










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.40 supl.1 Santa Marta Dec. 2011
OCURRENCIA Y TASAS DE ENCUENTRO DE MAMÍFEROS MARINOS EN LAS AGUAS DE LA ISLA MALPELO Y HACIA EL CONTINENTE
OCCURRENCE AND ENCOUNTER RATES OF MARINE MAMMALS IN THE WATERS AROUND THE MALPELO ISLAND AND TO THE CONTINENT
Julio César Herrera Carmona1, Juan José Capella Alzueta1, Germán Andrés Soler2, Sandra Bessudo2, Carolina García2 y Lilián Flórez-González1
1 Fundación Yubarta, Carrera 24 No 4-32, Cali, Colombia. juliocesar.herreracarmona@gmail.com (J.C.H.C); jjcapella@yahoo.com (J.J.C.A); lilian.florezgonzalez@gmail.com (L.F.G.).
2 Fundación Malpelo y Otros Ecosistemas Marinos, Carrera 7 No 32-33 Piso 27, Bogotá, Colombia. gsoler@fundacionmalpelo.org (G.A.S); sbessudo@fundacionmalpelo.org (S.B.); caritogarcia2004@yahoo.com (C.G.).
RESUMEN
Este trabajo presenta una recopilación de avistamientos de mamíferos marinos del Santuario de Fauna y Flora (SFF) Malpelo, y las aguas que lo separan del continente. Se llevaron a cabo entre 2001 y 2010 y provienen de cuatro fuentes de información con observaciones sobre la línea de transecto para la travesía y observaciones desde punto fijo para las aguas cercanas a la isla. En el SFF se reconocieron siete especies y los meses de octubre y noviembre presentaron la mayor riqueza de especies. Tursiops truncatus presentó la mayor tasa de encuentro (17.78 grupos/100 h), seguida por Megaptera novaeangliae (1.62) y Stenella attenuata (0.88). Las tres especies se distribuyeron en aguas cercanas a la isla, a distancias menores a 6 km. Las especies restantes en aguas de Malpelo fueron Stenella coeruleoalba, S. longirostris, Delphinus delphis y Zalophus wollebaeki. En la travesía se identificaron trece especies, y los meses con mayor riqueza fueron marzo y abril. Megaptera novaeangliae presentó la mayor tasa de encuentro (5.94), seguida por T. truncatus (3.30), S. attenuata (3.08), D. delphis (3.08) y S. coeruleoalba, Globicephala macrorhynchu y Orcinus orca, las últimas tres con una tasa de 0.66 cada una. Megaptera novaeangliae se distribuyó en aguas sobre la plataforma continental, los delfines T. truncatus y D. delphis en aguas oceánicas y el delfín S. attenuata en las dos zonas. El resto de especies encontradas en la travesía fueron Steno bredanensis, Pseudorca crassidens, Grampus griseus, Peponocephala electra, Physeter macrocephalus y Ziphius cavirostris. Delphinus delphis fue significativamente más abundante en las aguas intermedias y durante el primer trimestre (enero-marzo) y T. truncatus en las aguas del Santuario, pero no mostró estacionalidad y fue la única especie presente todo el año en ambas zonas (alrededor de la isla y en aguas oceánicas). Adicionalmente sus tasas de encuentro no presentaron diferencias significativas entre los años evaluados. La presencia de M. novaeangliae fue estacional (entre junio y noviembre), con una abundancia significativamente mayor en el tercer trimestre (julio-septiembre) en Malpelo y en la travesía. La estabilidad de las tasas de encuentro de T. truncatus sugiere que se trata de individuos de una misma población. La presencia de crías de jorobada sugiere que el uso que están haciendo de Malpelo es de carácter reproductivo. Esta nueva información permite conocer mejor la riqueza de la comunidad de mamíferos marinos y comprender mejor sus patrones de ocurrencia en el área del SFF Malpelo y en las aguas que lo separan del continente.
PALABRAS CLAVE: Mamíferos marinos, Cetáceos, Pacífico colombiano, Isla Malpelo.
ABSTRACT
This work compiles a decade (2001-2010) of marine mammal sightings in the Malpelo Fauna and Flora Sanctuary (FFS) and the area between the island and mainland coast. Four separate sources of data were consulted, which used visual searching during cruising efforts while Malpelo's surrounding waters were surveyed from a vantage point. Seven species were identified in the FFS: October and November were the months with higher species richness. Tursiops truncatus had the highest encounter rates (17.78 groups/100 h), followed by Megaptera novaeangliae (1.62) and Stenella attenuata (0.88). These species were usually within 6 km from the island. Other species seen around the island include Stenella coeruleoalba, Delphinus delphis, Stenella longirostris and Zalophus wollebaeki. On the other hand, thirteen species were identified during cruises, and March and April were the months with the highest species richness. Megaptera novaeangliae had the highest encounter rate (5.94), followed by T. truncatus (3.30), S. attenuata (3.08), D. delphis (3.08) and S. coeruleoalba, Globicephala macrorhynchus and Orcinus orca, each one with 0.66. Other species seen during the cruises include Steno bredanensis, Pseudorca crassidens, Grampus griseus, Peponocephala electra, Physeter macrocephalus and Ziphius cavirostris. Megaptera novaeangliae were associated to the continental shelf, T. truncatus and D. delphis to oceanic waters, and S. attenuata to both the continental shelf and oceanic waters. Delphinus delphis was more abundant in intermediate waters, during the first trimester (January-March), while T. truncatus was the most abundant species around the Sanctuary during all seasons, suggesting that is the same population. The latter was also the only species found all year round in both zones (around the island and in oceanic waters), and encounter rates did not change across years. Megaptera novaeangliae had a seasonal presence (mostly June to November) with a higher abundance during the third trimester (July-September), both around Malpelo and the transect. The presence of humpback whales calves suggests that Malpelo is used for reproductive purposes. This new information about the marine mammals found in Malpelo FFS and the area between the island and the continent contributes to the understanding of these species in Colombian waters.
KEY WORDS: Marine mammals, Cetaceans, Colombian Pacific, Malpelo Island.
INTRODUCCIÓN
Gracias a su ubicación oceánica y a las condiciones del medio circundante, las aguas de la isla Malpelo se caracterizan por una alta diversidad biológica marina que comprende diversos grupos, entre los que se encuentran los mamíferos marinos. La historia de su investigación en la zona es reciente, y sólo a partir de 2001, con una serie de cruceros multidisciplinarios organizados por la Dirección General Marítima (DIMAR) y la Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales (UAESPNN), además de algunos viajes de buceo con observadores de cetáceos a bordo, se inició una recopilación sistemática de información, que permitió evaluar algunos aspectos ecológicos como la riqueza de especies y su distribución alrededor de la isla y en las aguas hacia el continente. Según Herrera et al. (2007), de las 27 especies registradas en el Pacífico colombiano (23 cetáceos y cuatro otáridos) (Vidal, 1990; Flórez-González y Capella, 1995; Flórez-González et al., 2004), sólo 11 se han registrado en el Santuario de Fauna y Flora (SFF) Malpelo pertenecientes a cuatro familias (Delphinidae, Physeteridae, Balaenopteridae y Otariidae). Tursiops truncatus (delfín nariz de botella) y Stenella attenuata (delfín moteado) son las especies con mayor abundancia relativa en las aguas del Santuario, mientras a lo largo de la travesía hacia el continente lo son Delphinus delphis (delfín común de hocico corto), S. attenuata y Stenella coeruleoalba (delfín listado) (Herrera et al., 2007). Este trabajo permitió establecer que la riqueza y la abundancia relativa de cetáceos en los alrededores de la isla y en las aguas hacia el continente son mayores durante el primer semestre del año.
Los trabajos realizados por la National Oceanic and Atmospheric Administration (NOAA) a finales de los 80's y principios de los 90's (Vidal, 1990; Wade y Gerrodette, 1993; Gerrodette y Palacios, 1996), que incluyeron las aguas de la Zona Económica Exclusiva (ZEE) del Pacífico de Colombia, dieron un primer antecedente de la riqueza de especies de mamíferos marinos de la Cuenca Pacífica Colombiana (CPC) y permitieron estimar las abundancias absolutas para las especies más importantes, mediante el método de transecto lineal. Herrera (2009), utilizando como plataforma de oportunidad los buques que realizan los cruceros oceanográficos Pacífico-ERFEN (2006-2008), organizados por la DIMAR y el Centro de Investigaciones Oceanográficas e Hidrográficas del Pacífico (CCCP), encontró marcadas diferencias estacionales de la abundancia relativa y la riqueza entre las dos épocas climáticas del año, que caracterizan la CPC. La menor riqueza y abundancia relativa del segundo semestre en la CPC estarían influenciadas por migraciones regionales a través del Pacífico Oriental Tropical (POT), en las que las especies de cetáceos se desplazan hacia la Contracorriente Ecuatorial del Norte (Reilly, 1990; Reilly y Fiedler, 1994), por condiciones locales que restringen su presencia. Es así como durante el segundo semestre la CPC presenta una termoclina débil y profunda (Devis-Morales et al., 2008), menor concentración de clorofila en superficie (Rodríguez-Rubio y Stuardo, 2002) y menor probabilidad de frentes térmicos (Herrera, 2009), características que probablemente inciden en la disponibilidad de presas y finalmente en la menor abundancia de cetáceos. Así mismo, durante el primer semestre del año, cuando la riqueza y la abundancia son mayores, la zona del Suroeste de la isla Malpelo presenta una de las mayores tasas de encuentro de cetáceos de todo el Pacífico de Colombia. Posteriormente, Palacios et al. (en prensa) llevaron a cabo una extensa y completa recopilación de avistamientos realizados a bordo de cruceros dirigidos y de plataformas de oportunidad entre 1986 y 2008, y representaron la distribución espacial de 19 especies y un género abarcando todas las aguas de la Zona Económica Exclusiva (ZEE) del Pacífico de Colombia. A pesar de los avances, el único documento de referencia acerca de la comunidad de mamíferos marinos del SFF Malpelo (Herrera et al., 2007), tiene una cobertura temporal limitada (2001-2005). Dada la importancia biológica de Malpelo, la inclusión del Santuario como uno de los nodos del Corredor Marino de Conservación del Pacífico Este Tropical (CMAR) y su declaratoria como Patrimonio Natural de la Humanidad, es prioritario conocer mejor la riqueza de la comunidad de mamíferos marinos y comprender mejor sus patrones de ocurrencia en el área del SFF Malpelo y en las aguas que lo separan del continente. Este trabajo recopila e integra la información disponible publicada y aquella producto de nuevos avistamientos obtenidos en los últimos cinco años en los cruceros del proyecto SEASCAPE (realizado en el marco del CMAR), de viajes de buceo a la isla, y de los segmentos de esfuerzo en el Santuario y en las aguas hacia el continente durante los cruceros oceanográficos Pacífico-ERFEN. El análisis de esta base de datos con 10 años de registros, constituye un insumo necesario para las iniciativas de manejo de los recursos marinos a nivel local (e.g., plan de manejo del área) y a nivel regional (e.g., CMAR).
ÁREA DE ESTUDIO
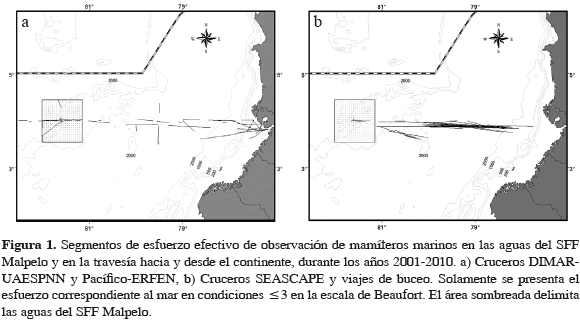
Malpelo es la única isla oceánica de Colombia en el Pacífico. Está localizada aproximadamente en el centro de la CPC a 3°59'59.96" N y 81°36'1.80" W, a 497 km al oeste de la costa del municipio de Buenaventura (López-Victoria y Rozo, 2006). En 1995 fue declarada Santuario de Fauna y Flora (SFF) en el marco del Sistema Nacional de Áreas Protegidas (SINA). La porción terrestre ocupa 1.2 km2 (López-Victoria y Rozo, 2006) y el área marina 9584 km2, y está definida por un polígono de forma cuadrada que contiene un círculo con un radio de 46.3 km con centro en la isla Malpelo (Figura 1). La profundidad de las aguas circundantes desciende rápidamente hasta 4000 m por el costado oriental. En el eje suroestenoreste se encuentra la Dorsal de Malpelo (una cordillera volcánica submarina), con una predominancia de la isobata de 2000 m, y donde se encuentra el Bajo Bojacá a una profundidad de 281 m. La climatología de Malpelo, al igual que la de la CPC, presenta un comportamiento estacional semestral determinado por el movimiento de la Zona de Convergencia Inter Tropical (ZCIT) (Stevenson, 1970; Rodríguez-Rubio et al., 2003; Rueda et al., 2007; Devis-Morales et al., 2008). Durante el segundo semestre del año, los vientos del sureste dominan toda la CPC ubicando a la ZCIT en su posición más septentrional (8-10° N) y permitiendo que predomine un patrón de circulación de aguas superficiales ciclónico en sentido contrario a las manecillas del reloj (Wooster, 1959; Wyrtki, 1966; Stevenson, 1970; Andrade, 1992). Durante el fin e inicio de año, la ZCIT se ubica en su posición más meridional debido a la predominancia de los vientos alisios del Norte, que empujan la ZCIT hacia el Sur por medio del proceso físico denominado chorro de viento de Panamá (Chelton et al., 2000; Rodríguez-Rubio et al., 2003). La fuerza de estos vientos ocasiona un debilitamiento de la corriente costera de Colombia y la aparición de la corriente de Chocó (Andrade, 1992), con flujo en dirección hacia el sur, desde el este del golfo de Panamá. Durante este período, la mayor parte del agua que sale del golfo de Panamá, donde ocurre un fuerte afloramiento y un estrechamiento de la termoclina, se desvía al oeste y se une a una circulación anticiclónica (a favor de las manecillas del reloj), centralizada cerca de los 5° N y 88° W.
MATERIALES Y MÉTODOS
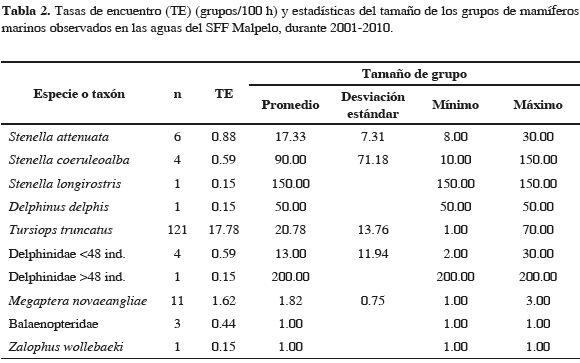
Se utilizaron cuatro fuentes de información: 1) Cuatro cruceros de investigación multidisciplinaria organizados por la DIMAR y la UAESPNN, llevados a cabo entre 2001 y 2005, a bordo de los Buques Oceanográficos de la Armada ARC Malpelo y ARC Providencia. Los dos buques presentan las mismas características generales (50.9 m de eslora, 10 m de manga y 4.4 m de calado). Sus velocidades en transecto oscilan entre 10 y 11 nudos, y la altura de la plataforma de muestreo es de 10 m. 2) Siete cruceros oceanográficos Pacífico-ERFEN, llevados a cabo entre 2004 y 2008, a bordo de las mismas unidades de investigación. 3) Doce cruceros de investigación del proyecto SEASCAPE, realizados entre 2006 y febrero de 2010, a bordo de la M/N María Patricia. La velocidad promedio fue de ocho nudos y la altura de muestreo de 4.5 m. 4) Trece viajes de buceo realizados entre 2004 y 2007, a bordo de la misma embarcación anterior. Para las dos primeras fuentes, el muestreo lo realizó un solo observador utilizando binoculares 7x50, con descansos cada dos horas durante la navegación. Para las dos últimas fuentes, el muestreo fue llevado a cabo por uno a tres observadores, con binóculos o a simple vista. A pesar de que el objetivo de este trabajo no fue la estimación de abundancias absolutas ni se ajustó una función de detección para el cálculo de densidades, en todos los casos se siguió el protocolo del método de transecto lineal (Buckland et al., 2001; Forney, 2002), el cual permitió la aproximación al cálculo de abundancias relativas mediante el conteo de grupos a lo largo de la línea de transecto. El tipo de observación fue passing mode (definición según Dawson et al., 2008), que consiste en que la embarcación continúe su travesía después de que un grupo de mamíferos marinos ha sido observado. En este caso, el tamaño del grupo y la identidad de la especie son estimados desde el transecto lineal, mientras el esfuerzo de muestreo continúa. Difiere del tipo de observación closing mode, en el que la embarcación abandona el transecto lineal, para aproximarse a los grupos avistados y poder así estimar con mayor precisión el tamaño grupal y la identidad de la especie. El campo visual cubierto fue de 90° a cada lado de la línea del transecto. Las observaciones solamente se realizaron cuando las condiciones climáticas lo permitieron (sin bruma o lluvia y estado del mar con Beaufort cuatro o menor). En las aguas del Santuario, se realizó esfuerzo de observación en transecto lineal (principalmente durante los cruceros Pacífico-ERFEN y los tramos de los cruceros restantes dentro del área protegida), y desde punto fijo durante la permanencia en la isla, con un promedio de esfuerzo de cuatro horas diarias. La identificación de los grupos observados fue hecha hasta el mínimo nivel taxonómico posible, generalmente hasta especie. Cuando no se logró identificar la especie, se incluyó la observación en la categoría taxonómica más cercana, con base en algunas características como tamaño, forma y comportamiento (Delphinidae, Ziphiidae, Balaenopteridae y cetáceo no identificado). Dado que la categoría Delphinidae incluyó especies no identificadas con características disímiles (e.g., tamaño grupal), se fraccionó en dos el total de los avistamientos, teniendo como umbral 48 individuos. Este número aparentemente arbitrario, obedece al mejor ajuste obtenido a través del método de distancia, en la estimación de la función de detección de delfínidos en el trabajo de Herrera (2009). De esta manera se buscó que observaciones con características similares en su tamaño grupal estuvieran en un mismo conjunto.
Para el análisis, se consideraron solo las realizadas en esfuerzo y con estado del mar tres o menor según la escala de Beaufort. Debido a que algunos viajes de buceo procedían de la isla Gorgona (2°57.6' N -78°11.4' W), se filtraron las observaciones en el ambito latitudinal 3.53-4.43° N, que corresponden a los extremos sur y norte de las aguas del Santuario. Las tasas de encuentro fueron estimadas como número de grupos por 100 horas de observación. Espacialmente, el área de estudio fue dividida en dos zonas: las aguas del SFF Malpelo (incluidos algunos segmentos de esfuerzo al occidente, producto de los cruceros Pacífico-ERFEN) y el área comprendida entre el continente y la isla, para un total aproximado de 41800 km2 (Figura 1). Para el análisis estacional los datos fueron divididos en trimestres. Para evaluar diferencias estacionales se utilizó la prueba de Kruskal-Wallis y para evaluar diferencias entre zonas (santuario-travesía) se utilizó la prueba de Mann-Whitney. Debido a que T. truncatus fue la especie más abundante en el Santuario y se encontró a lo largo de todo el año, se realizó un ANOVA para evaluar si las tasas de encuentro de esta especie han variado entre años. Todos los análisis se realizaron con ayuda del programa Statistica version 7 (StatSoft, 2004).
RESULTADOS
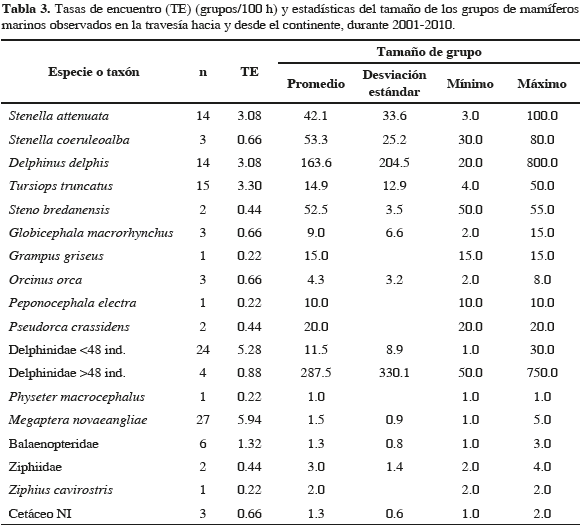
Entre 2001 y 2010, el esfuerzo de búsqueda total fue de 1135.4 horas, de las cuales el 40 % se realizó durante la travesía y el 60 % restante en aguas del SFF Malpelo, tanto desde punto fijo como en desplazamiento (Tabla 1). La Figura 1 presenta los segmentos de esfuerzo realizados en transecto. De los 339 avistamientos del rango latitudinal 3.53-4.43° N, solamente se consideró el 82 % (279) debido a que fueron obtenidos durante condiciones óptimas de muestreo (Beaufort ≤3). Además de las categorías Delphinidae, Ziphiidae, Balaenopteridae y Cetáceo NI se encontraron 15 especies de mamíferos marinos en el área de estudio: Stenella attenuata, S. coeruleoalba, S. longirostris (delfín tornillo o girador), Delphinus delphis, Tursiops truncatus, Steno bredanensis (delfín de dientes rugosos), Peponocephala electra (delfín cabeza de melón), Globicephala macrorhynchus, (calderón de pectoral corta), Orcinus orca (orca), Pseudorca crassidens (falsa orca), Grampus griseus (delfín de Risso o calderón gris), Physeter macrocephalus (cachalote), Ziphius cavirostris (zifio de Cuvier), Megaptera novaeangliae (ballena jorobada) y Zalophus wollebaeki (lobo común de Galápagos). Del total de grupos considerados el 83.2 % (232) se identificó hasta el nivel de especie. Las Tablas 2 y 3 muestran un detalle de los grupos observados en aguas del SFF y en la travesía, y de las estadísticas del tamaño de grupo, mientras que la Figura 2 muestra las posiciones geográficas donde se encontraron.
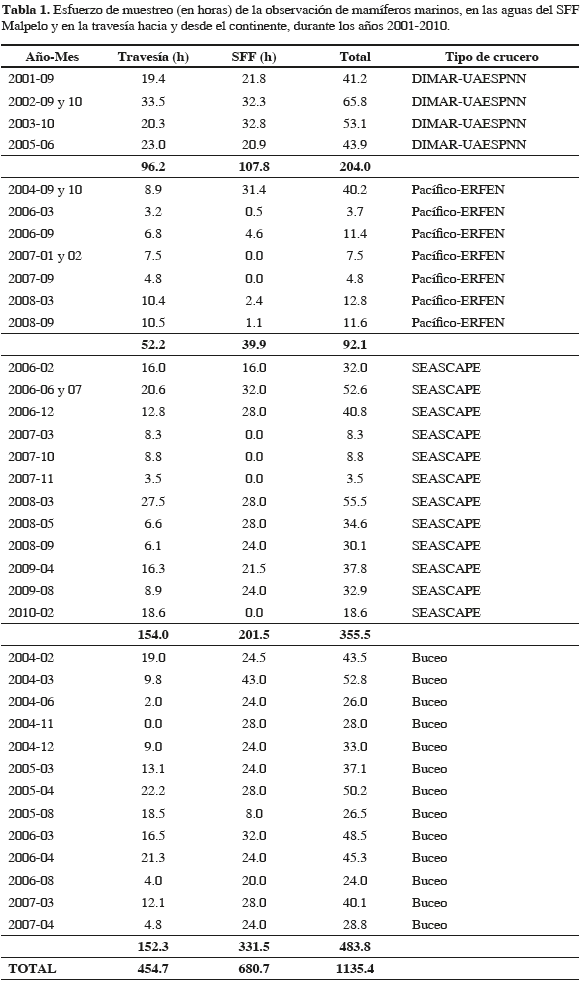
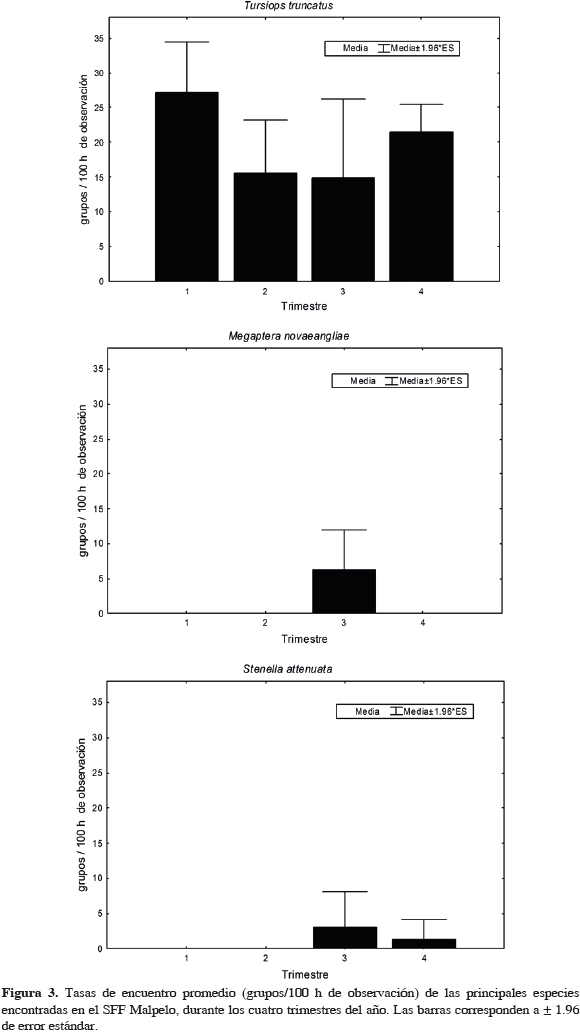
En aguas del Santuario se identificaron siete especies, de las cuales T. truncatus presentó la mayor tasa de encuentro (17.78 grupos/100 h; de aquí en adelante todas las tasas están dadas en las mismas unidades). Su distribución fue muy cercana a la isla, y solo dos grupos se encontraron a distancias mayores a 6 km. El tamaño de grupo promedio fue de 20.8 individuos (+/- 13.8). La segunda especie más importante en los alrededores de Malpelo, fue M. novaeangliae (1.62), y se encontró, en todas las observaciones, a una distancia de la isla menor a 6 km. El tamaño de grupo promedio fue de 1.82 individuos (+/- 0.75). En al menos tres de las once observaciones el grupo incluyó una cría. Los taxa restantes fueron delfínidos, entre los que se destacó S. longirostris, debido a que sólo se observó un solo grupo (en el mes de marzo) que es el único para toda el área y período de estudio; el lobo marino Z. wollebaeki, el cual se encontró solitario dos veces (solo uno durante muestreo con esfuerzo de observación), en las rocas aledañas a la isla en los meses de agosto (2009) y octubre-noviembre (2005); y la categoría Balaenopteridae. En este último caso, las tres observaciones presentaron un solo individuo, una se realizó en el mes octubre (a 1.8 km de la isla) y las otras dos en el período febrero-marzo (a 3 y 10 km, respectivamente), por lo que estás últimas podrían tratarse de alguna especie migratoria proveniente del hemisferio Norte.
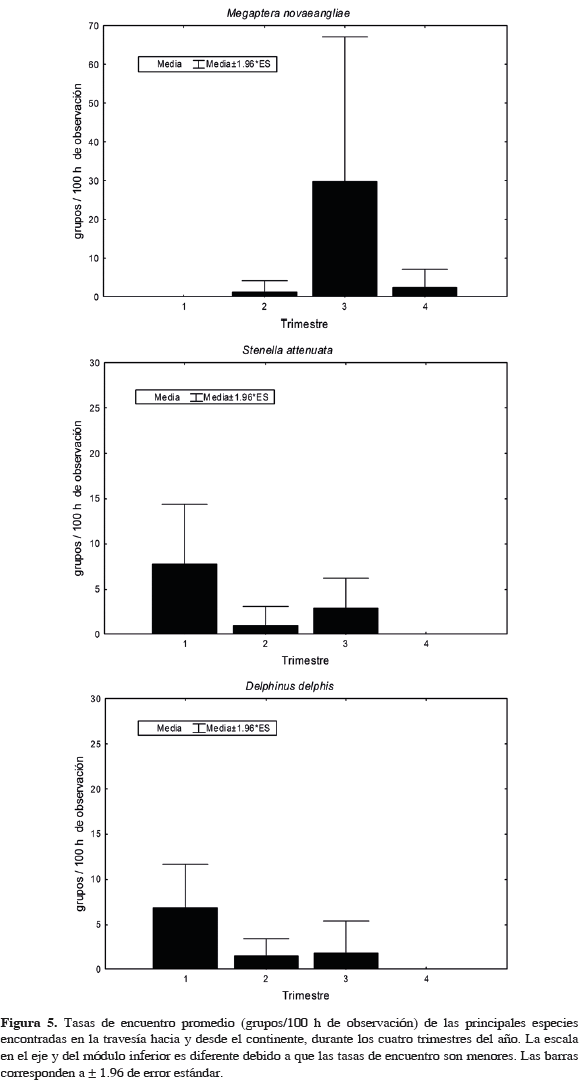
En las aguas intermedias, externas al Santuario, se encontraron 13 especies. Megaptera novaeangliae presentó la mayor tasa de encuentro (5.94), seguida por T. truncatus (3.30), S. attenuata (3.08) y D. delphis (3.08). El tamaño de grupo promedio de M. novaeangliae fue 1.5 (+/- 0.9). Todas las observaciones, excepto una, fueron realizadas sobre la plataforma continental en aguas con profundidades menores a 200 m. El delfín T. truncatus en travesía tuvo un tamaño de grupo promedio menor (14.9 +/- 12.9) que en las aguas del Santuario y osciló entre cuatro y 50 individuos. Todos los grupos, excepto uno, se encontraron en aguas oceánicas con profundidades mayores a 2000 m. Stenella attenuata presentó un tamaño de grupo promedio de 42.1 +/- 33.6 y D. delphis de 163.6 +/- 204.5 (el mayor de las especies identificadas en la travesía y en el Santuario). Delphinus delphis se encontró en aguas oceánicas (profundidad >2000 m) al igual que S. attenuata, aunque de esta última especie algunas observaciones estuvieron presentes en aguas costeras (<200 m) y sobre el talud continental. La categoría Balaenopteridae presentó una tasa de encuentro de 1.32, se observó en los meses de marzo (tres grupos) y junio (tres grupos) con un tamaño promedio de grupo de 1.3 +/- 0.8 y sólo se encontró en aguas oceánicas. Sin discriminar entre zonas (SFF y travesía) la mayor tasa de encuentro fue la de T. truncatus (11.98), seguida por M. novaeangliae (3.35), S. attenuata (1.76), D. delphis (1.32) y S. coeruleoalba (0.62). De otro lado, las menores tasas de encuentro fueron las de S. longirostris, P. electra, G. griseus, Z. cavirostris, P. macrocephalus y Z. wollebaeki todas con 0.09.
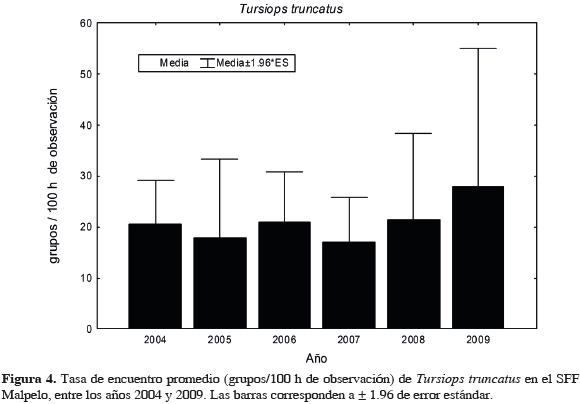
Al evaluar si existen diferencias en las tasas de encuentro entre las aguas del Santuario y las aguas hacia el continente, sólo se encontró significancia para D. delphis (U= 275, n1= 25, n2= 30, p= 0.01) y para T. truncatus (U= 100, n1= 25, n2= 30, p <0.001). El primero fue más abundante en las aguas intermedias y el segundo en las del Santuario. Respecto a la presencia estacional (Tabla 4, que incluye también observaciones fuera de esfuerzo), T. truncatus fue la única especie que estuvo presente a través de todo el año en las aguas de Malpelo, mientras que M. novaeangliae lo estuvo entre junio y noviembre aunque ausente en julio. En las aguas de la travesía (a pesar de haber realizado esfuerzo en enero, durante este mes no se realizaron avistamientos), T. truncatus estuvo a lo largo de todo el año, excepto en algunos meses de mitad de año. Megaptera novaeangliae se encontró en el segundo semestre, pero ausente en agosto, y S. attenuata estuvo en el primer y último cuarto del año (Tabla 4). En el SFF Malpelo la mayor riqueza se encontró entre octubre y noviembre, mientras que en la travesía los meses con mayor riqueza fueron marzo y abril. Al evaluar las diferencias estacionales de las tasas de encuentro de mamíferos marinos del Santuario entre trimestres del año (solo grupos en esfuerzo), solo se encontró significancia para M. novaeangliae (H3, 25= 11.6 p = 0.009) la cual presentó una mayor abundancia en el trimestre julio-septiembre. Aunque en las otras especies no hubo diferencias significativas, la tasa de encuentro de S. attenuata fue mayor en el tercer trimestre, mientras que la de T. truncatus levemente en el primero (Figura 3). La abundancia de T. truncatus no presentó diferencias significativas entre años (F= 0.48, p= 0.84) (Figura 4). En las aguas hacia el continente, M. novaeangliae (H3, 30= 8.04, p= 0.0452) y D. delphis (H3, 25= 8.05, p= 0.0488) presentaron diferencias significativas en la tasa de encuentro entre trimestres. La primera de estas especies fue más abundante en el tercer trimestre (julio-septiembre) y la segunda en el primero (enero-marzo). En el resto de especies no hubo diferencias, sin embargo S. attenuata presentó una tasa de encuentro mayor en el primer trimestre y T. truncatus en el segundo (Figura 5).

DISCUSIÓN
Ninguno de los cruceros realizados fue diseñado exclusivamente para el estudio de cetáceos. Por lo tanto, se parte del supuesto de que el muestreo fue aleatorio y que los avistamientos fueron independientes entre sí. Adicionalmente, hay diferencias marcadas en el tipo de muestreo realizado entre los diferentes cruceros, en lo que concierne al número de observadores y su experiencia, y en general a las características de las embarcaciones utilizadas (altura de la plataforma de observación y velocidad). Sin embargo, la aproximación de las abundancias a través de las tasas de encuentro minimiza el sesgo implícito en el muestreo, y permite hacer una extensa recopilación de información, que considerando las limitaciones de tipo logístico, difícilmente podría llegar a hacerse. También se debe reconocer que no hubo un acercamiento a cada grupo observado para identificar con precisión la especie y para realizar un conteo apropiado del tamaño de grupo. El tipo de muestreo empleado (passing mode según Dawson et al., 2008) aumenta la probabilidad de identificar incorrectamente algunos grupos, o en caso de grupos mixtos, de no detectar una de las especies.
De las 27 especies de mamíferos marinos registradas en aguas del Pacífico colombiano (Flórez-González et al., 2004), el registro directo entre 2001 y 2010, sumado a las observaciones registradas por terceros (Herrera et al., 2007), dan cuenta de la presencia de 11 especies y un género de mamíferos marinos en las aguas del SFF Malpelo. Entre éstas, P. macrocephalus (registrada en 1992), O. orca (registrada en 1986), G. macrorhynchus (registrada entre 2000-2003), P. crassidens (entre 1992- 1999) y Arctocephalus sp. (registrada en 2005, probablemente como A. galapagoensis) fueron registradas por buzos, personal de la Armada u otros investigadores. Para la travesía el total es de 13 especies y para toda el área de estudio 15 especies y un género. De éstas, las únicas especies que no estuvieron en el trabajo de Herrera et al., (2007) fueron Z. cavirostris y P. electra, ambas encontradas en la travesía. La primera ha sido registrada por Herrera (2009) y Palacios et al. (en prensa) en aguas oceánicas al oriente y occidente de la dorsal de Malpelo, con tasas de encuentro muy bajas, mientras que la segunda ha sido registrada al Sur de los 5º N, principalmente fuera de la costa, en el borde de la isóbata de los 1000 m (Palacios et al. en prensa). Considerando la pequeña porción terrestre de la isla Malpelo, la riqueza de mamíferos marinos de sus aguas circundantes es alta (nueve odontocetos, un mysticeto y dos otáridos). La diversidad de mamíferos marinos en otras áreas insulares oceánicas, con un área incluso mayor, son un buen punto de comparación: en el Pacífico tropical, por ejemplo en las islas Hawaii se han registrado 21 especies (Barlow, 2006), 20 en el archipiélago de Galápagos (Palacios y Salazar, 2002) y diez en las islas de la Sociedad y las islas Marquesas (Gannier, 2000, 2002). En el Atlántico, en el archipiélago de las Azores se han registrado 23 especies (Silva et al., 2003; Steiner et al., 2007) y diez en el Índico en la isla de La Reunión (Dulau-Drouot et al., 2008). El otro sistema insular del Pacífico colombiano, aunque con una condición costera (Parque Nacional Natural Gorgona), cuenta con una riqueza algo mayor de mamíferos marinos (quince especies, según Flórez-González y Capella, 2001). Sin embargo, su superficie geográfica y el período de tiempo analizado son considerablemente mayores.
Sin duda, T. truncatus encuentra en las aguas del SFF Malpelo un hábitat propicio para su permanencia. Es la especie más abundante en las aguas del Santuario, se distribuye en aguas cercanas a la isla y está presente a lo largo de todo el año. Según Leatherwood y Reeves (1990), a nivel global se han identificado dos ecotipos de este delfín, uno en aguas pelágicas y otro costero, que habita sobre la plataforma continental. La forma oceánica se caracteriza, entre otros aspectos, por su mayor corpulencia y longitud corporal (Wells y Scott, 2002) y algunos trabajos sugieren que esta forma es genéticamente divergente de la costera (Curry y Smith, 1997). A pesar de no contar con muestras genéticas que permitan corroborar el ecotipo encontrado en Malpelo, su ubicación en los alrededores de una isla oceánica y su mayor tamaño a simple vista, comparado por ejemplo con los que hay en Bahía Málaga, en la misma latitud pero en aguas costeras, sugieren que se trata del ecotipo oceánico. En las islas Galápagos análisis craniométricos indican que éstas se encuentran habitadas por el ecotipo oceánico (Palacios et al., 2004).
Algunas poblaciones de T. truncatus son residentes a lo largo del año, especialmente en áreas costeras (Shane et al., 1986). Por ejemplo, en la costa oeste de la península de La Florida, varias comunidades han permanecido relativamente estables en número y ubicación, sobreponiendo ligeramente sus ámbitos de distribución, durante más de treinta años de observaciones y a través de al menos cuatro generaciones (Wells y Scott, 1999). También existe registro de que su residencia puede ser estable en aguas profundas cercanas a costas de islas oceánicas (Rossbach y Herzing, 1999), lo cual pude ser el caso de Malpelo. La estabilidad de la permanencia puede verse afectada por cambios ambientales como los eventos ENOS (El Niño Oscilación Sur). En Argentina y California se han registro movimientos de varios cientos de kilómetros de algunas poblaciones costeras de T. truncatus, como respuesta al incremento de la temperatura superficial del mar (Wells y Scott, 2002). A pesar de que la CPC está sometida a cambios oceanográficos interanuales, debido a la presencia de eventos El Niño y La Niña, como los que se evidenciaron consecutivamente entre 2006 y 2007, que pueden incluso afectar las abundancias relativas de algunas especies de cetáceos (Herrera, 2009), las tasas de encuentro de T. truncatus en el Santuario permanecieron constantes entre los años evaluados. Igualmente, no hubo diferencias entre trimestres del año, a pesar de la fuerte estacionalidad de la CPC (CCCP, 2002). En la travesía, la forma oceánica de T. truncatus se encontró a lo largo de todo el año (aunque ausente en mayo, julio y agosto), pero en una abundancia significativamente menor a la encontrada en Malpelo. Hasta el momento no se cuenta con elementos de juicio concluyentes (e.g., estudios de fotoidentificación) para saber si los individuos que componen la población de Malpelo son los mismos a través del año y residen permanentemente en la zona, pero es claro que la abundancia poblacional es estable en el tiempo (su tasa de encuentro no fue diferente entre trimestres ni entre años), incluso ante condiciones ambientales adversas como ENOS.
A pesar de su condición migratoria, M. novaeangliae fue la principal especie en la travesía, y se encontró en la segunda mitad del año en aguas costeras de la plataforma. En la recopilación de Palacios et al. (en prensa) fue la especie más frecuentemente avistada en la CPC. Su presencia entre junio y noviembre obedece a que pertenece a la población del Pacífico Sudeste (IWC, 2006), la cual migra durante el invierno austral desde sus zonas de alimentación, localizadas en la costa oeste de la península Antártica (Stevick et al., 2004; Flórez-González et al., 2007) y en el Estrecho de Magallanes (Gibbons et al., 2003; Capella et al., 2008), hacia la zona de reproducción, que se extiende desde el norte de Perú hasta Panamá (Flórez-González et al., 2007), e incluso el sur de Costa Rica (Rassmussen et al., 2007), con áreas de agregación en Colombia, en el golfo de Cupica, golfo de Tribugá, alrededores de Bahía Málaga, en la isla Gorgona y en la zona externa de la ensenada de Tumaco.
Aunque en las zonas de reproducción se le considera costera (Clapham y Mead, 1999), en el Pacífico se reproduce en sitios como las islas Hawaii (Smith et al., 1999; Calambokidis et al., 2001; Johnston et al., 2007), el archipiélago de Revillagigedo (Urbán et al., 2000), y se han observado crías en las islas Galápagos (Merlen, 1995). Sorprendentemente fue la segunda especie más abundante en las aguas de la isla Malpelo, y a pesar de que la zona no presenta sitios poco profundos para la reproducción y la crianza, se observó en varias ocasiones el conjunto de la madre y su cría. Aunque no es posible saber si los nacimientos están teniendo lugar ahí o en otra zona del Pacífico colombiano el registro de su presencia durante los meses de junio a noviembre y su tasa de encuentro, aunque baja comparada con otras zonas reproductivas, permite suponer que el uso que están haciendo del SFF Malpelo es de carácter reproductivo. En algunas zonas donde se observan individuos cerca al continente y en áreas insulares oceánicas, se ha encontrado que las unidades poblacionales son diferentes genéticamente. Es el caso de la población de M. noveangliae del Pacífico Noreste que se reproduce en el archipiélago de Revillagigedo, localizado a 700 km de la costa central de México (Urbán et al., 2000). También es probable que su presencia en Malpelo corresponda a una estadía temporal de grupos en tránsito hacia o desde las áreas de concentración costeras en Colombia (Flórez-González, 1991; Flórez-González et al., 1998), Ecuador (Félix y Haase, 2005), Panamá y/o Costa Rica (Rasmussen et al., 2002). No se puede descartar que algunas de las observaciones de la categoría Balaenopteridae del período febrero-marzo sean de M. novaeangliae. La especie ya ha sido observada durante el primer trimestre del año en cercanías a Cabo Marzo y al golfo de Cupica, lo que sugiere que individuos pertenecientes a la población del Pacífico Noreste también están usando las aguas de la CPC, lo cual es probable, ya que su área de reproducción se sobrepone con la de la población del Pacífico Sureste, entre Panamá y Costa Rica (Rasmussen et al., 2007), a una distancia aproximada de 470 km de la isla Malpelo.
El delfín D. delphis, que junto con S. attenuata presentó la tercera mayor tasa de encuentro, fue más abundante en las aguas oceánicas de la travesía que en el Santuario, encontrándose principalmente entre enero-marzo, período que coincide con el de mayor riqueza en las aguas intermedias. Durante la formación del chorro de viento de Panamá, en el primer trimestre del año, se generan surgencias o afloramientos de aguas profundas ricas en nutrientes, favoreciendo la creación de grandes parches de fitoplancton en el centro de la CPC (Rodríguez-Rubio y Stuardo, 2002). Según Herrera (2009), esta condición favorece la presencia de algunas especies que realizan migraciones regionales desde la zona central del Pacífico Oriental Tropical, justo cuando su distribución se restringe hacia zonas costeras, en el eje de la Contracorriente Ecuatorial del Norte (Reilly, 1990; Reilly y Fiedler, 1994). A pesar de que S. attenuata es una de las especies más abundantes durante el primer semestre del año (Herrera, 2009), solamente se encontró, tanto en el Santuario como en la travesía, en el último cuarto del año. Igualmente, S. coeruleoalaba, que es el delfínido más abundante en la CPC según Gerodette y Palacios (1996) y la especie de odontoceto que presenta la mayor tasa de encuentro según Herrera (2009), se encontró en muy bajas proporciones en toda el área de estudio. Las dos especies de otáridos Z. wollebaeki y Arctocephalus sp. (lobo fino o de dos pelos), no tienen colonias reproductivas en el Pacífico colombiano por lo que no son habitantes permanentes (Flórez-González et al., 2004). Sin embargo, se observan con alguna regularidad y existe evidencia de la presencia de cinco especies de lobos marinos (Palacios et al., 1997; Capella et al., 2002). Estos animales poseen una elevada capacidad de desplazamiento y dispersión poblacional por lo que probablemente provienen desde sus áreas de concentración en las islas Galápagos y las costas de Perú (Capella et al., 2002).
La comunidad de mamíferos marinos de las aguas del SFF Malpelo, representada por diez odontocetos, un mysticeto, y dos otáridos, presenta una alta riqueza, si se consideran las dimensiones del área emergida de la isla. Las características oceanográficas de las aguas circundantes, la disponibilidad de presas y su condición de refugio permiten que brinde soporte para la presencia de diferentes especies de cetáceos entre las que se destaca, por su permanencia a lo largo de todo el año, el delfín nariz de botella. Debido a que no se tuvo una cobertura completa de todas las aguas del Santuario, es necesario a futuro realizar muestreos sistemáticos, que incluya la Dorsal de Malpelo en donde se encuentra el Bajo Bojacá a una profundidad de 281 m, y que probablemente es otro sitio clave de agrupación de mamíferos marinos.
AGRADECIMIENTOS
Los autores agradecen a la Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales (SFF Malpelo), a la Dirección General Marítima, al Centro de Investigaciones Oceanográficas e Hidrográficas del Pacífico (CCCP), a los oficiales, suboficiales y personal civil que integran la tripulación de los buques oceanográficos ARC Providencia y ARC Malpelo, y a la tripulación de la M/N María Patricia, en especial a su Capitán y propietario Harold Botero. El proyecto SEASCAPE y los cruceros científicos al SFF Malpelo contaron con el apoyo económico de Conservación Internacional, el Fondo para la Acción Ambiental y la Niñez, Walton Family Foundation, The Whitley Award, Fondo Patrimonial Exclusivo para el SFF Malpelo, UNESCO y Fundación Malpelo. Para la participación en los cruceros Pacífico-ERFEN de 2007 y 2008 se contó con el apoyo económico de WWF-Colombia y la Fundación Yubarta. Elizabeth Hernández, Isabel Ávila, y Wilfredo Henao participaron como observadores en alguno de los cruceros.
BIBLIOGRAFÍA
1 Andrade, C. A. 1992. Movimiento geostrófico en el Pacífico colombiano. Bol. Cient. CIOH, 12: 23-38. [ Links ]
2 Barlow, J. 2006. Cetacean abundance in Hawaiian waters estimated from a summer/fall survey in 2002. Mar. Mammal Sci., 22 (2): 446-464. [ Links ]
3 Buckland, S. T., D. R. Anderson, K. P. Burnham, J. L. Laake, D. L. Borchers y L. Thomas. 2001. Introduction to Distance Sampling. Oxford University Press, Oxford. 432 p. [ Links ]
4 Calambokidis, J., G. Steiger, J. Straley, L. Herman, S. Cerchio, D. Salden, J. Urbán, J. Jacobsen, O. von Ziegesar, K. Balcomb, C. Gabriele, M. Dahlheim, S. Uchida, G. Ellis, Y. Miyamura, P. Ladrón de Guevara, M. Yamaguchi, F. Sato, S. Mizroch, L. Schlender, K. Rasmussen, J. Barlow y T. Quinn II. 2001. Movements and population structure of humpback whales in the North Pacific. Mar. Mammal Sci., 17: 769-794. [ Links ]
5 Capella, J., L. Flórez-González, P. Falk y D. Palacios. 2002. Regular appearance of Otariid Pinnipeds along the Colombian Pacific coast. Aquatic Mammals., 28 (1): 67-72. [ Links ]
6 Capella, J. J., J. Gibbons, L. Flórez-González, M. Llano, C. Valladares, V. Sabaj y Y. Vilina. 2008. Migratory round-trip of individually identified humpback whales at the Strait of Magellan: clues on transit times and phylopatry to destinations. Rev. Chil. Hist. Nat., 81: 547-560. [ Links ]
7 Centro Control Contaminación Pacífico CCCP. 2002. Compilación oceanográfica de la Cuenca Pacífica Colombiana. Serie Publicaciones Especiales Vol. 1, CCP, San Andrés de Tumaco. 107 p. [ Links ]
8 Chelton, D. B., M. H. Freilich y S. K. Esbensen. 2000. Satellite observations of the wind jets off the Pacific Coast of Central America. Part II: Relationships and dynamical considerations. Mon. Weather Rev., 128: 2019-2043. [ Links ]
9 Clapham, P. y J. Mead. 1999. Megaptera novaeangliae. Mammalian Species, 604: 1-9. [ Links ]
10 Curry, B. E. y J. Smith. 1997. Phylogeographic structure of the bottlenose Dolphin (Tursiops truncatus): stock identification and implications for management. 227-247. En: Dizon, A. E., S. J. Chivers y W. F. Perrin (Eds.). Molecular genetics of marine mammals. Special publication, Society for Marine Mammalogy, Lawrence. 412 p. [ Links ]
11 Dawson, S., P. Wade, E. Slooten y J. Barlow. 2008. Design and field methods for sighting surveys of cetaceans in coastal and riverine habitats. Mammal Rev., 38 (1): 19-49. [ Links ]
12 Devis-Morales, A., W. Schneider, R. A. Montoya-Sánchez y E. Rodríguez-Rubio. 2008. Monsoon-like winds reverse oceanic circulation in the Panama Bight. Geophys. Res. Lett., 35 L20607: 1-6. [ Links ]
13 Dulau-Drouot, V., V. Boucaud y B. Rota. 2008. Cetacean diversity off La Reunion Island (France). J. Mar. Biol. Assoc. UK, 88 (6): 1263-1271. [ Links ]
14 Félix, F. y B. Haase. 2005. Distribution of humpback whales along the coast of Ecuador and management implications. J. Cetacean Res. Manage., 7 (1): 21-31. [ Links ]
15 Flórez-González, L. 1991. Humpback whales Megaptera novaeangliae in the Gorgona Island, Colombian Pacific breeding waters: population and pod characteristics. Mem. Queensland Mus., 30 (2): 291-295. [ Links ]
16 Flórez-González, L. y J. Capella. 1995. Mamíferos acuáticos de Colombia. Una revisión y nuevas observaciones sobre su presencia, estado del conocimiento y conservación. Inf. Mus. Mar (39), 29 p. [ Links ]
17 Flórez-González, L. y J. Capella. 2001. Mamíferos marinos locales y regionales. 133-140. En: Barrios, L. y M. López-Victoria (Eds.). Gorgona marina: Contribución al conocimiento de una isla única. Serie Publicaciones Especiales No 7, INVEMAR, Santa Marta. 160 p. [ Links ]
18 Flórez-González, L., J. Capella, B. Haase, G. Bravo, F. Félix y T. Gerrodette. 1998. Changes in winter destinations and northernmost record of Southeastern Pacific humpback whales. Mar. Mammal Sci., 14 (1): 189-196. [ Links ]
19 Flórez-González, L., J. Capella y P. Falk. 2004. Guía de campo de los mamíferos acuáticos de Colombia. Tercera edición, Editorial Sepia Ltda., Cali. 124 p. [ Links ]
20 Flórez-González, L., I. C. Ávila, J. Capella, P. Falk, F. Félix, J. Gibbons, H. Guzmán, B. Haase, J. Herrera, V. Peña, L. Santillán, I. C. Tobón y K. Van Warebeek. 2007. Estrategia para la conservación de la ballena jorobada del Pacífico Sudeste. Lineamientos de un plan de acción regional e iniciativas nacionales, Fundación Yubarta. Cali. 106 p. [ Links ]
21 Forney, K. A. 2002. Surveys. 1203-1205. En: Perrin, W. F., B. Würsig y J. G. M. Thewissen (Eds.). Encyclopedia of marine mammals. Academic Press, San Diego. 1414 p. [ Links ]
22 Gannier, A. 2000. Distribution of cetaceans off the Society Islands (French Polynesia) as obtained from dedicated surveys. Aquatic Mammals, 26 (2): 111-126. [ Links ]
23 Gannier, A. 2002. Cetaceans of the Marquesas Islands (French Polynesia): distribution and relative abundance as obtained from a small boat dedicated survey. Aquatic Mammals, 28 (2): 198-210. [ Links ]
24 Gerrodette, T y D. Palacios. 1996. Estimates of cetacean abundance in EEZ. Waters of the eastern Tropical Pacific. Administrative Report LJ 96-10, NMFS-SFSC, La Jolla, Estados Unidos. 28 p. [ Links ]
25 Gibbons, J., J. J. Capella y C. Valladares. 2003. Rediscovery of a humpback whale, Megaptera novaeangliae, feeding ground in the Straits of Magellan, Chile. J. Cetacean Res. Manage., 5 (2): 203-208. [ Links ]
26 Herrera, J. C. 2009. Distribución y abundancia relativa de cetáceos en el Pacífico colombiano y su relación con las condiciones oceanográficas. Tesis de Maestría, Universidad del Valle, Cali. 152 p. [ Links ]
27 Herrera, J. C., I. Ávila, P. Falk, G. Soler, C. García, I. Tobón y J. Capella. 2007. Los mamíferos marinos en el Santuario de Fauna y Flora Malpelo y Aguas hacia el Continente, Pacífico colombiano. 113-130. En: DIMAR-CCCP y UAESPNN (Eds.). Santuario de Fauna y Flora Malpelo: descubrimiento en marcha. DIMAR, Bogotá. 142 p. [ Links ]
28 International Whaling Commission. 2006. Report of the scientific committee. Annex H. Report of the Sub-committee on the other Southern hemisphere whale stocks. J. Cetacean Res. Manage., (Supl.) 8: 151-70. [ Links ]
29 Johnston, D. W., M. E. Chapla, L. E. Williams y D. K. Mattila. 2007. Identification of humpback whale Megaptera novaeangliae wintering habitat in the Northwestern Hawaiian Island using spatial habitat modeling. Endang. Species Res., 3: 249-257. [ Links ]
30 Leatherwood, S. y R. Reeves. 1990. The bottlenose dolphin. Academic Press, San Diego. 653 p. [ Links ]
31 López-Victoria, M. y D. M. Rozo. 2006. Model-based geomorphology of Malpelo Island and spatial distribution of breeding seabirds. Bol. Invest. Mar. Cost., 35:111-131. [ Links ]
32 Merlen, G. 1995. A field guide to the marine mammals of Galápagos. Instituto Nacional de Pesca, Guayaquil. 130 p. [ Links ]
33 Palacios, D. M. y S. Salazar. 2002. Cetáceos. 291-304. En: Danulat, E. y G. J. Edgar (Eds.). Reserva marina de Galápagos: Línea de base de la biodiversidad. Fundación Charles Darwin/Servicio del Parque Nacional Galápagos, Santa Cruz, Galápagos. 484 p. [ Links ]
34 Palacios, D., F. Félix, L. Flórez-González, J. Capella, D. Chiluiza y B. Haase. 1997. Sightings of Galápagos sea lions (Zalophus californianus wollebaeki) on the coasts of Colombia and Ecuador. Mammalia, 61 (1): 114-116. [ Links ]
35 Palacios, D. M., S. K. Salazar y D. Day. 2004. Cetacean remains and strandings in the Galápagos Islands, 1923-2003. Lat. Am. J. Aquat. Mamm., 3(2):127-150. [ Links ]
36 Palacios, D. M., J. C. Herrera, T. Gerrodette, C. García, G. A. Soler, I. C. Ávila, S. Bessudo, E. Hernández, F. Trujillo y L. Flórez-González. En prensa. Cetacean distribution and relative abundance in Colombia's Pacific EEZ from survey cruises and platforms of opportunity. J. Cetacean Res. Manag. [ Links ]
37 Rasmussen, K., J. Calambokidis y G. Steiger. 2002. Humpback whales and other marine mammals of Costa Rica and surrounding waters, 1996-2002. Report of the Oceanic Society 2002. Field season in cooperation with Elderhostel volunteers, Oceanic Society Expeditions, Cascadian Research. Olympia, Estados Unidos. 2221 p. [ Links ]
38 Rasmussen, K., D. Palacios, J. Calambokidis, M. Saborio, L. Dalla Rosa, E. Secchi, G. H. Steiger, J. M. Allen y G. Stone. 2007. Southern Hemisphere humpback whales wintering off Central America: insights from water temperature into the longest mammalian migration. Biol. Lett., 3 (3): 302-305. [ Links ]
39 Reilly, S. B. 1990. Seasonal changes in distribution and habitat differences among dolphins in the eastern tropical Pacific. Mar. Ecol. Prog. Ser., 66: 1-11. [ Links ]
40 Reilly, S. B. y P. C. Fiedler. 1994. Interannual variability of dolphin habitats in the eastern tropical Pacific. I: Research vessel surveys, 1986-1990. Fish. B-NOAA, 92: 434-450. [ Links ]
41 Rodríguez-Rubio, E. y J. Stuardo. 2002. Variability of photosynthetic pigments in the Colombian Pacific Ocean and its relationship with the wind field using ADEOS-I data. Proc. Indian Acad. Sci. (Earth Planet. Sci.), 111 (3): 227-236. [ Links ]
42 Rodríguez-Rubio, E., W. Schneider y R. Abarca del Rio. 2003. On the seasonal circulation within the Panama Bight derived from satellite observations of wind, altimetry and sea surface temperature. Geophys. Res. Lett., 30 (7): 1410. [ Links ]
43 Rossbach, K. y D. Herzing. 1999. Inshore and offshore bottlenose dolphins (Tursiops truncatus) communities distinguished by association patterns near Grand Bahama Island, Bahamas. Can. J. Zool., 77: 581-592. [ Links ]
44 Rueda, J. G., E. Rodríguez-Rubio y J. R. Ortiz. 2007. Caracterización espacio temporal del campo de vientos superficiales del Pacífico colombiano y el Gofo de Panamá a partir de sensores remotos y datos in situ. Bol. Cient. CCCP, 14: 49-68. [ Links ]
45 Shane, S., R. Wells y B. Wursig. 1986. Ecology, behavior and social organization of the bottlenose dolphin: a review. Mar. Mammal Sci., 2: 34-63. [ Links ]
46 Silva, M. A., R. Prieto, S. Magalhães, R. Cabechinas, A. Cruz, J. M. Goncalves y R. S. Santos. 2003. Occurrence and distribution of cetaceans in the waters around the Azores (Portugal), Summer and Autumn 1999-2000. Aquatic Mammals, 29 (1): 77-83. [ Links ]
47 Smith, T., J. Allen, P. Clapham, P. Hammond, S. Katona, F. Larsen, J. Lien, D. Mattila, P. Palsboll, J. Sigurjónsson, P. Stevick y N. Oien. 1999. An ocean-wide mark-recapture study of the North Atlantic humpback whale (Megaptera novaeangliae). Mar. Mammal Sci., 15: 1-32. [ Links ]
48 Steiner, L., M. A. Silva, J. Zereba y M. J. Leal. 2007. Bryde's Whales, Balaenoptera edeni, observed in the Azores: a new species record for the region. Mar. Biod. Rec., 1: 1-6. [ Links ]
49 Stevenson, M. 1970. Circulation in the Panama Bight. J. Geophys. Res., 75, 659-672. [ Links ]
50 Stevick, P. T., A. Aguayo, J. Allen, I. C. Avila, J. Capella, C. Castro, K. Chater, L. Dalla Rosa, M. Engel, F. Felix, L. Florez-Gonzalez, A. Freitas, B. Haase, M. Llano, L. Lodi, E. Munoz, C. Olavarria, E. Secchi, M. Scheidat y S. Siciliano. 2004. Migrations of individually identified humpback whales between the Antarctic Peninsula and South America. J. Cetacean Res. Manag., 6 (2): 109-113. [ Links ]
51 Urbán, J., A. Jaramillo, A. Aguayo, P. Ladrón de Guevara, M. Salinas, C. Álvarez, L. Medrano, J. K. Jacobsen, K. C. Balcomb, D. E. Claridge, J. Calambokidis, G. H. Steiger, J. M. Straley, O. von Ziegesar, J. M. Waite, S. Mizroch, M. E. Dalheim, J. D. Darling y C. S. Baker. 2000. Migratory destinations of humpback whales wintering in the Mexican Pacific. J. Cetacean Res. Manag., 2 (2): 101-110. [ Links ]
52 Vidal, O. 1990. Lista de los mamíferos acuáticos de Colombia. Inf. Mus. Mar (37), 18 p. [ Links ]
53 Wade, P. y T. Gerrodette. 1993. Estimates of cetacean abundance and distribution in the Eastern Tropical Pacific. Report of the International Whaling Commission, 43: 477-493. [ Links ]
54 Wells, R. S. y M. D. Scott. 1999. Bottlenose dolphin-Tursiops truncatus (Montagu, 1821). 137-182. En: Ridgway, S. H. y R. J. Harrison (Eds.). Handbook of marine mammals. Volume 6: The second book of dolphins and porpoises. Academic Press, Londres. 486 p. [ Links ]
55 Wells, R. y M. Scott. 2002. Bottlenose dolphins Tursiops truncatus and T. aduncus. 122-128. En: Perrin, W., B. Wursig y J. G. Thewissen (Eds.), Encyclopedia of Marine Mammals. Academic Press, San Diego. 1414 p. [ Links ]
56 Wooster, W. S. 1959. Oceanographic observations in the Panamá Bight, Asoky Expedition 1941. Bull. Am. Mus. Nat. Hist., 118: 115-151. [ Links ]
57 Wyrtki, K. 1966. Oceanography of the Eastern Equatorial Pacific Ocean. Oceanogr. Mar. Biol. Ann. Rev. 4: 33-68 p. [ Links ]
FECHA DE RECEPCIÓN: 01/10/2010 FECHA DE ACEPTACIÓN: 12/10/2011
