-building corals in the eastern Pacific arriving from the central Pacific, after the closure of the Central American portal, ca. 3.7-3 Ma (Coates and Obando, 1996).
During neutral years, larval exchange among distant populations could be favored, given that larval transit time is reduced in the NECC, this could increase survival, dispersal kernel distribution, settlement (particularly at sink population), and genetic variability (due to immigration) of marine populations in the Colombian Pacific (e.g., islands of Malpelo and Gorgona) and in consequence, their persistence and adaptability to climate changes (evolutionary connectivity; Sale et al., 2010). This current would favor transport and survival of teleplanktonic larvae from the Central (Clipperton Atoll; Glynn and Ault, 2000) or Western Pacific whose duration is over 80 days (Grantham et al., 2003; Kinlan and Gaines, 2003); with the ETP acting as a selective filter for the dispersion of different marine taxa (Scheltema, 1988; Glynn and Ault, 2000). Developmental stages of cnidarians, polychaete worms, sipunculans, gastropods, decapods crustaceans, echinoderms, and fish have been collected alive in the east-flowing NECC and the west-flowing SEC, suggesting significant genetic exchange (Glynn and Ault, 2000). Suggestions of connectivity in the past among islands from the Central Pacific and the Colombian Pacific have been reported by Glynn and Ault (2000) through the demonstration of similarity between the assembly of coral species.
The SEC (northern hemisphere) presented a 51.6 % increased mean velocity during moderate La Niña years when contrasted with moderate El Niño years and a 16.5 % increase with relation to neutral years. This minimum value of SEC velocity during El Niño years would be explained by the appearance of strong winds from the West, this leads to massive water transfer in a West-East direction (Kessler, 2006), contrary to the SEC flow direction. The pattern observed coincides with that expected in the literature (Dana, 1975; Kessler and Taft, 1987; Kessler and McPhaden, 1995; Lukas, 2001; Kessler, 2006). It also agrees with that established by Lukas (2001) regarding increased SEC mean velocity for La Niña years, independent of the spatio-temporal scale of the generalizations, making this current somewhat more stable than the NECC in terms of the expected velocity pattern. According to IDEAM (2000) and CCCP (2002), the SEC circulation would be due to cooling conditions of the water mass, strengthening of trade winds from the southeast that cross the Colombian Pacific (Intertropical Convergence Zone between 14 and 11º N between September and November, respectively), and to intensifying equatorial upwelling (2-5º N, 80º W).
Thereby, SEC-facilitated connectivity would be favored during La Niña events, when its velocity is increased, this increment would permit marine larvae from the Eastern Tropical Pacific coast (i.e. Panama, Colombia, Ecuador) to reach distant populations found in islands in the Central Pacific. The latter idea agrees with the “vortex model”, a dispersion hypothesis postulating a predominantly East to West coral dispersal (Jokiel and Martinelli, 1992). Although, there is indirect biological evidence that this could occur (Scheltema, 1988; Glynn and Ault, 2000), this hypothesis, however, still needs to be demonstrated. Diminished SEC velocity during moderate El Niño events (Kessler, 2006) could imply that larvae with short PLD would not survive the voyage to settle and colonize islands of the Central Pacific. Chaigneau et al. (2006) showed that some buoys in the NECC are deflected towards the SEC during certain times of the year; so, survival of larvae following this spatial pattern, outside of the domain, would be very low. This study could support the reciprocal gene flow theory, where potential larvae movement is possible from East to West (SEC) and from West to East (NECC) along the Tropic Pacific.
Regarding the CoaC, it presented its minimum mean velocity value during years of moderate El Niño, being 4.4 % lower in relation to the mean velocity value for neutral and La Niña years. Diminished velocity during El Niño years could be due to the NECC direction, which is perpendicular to the CoaC southward direction, reducing its magnitude. It could also be explained by diminished wind intensity in Colombia, the appearance of winds from the West near Panama upon the descent of the Intertropical Convergence Zone and the presence of upwelling (193-476 kilometers long, 97-145 kilometers wide, 100 m thick) south of the Colombian Pacific (CCCP, 2002).
The CoaC could be very important for demographic connectivity, given that it could be the current in charge of transporting and distributing pelagic larvae along the Colombian Pacific coast, including larvae arriving from the Central Pacific, transported by the NECC together with the Panama Bight cyclonic eddy (Chaigneau et al., 2006). Likewise, marine species with a pelagic phase reproducing in the North of the Colombian Pacific from September to November could send their reproductive material to the populations found to the South (downstream). Nevertheless, as indicated by Rodríguez-Rubio et al. (2003) the CoaC changes direction, moving northward during February to March, upon modification of the wind pattern, which in theory permits dispersion, replacement of genetic material, and bidirectional connectivity for those populations with two reproductive cycles per year (e.g., broadcast spawner species, with teleplanktonic larvae capable of long distance dispersal). In species reproducing at the beginning of the year, where the turbulence, velocity, and kinetic energy of the water mass in the domain are greatest (Chaigneau et al., 2006), their reproductive material would potentially disperse northward (Panama), where buoy data indicates it would be trapped in the Panama Bight cyclonic eddy for up to two months (Chaigneau et al., 2006). What has been demonstrated, is that genetic connectivity is limited for populations of mangrove plants (e.g., Avicenia germinans) between the North and South of the Colombian Pacific, evidenced by their genetic-population structure (Cerón-Zousa et al., 2005).
Species and populations with low-dispersion strategies (i.e. brooders, short PLD) may have evolved to reproduce during months with lower current velocities (i.e. October-November) or have developed larvae with swimming (e.g. lobster larvae swimming at 15 cm per second for 2-4 weeks) or behavior abilities (changing orientation according to particular cues from suitable substratum) that allow them to overcome prevailing currents or to be close to the natal habitat-self-recruitment (Sale et al., 2010), while those depending on large-scale dispersion would reproduce during winter, when the current velocity and energy is greater (January-March; Chaigneau et al., 2006). This coastal current (Pineda, 1995) could also indirectly help to export larvae generated in Colombia to other sites in the Central Pacific, by directing larvae towards the SEC. Low genetic divergence, suggesting genetic flow and population connectivity across the entire Eastern Pacific Ocean Basin, have been documented for crabs (Huber, 1985), fish, and sea-stars (Nishida and Lucas, 1988). This would explain the long distance dispersal proposed by Glynn and Ault (2000) between the Colombian Pacific Islands and the Central Pacific.
The large radius and long-life (months) anticyclonic eddy presented its maximum mean velocity values during La Niña years, possibly as a consequence of the Chocó Jet, which affects the Panama Bight region, entering the basin at 79° W (Devis-Morales et al., 2008), and the anomalies in the ocean's thermal structure (Kessler, 2002). Although the basic physics for eddy generation is the Ekman pumping associated to wind stress, Muller-Karger and Fuentes-Yaco (2000) and Willett et al. (2006) indicate that this mechanism does not necessarily explain the formation of all the eddies from the ETP and, therefore, other processes must be explored as indicated by the numerical models of Zamudio et al. (2001).
However, the anticyclonic eddy could fulfill two roles in terms of population connectivity; the first, to function as a barrier or trap for the dispersion between the Eastern Pacific and Central Pacific, if larvae species arriving from the CoaC are trapped for months in the swirling circulation pattern (Steneck et al., 2009), maturing there and perishing (mortality sink). Given that the anticyclonic eddy has a high mean kinetic energy of ~800 cm2/s2 and an eddy kinetic energy of ~200 cm2/s2 (Chaigneau et al., 2006), it would be difficult for larvae to escape from the eddy, the only known mechanisms could be to swim against the current or to change buoyancy to favor vertical migration (Sale et al., 2010). Nonetheless, it could be beneficial for those species with pelagic teleplanktonic larvae (month or months of duration; Scheltema, 1988) given that initially they could be exported by the CoaC, but upon entering the anticyclonic eddy, they may again return to the continent and enter the NECC and then the CoaC. This is important for some marine species, since they need to develop different larval stages offshore, and when larvae matures (after some weeks or months) it returns to its natal habitat to complete its life cycle (Sale et al., 2010).
The appearance of additional cyclonic eddies during El Niño years seem to be associated with upwelling and seasonal climate variability (DIMAR, 2007). But according to Willet et al. (2006), vorticity conservation when the NECC turns North, when nearing to the Central America coast, could be the mechanism generating eddies in the ETP. Some eddies located near Panama and Costa Rica could be related to the instability of the Countercurrent and the Costa Rican Dome (Zamudio et al., 2001; Kessler, 2002); however, the mechanism still needs to be confirmed. In this scenario of moderate El Niño, the high number of eddies observed would significantly reduce the probability of larvae from the Central Pacific being transported by the NECC to settle in the American continent, supporting the selective permeability hypothesis proposed by Scheltema (1988) and Glynn and Ault (2000).
In conclusion, the ocean surface circulation in the Panama Bight from September to November indicates that during moderate La Niña years the NECC diminishes its flow velocity with respect to neutral years. The NECC also diminishes its flow velocity and increases the number of cyclonic eddies during El Niño events. The SEC (northern hemisphere) increases its velocity during La Niña years but diminishes during El Niño years, while the CoaC presents its minimum value during El Niño years, flowing southward, parallel to the Colombian coast. The main anticyclonic eddy presented its maximum velocity values during La Niña years. This all suggests that moderate El Niño and La Niña events affect the circulation in the Panama Bight in different ways.
Finally, the currents and eddies studied operate comprehensively as a conveyor belt, moderating the dispersal and reciprocal gene flow between the East and West Pacific and acting as a dispersion filter bridge during the quarter studied. Where the amount of eddies during El Niño would reduce the probability of NECC-facilitated larval connectivity between the Central and Eastern Pacific; and the CoaC and SEC would favor larva export from the American continent to the Central Pacific, as long as they manage to overcome the anticyclonic eddy.
ACKNOWLEDGEMENTS
The authors thank the Academic Vice Rectory at Pontificia Universidad Javeriana, Bogotá, Colombia for the financial support (ID PPTA 4159 and 4503). We also thank Conservación Internacional Colombia for their financial support. We are grateful to Sandra Bessudo and Germán Soler from Fundación Malpelo for facilitating and creating the liaison with the Collecte Localisation Satellites-France; to Mauricio Romero (M. Sc.) and Iván Pérez (Ph. D. Universidad de Concepción - Chile) for their valuable comments on the document and to Martin E. Rojas, Albert Ortiz (City College of the City University of New York) and to Gypsy Español (Traducciones Técnicas T&T) for the English proofreading. The authors also wish to thank four anonymous reviewers for their extensive comments and advice, which helped shape the final version of this manuscript. The authors declare that they have no conflict of interest.
LITERATURE CITED
1 Almany, G. R., M. L. Berumen, S. R. Thorrold, S. Planes and G. P. Jones. 2007. Local replenishment of coral reef fish populations in a marine reserve. Science, 316: 742-744. [ Links ]
2 Baird, A. H., J. R. Guest and B. L. Willis. 2009. Systematic and biogeographical patterns in the reproductive biology of Scleractinian corals. Annu. Rev. Ecol. Evol. Syst., 40: 551-71. [ Links ]
3 Ballance, L. T., R. L. Pitman and P. C. Fiedler. 2006. Oceanographic influences on seabirds and cetaceans of the eastern tropical Pacific: a review. Progress in Oceanography, 69: 360-390. [ Links ]
4 CCCP. 2002. Compilación oceanográfica de la cuenca Pacífico colombiana. Centro Control Contaminación del Pacífico, Tumaco, Colombia. 109 p. [ Links ]
5 Cerón-Zouza I., N. Toro-Perea and H. Cardenas-Henao. 2005. Population genetic structure of neotropical mangrove species on the Colombian Pacific coast: Avicennia germinans (Avicenniaceae). Biotropica, 37: 258-265. [ Links ]
6 Chaigneau, A., R. Abarca del Río and F. Colas. 2006. Lagrangian study of the Panama Bight and surrounding regions. J. Geophys. Res., 111: C09013. [ Links ]
7 Chávez, F. P., P. G. Strutton, G. E. Friederich, R. A. Feely, G. C. Feldman, D. G. Foley and M. J. McPhaden. 1999. Biological and chemical response of the equatorial Pacific ocean to the 1997-98 El Niño. Science, 286: 2126-2131. [ Links ]
8 Chen, D. and M. Cane. 2008. El Niño prediction and predictability. J. Comput. Phys., 227: 3625-3640. [ Links ]
9 Clarke, T. 1995. Larvae of nearshore fishes in oceanic waters of the central equatorial Pacific. Pac. Sci., 49: 134-142. [ Links ]
10 CMAR. 2004. Corredor marino de conservación entre las islas de Galápagos, Coco, Malpelo, Gorgona y Coiba. Gobiernos de Ecuador, Costa Rica, Colombia y Panamá. http://www.cmarpacifico.org/Docs/Declaracion%20Conjunta%20%20Ministros%20CMAR[PDF].pdf. 28/01/2012. [ Links ]
11 Coates, A. G. and J. A. Obando. 1996. The geologic evolution of the Central American isthmus. 21-56. In: Jackson, J. B. B., A. F. Budd and A. G. Coates (Eds.). Evolution and environment in tropical America. Univ Chicago Press, Chicago. 409 p. [ Links ]
12 Combosch, D. J., H. M. Guzman, H. Schuhmacher and S. V. Vollmer. 2008. Interspecific hybridization and restricted trans-Pacific gene flow in the Eastern Tropical Pacific Pocillopora. Mol. Ecol., 17: 1304-1312. [ Links ]
13 Cowen, R. and S. Sponaugle. 2009. Larval dispersal and marine population connectivity. Annu. Rev. Mar. Sci., 1: 443-66. [ Links ]
14 Cowen, R., C. Paris and A. Srinivasan. 2006. Scaling of connectivity in marine populations. Science, 311: 522-527. [ Links ]
15 Crooks, K. R. and M. Sanjayan. 2006. Connectivity conservation: maintaining connections for nature. 1-20. In: Crooks, K. R and M. Sanjayan (Eds.). Connectivity conservation. Cambridge University Press, Cambridge. 726 p. [ Links ]
16 Dana, T. 1975. Development of contemporary Eastern Pacific coral reefs. Mar. Biol., 33: 355-374. [ Links ]
17 D'Croz, L. and A. O'Dea. 2007. Variability in upwelling along the Pacific shelf of Panama and implications for the distribution of nutrients and chlorophyll. Estuarine, Coastal and Shelf Science, 1-16. [ Links ]
18 Devis-Morales, A., W. Schneider, R. Montoya-Sánchez and E. Rodríguez-Rubio. 2008. Monsoon-like winds reverse oceanic circulation in the Panama Bight. J. Geophys. Res. Lett., 35: L20607. [ Links ]
19 DiBacco, C., L. A. Levi and E. Sala. 2006. Connectivity in marine ecosystems: the importance of larval and spore dispersal. 185-212. In: Crooks, K. R and M. Sanjayan (Eds.). Connectivity conservation. Cambridge University Press, Cambridge. 726 p. [ Links ]
20 DIMAR-CCCP and UAESPNN-DTSO. 2007. Santuario de Fauna y Flora Malpelo: descubrimiento en marcha. Dirección General Marítima-Centro Control Contaminación del Pacífico y Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales-Dirección Territorial Suroccidente. Ed. DIMAR, Serie de Publicaciones Especiales CCCP Vol.5, Bogotá. 142 p. [ Links ]
21 Ekman, S. 1953. Zoogeography of the sea. Sidgwick and Jackson, London. 417 p. [ Links ]
22 Escarria, E., B. Beltrán-León and A. Giraldo. 2006. Ictioplancton superficial de la cuenca del Océano Pacífico colombiano (septiembre de 2003). Invest. Mar., 34 (2): 169-173. [ Links ]
23 Fiedler, P. C. 2002. The annual cycle and biological effects of the Costa Rica Dome. Deep Sea Res. Parte 1, 49: 321-338. [ Links ]
24 Figueira, W. 2009. Connectivity or demography: Defining sources and sinks in coral reef fish metapopulations. Ecol. Model., 220: 1126-1137. [ Links ]
25 Glynn, P. and J. Ault. 2000. A biogeographic analysis and review of the far eastern Pacific coral reef region. Coral Reef, 19: 1-23. [ Links ]
26 Glynn, P. and G. M. Wellington. 1983. Corals and corals reefs of the Galápagos Islands. University Of California, Berkeley. 330 p. [ Links ]
27 Glynn, P., J. Veron and G. M. Wellington. 1996. Clipperton Atoll (eastern Pacific): oceanography, geomorphology, reef-building coral ecology and biogeography. Coral Reefs, 15: 71 - 99. [ Links ]
28 Goldstein, J., L. Pejchar and D. Gretchen. 2008. Using return-on-investment to guide restoration: a case study from Hawaii. Cons. Let., 1: 236-243. [ Links ]
29 González-Silvera, A., E. Santamaria de Ángel, R. Millan-Nuñez and H. Manzo-Monroy. 2004. Satellite observations of mesoscale eddies in the Gulfs of Tehuantepec and Papagayo (Eastern Tropical Pacific). Deep Sea Research II, 51: 587-600. [ Links ]
30 Grantham, B. A., G. L. Eckert and A. L. Shanks. 2003. Dispersal potential of marine invertebrates in diverse habitats. Ecol. Appl., 13: S108-S116. [ Links ]
31 Grodsky, S. and J. Carton. 2001. Intense surface currents in the tropical Pacific during 1996-1998. J. Geophys. Res., 106: 16673-16684. [ Links ]
32 Hanski, I. and D. Simberloff. 1997. The metapopulation approach, its history, conceptual domain, and application to conservation. 5-26. In: Hanski, I. A. and M. E. Gilpin (Eds.) Metapopulation biology: Ecology, genetics and evolution. Academic Press, San Diego. 512 p. [ Links ]
33 Huber, M. 1985. Population genetics of eight species of Trapezia (Brachyura, Xanthidae), symbionts of corals. Mar. Biol., 85: 23-36. [ Links ]
34 IDEAM. 2000. Movimientos en masa dañinos ocurridos en Colombia durante el fenómeno frío del Pacífico (La Niña) 1999-2000. República de Colombia, Ministerio del Medio Ambiente. Instituto de Hidrología, Meteorología y Estudios Ambientales, Bogotá. 250 p. [ Links ]
35 IDEAM. 2002. Efectos naturales y socioeconómicos del fenómeno El Niño en Colombia. República de Colombia, Ministerio del Medio Ambiente, Instituto de Hidrología, Meteorología y Estudios Ambientales, Bogotá. 250 p. [ Links ]
36 James, E. K., P. Gyaneswar, N. Mathan, W. L. Barraquio, P. M. Reddy, P. P. Iannetta, F. L. Olivares and J. K. Ladha. 2002. Infection and colonization of the rice seedlings by the plant growth-promoting bacterium Herbaspirillum seropedicae Z67. Mol. Plant Microb. Interact., 15:894-906. [ Links ]
37 Jiang, J., Y. Lu and W. Perrie. 2005. Estimating the energy flux from wind to ocean inertial motions: The sensitivity to surface wind fields. J. Geophys. Res. Lett., 32: L15610. [ Links ]
38 Johnson, G., B. Sloyan, W. Kessler and K. McTaggart. 2002. Direct measurements of upper ocean currents and water properties across the tropical Pacific during the 1990s. Prog. Oceanogr., 52: 31-61. [ Links ]
39 Jokiel, P. and F. J. Martinelli. 1992. The vortex model of coral reef biogeography. J. Biogeogr., 19: 449- 458. [ Links ]
40 Kessler, W. 2002. Mean three-dimensional circulation in the north eastern tropical Pacific. J. Phys. Oceanogr., 32: 2457-2471. [ Links ]
41 Kessler, W. 2006. The circulation of the eastern tropical Pacific: A review. Prog. Oceanogr., 69: 181-217. [ Links ]
42 Kessler, W. and M. McPhaden. 1995. The 1991-93 El Niño in the central Pacific. Deep-Sea Research II, 42: 295-333. [ Links ]
43 Kessler, W. and B. Taft. 1987. Dynamic heights and zonal geostrophic transports in the central tropical Pacific during 1979-84. J. Phys. Oceanogr., 17: 97-122. [ Links ]
44 Kinlan, B. P. and S. D. Gaines. 2003. Propagule dispersal in marine and terrestrial environments: a community perspective. Ecology, 84: 2007-2020. [ Links ]
45 Lalli, C. M. and T. R. Parsons. 1993. Biological oceanography: An introduction. University of British Columbia. Pergamon Press, Oxford. 301 p. [ Links ]
46 Lavin, M. F., P. C. Fiedler, J. A. Amador, L. T. Balance, J. Farber-Lorda and A. M. Mestas-Nuñez. 2006. A review of eastern tropical Pacific oceanography: Summary. Progr. Oceanogr., 69: 391-398. [ Links ]
47 Lukas, R. 2001. Pacific ocean equatorial currents. 103-110. In: Steele, J. H, S. A. Thorpe and K. K. Turekian (Eds.). Ocean currents. A derivative of Encyclopedia of Ocean Sciences, second edition. Academic Press, London. 612 p. [ Links ]
48 Muller-Karger, F. E. and C. Fuentes-Yaco. 2000. Characteristics of wind-generated rings in the Eastern Tropical Pacific Ocean. J. Geophys. Res. Oceans., 105: 1271-1284. [ Links ]
49 Munday, P., J. Leis, J. Lough, C. Paris, M. Kingsford, M. Berumen and J. Lambrechts. 2009. Climate change and coral reef connectivity. Coral Reefs, 28: 379-395. [ Links ]
50 Nishida, M. and J. S. Lucas. 1988. Genetic differences between geographic populations of the crown-ofthorns starfish throughout the Pacific region. Mar. Biol., 98: 359-368. [ Links ]
51 Niwa, Y. and T. Hibiya. 2001. Numerical study of the spatial distribution of the M2 internal tide in the Pacific Ocean. J. Geophys. Res., 106: 22441-22449. [ Links ]
52 Palumbi, S. R. 2004. Marine reserves and ocean neighborhoods: the spatial scale of marine populations and their management. Ann. Rev. Envir. Res., 29: 31-68. [ Links ]
53 Pascual, A., Y. Faugére, G. Larnicol and P. Le Traon. 2006. Improved description of the ocean mesoscale variability by combining four satellite altimeters. J. Geophys. Res. Lett., 33: 2611. 4 p. [ Links ]
54 Patterson, J., S. Deepak, E. Patterson and D. Wilhelmsson. 2005. Improving living conditions for reef dependent fisher families in Tuticorin, Gulf of Mannar, Southeast coast of India. CORDIO Report, 2005: 252-262. [ Links ]
55 Philander, S. G. 1983. El Niño Southen oscillation phenomena. Nature, 302: 295-301. [ Links ]
56 Pineda, A. R. 1995. Condiciones hidrológicas en la cuenca del Pacífico colombiano. Bol. Cient. CCCP No. 5: 73-97. [ Links ]
57 Poveda, G. and O. J. Mesa. 2000. On the existence of Lloró (the rainiest locality on earth): Enhanced ocean-land-atmosphere interaction by a low-level jet. J. Geophys. Res, Let., 27: 1675-1678. [ Links ]
58 Reyes-Bonilla, H., J. D. Carriquiry, G. E. Leyte-Morales and A. L. Cupul-Magana. 2002. Effects of the El Nino-Southern Oscillation and the anti-El Nino event (1997-1999) on coral reefs of western coast of Mexico. Coral Reefs, 21: 368-372. [ Links ]
59 Richmond, R. H. 1989. Competency and dispersal of spawned versus brooded coral planula larvae. Proc. 6th Int. Cor. Reef Symp., 2: 827-831. [ Links ]
60 Río, M. and F. Hernández. 2003. High-frequency response of wind-driven currents measured by drifting buoys and altimetry over the world ocean. J. Geophys. Res., 108: 3283. 1-19. [ Links ]
61 Roberts, C. M. 1997. Connectivity and management of Carribean coral reefs. Science, 278: 1454-1457. [ Links ]
62 Rodríguez-Rubio, E., R. Abarca del Río and W. Schneider. 2003. On the seasonal circulation with the Panama Bight derived from satellite observations of wind, altimetry and sea surface temperature. J. Geophys. Res. Lett., 30 (7): 1410. [ Links ]
63 Rodríguez-Rubio, E., J. Ortiz and J. Rueda. 2007. Aspectos oceanográficos de Isla Malpelo. 5:29-44. In: DIMAR-CCCP y UAESPNN-DTSO (Eds.). Santuario de Fauna y Flora Malpelo: descubrimiento en marcha. DIMAR, Bogotá. 142 p. [ Links ]
64 Rojas, X. 2005. Caracterización de las corrientes marinas del Pacífico colombiano, Sur América. UPC. Centro de Estudios Avanzados de Blances-CEAB, Bogotá. 250 p. [ Links ]
65 Sale, P. F., H. Van Lavieren, M. C. Ablan Lagman, M. Butler, C. Fauvelot, J. D. Hogan, G. P. Jones, K. C. Lindeman, C. B. Paris, R. Steneck and H. L. Stewart. 2010. Preserving reef connectivity: A handbook for marine protected area managers. Connectivity Working Group, Coral Reef Targeted Research and Capacity Building for Management Program UNU-INWEH. Melbourne. 78 p. [ Links ]
66 Scheltema, R. S. 1988. Initial evidence for the transport of teleplanic larvae invertebrates across the East Pacific Barrier. Biol. Bull., 174: 145-152. [ Links ]
67 Seidel, H. and B. Giese. 1999. Equatorial currents in the Pacific Ocean 1992-1997. J. Geophys. Res., 104 (C4): 7849-7863. [ Links ]
68 Steneck, R., C. Paris, S. Arnold, M. Ablan, A. Alcala, M. Butler, L. McCook, G. Russ and P. Sale. 2009. Thinking and managing outside the box: coalescing connectivity networks to build region-wide resilience in coral reef ecosystems. Coral Reefs, 28: 367-378. [ Links ]
69 Thompson, R. C., T. P. Crowe and S. J. Hawkins. 2002. Rocky intertidal communities: past environmental changes, present status and predictions for the next 25 years. Environ. Conserv., 29: 168-191. [ Links ]
70 Tomczak, M. and J. S. Godfrey. 1994. Regional oceanography: An introduction. Pergamon Press, United Kingdom, London. 422 p. [ Links ]
71 Treml, E., P. Halpin, D. Urban and L. Pratson. 2008. Modelling population connectivity by ocean currents, a graph-theoretic approach for marine conservation. Landscape Ecol., 23: 19-36. [ Links ]
72 Trenberth, K. E. 1997. The definition of El Niño. Bull. Amer. Met. Soc., 78: 2771-2777. [ Links ]
73 Trenberth, K. E. and T. J. Hoar. 1996. The 1990-1995 El Niño-Southern oscillation event: longest on record. Geophysical Research Letters, 23: 57-60. [ Links ]
74 Turner, J. 2004. The El Niño-Southern oscillation and Antarctica. Int. J. Climatol., 24: 1-31. [ Links ]
75 Werner, F. E., R. K. Cowen and C. B. Paris. 2007. Coupled biological and physical models: present capabilities and necessary developments for future studies in population connectivity. Oceanography, 20 (3): 54-69. [ Links ]
76 Willett, C. S. 1996. A study of anticyclonic eddies in the eastern tropical Pacific Ocean with integrated satellite, in situ, and modeled data. Ph.D. thesis, University of Colorado, Boulder, United States. 127 p. [ Links ]
77 Willet, C. S., R. R. Leben and M. F. Lavin. 2006. Eddies and tropical instability waves in the Eastern Tropical Pacific: A review. Prog. Oceanogr., 69: 218-238. [ Links ]
78 Wooster, W. S. 1959. Oceanographic observations in the Panama Bight, Askoy expeditions, 1941. Bull. Amer. Mus. Nat. Hist., 118 (3): 117-151. [ Links ]
79 Wyrtki, K. 1961. The thermohaline circulation in relation to the general circulation in the oceans. Deep Sea Res., 8: 39-64. [ Links ]
80 Wyrtki, K. 1967. Circulation and water masses in the eastern equatorial Pacific Ocean. International Journal of Oceanology and Limnology, 1 (2): 117-147. [ Links ]
81 Wyrtki, K. 1985. Sea level fluctuations in the Pacific during the 1982-83 El Niño. J. Geophys. Res. Lett., 12: 125-128. [ Links ]
82 Zamudio, L., A. P. Leonardi, S. D. Meyers and J. J. O'Brien. 2001. ENSO and eddies on the southwest coast of Mexico. J. Geophys. Res. Lett., 28: 13-16. [ Links ]
FECHA DE RECEPCIÓN: 20/12/2010 FECHA DE ACEPTACIÓN: 12/10/2011
