










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.40 supl.1 Santa Marta Dec. 2011
VERTICAL AND HORIZONTAL MOVEMENTS OF THE SCALLOPED HAMMERHEAD SHARK (SPHYRNA LEWINI) AROUND MALPELO AND COCOS ISLANDS (TROPICAL EASTERN PACIFIC) USING SATELLITE TELEMETRY
MOVIMIENTOS VERTICALES Y HORIZONTALES DEL TIBURÓN MARTILLO (SPHYRNA LEWINI) ALREDEDOR DE LAS ISLAS MALPELO Y COCOS (PACÍFICO ORIENTAL TROPICAL) USANDO TELEMETRÍA SATELITAL
Sandra Bessudo1, Germán A. Soler1 , Peter A. Klimley2, James Ketchum2, Randall Arauz3 , Alex Hearn2,4, Adriana Guzmán1 and Beatriz Calmettes5
1 Fundación Malpelo y Otros Ecosistemas Marinos, Carrera 7 No 32-33 Piso 27, Bogotá, Colombia. sandrabessudo@presidencia.gov.co (S.B.), gsoler@fundacionmalpelo.org (G.A.S.), aguzman@fundacionmalpelo.org (A.G.).
2 University of California, Davis, One Shields Avenue, Davis, CA 95616, United States of America apklimley@ucdavis.edu (P.A.K.), jtketchum@ucdavis.edu (J.K.), arhearn@ucdavis.edu (A.H.).
3 PRETOMA, 1203-1100, Tibás, San José, Costa Rica. rarauz@pretoma.org.
4 Charles Darwin Foundation, Santa Cruz, Galápagos, Ecuador.
5 Collecte Localisation Satellite-CLS Modélisation des Ecosystèmes Marins et Monitoring par Satellites Division océanographie spatiale 8-10 rue Hermès 31520 Ramonville St Agne, France. bcalmettes@cls.fr.
ABSTRACT
Understanding distribution ranges and the daily movement patterns of pelagic fishes are key aspects for the establishment and planning of protected areas for their conservation. In this study the vertical and horizontal movements of scalloped hammerhead sharks, Sphyrna lewini, were recorded in Malpelo and Cocos Island using satellite telemetry. Nine sharks were tagged with satellite transmitters during March 2006, 2007 and 2008 at Malpelo Island, and three hammerhead sharks were tagged at Cocos Island in June 2008. The sharks moved between islands in the Tropical Eastern Pacific and made regional movements from Malpelo to the south of Cocos and around the Malpelo ridge. When away from the island, sharks made infrequent nocturnal short dives down to 1000 m where temperatures were as low as 4 °C. For a shark that was tracked for five months, results indicated significant differences between the time spent at different depth ranges, as well as an interaction between depth, water layers, and the time of the day. The shark swam mostly at surface waters (0-10 m) during the cold water season, and spent more time at the deepest depth range (>100 m) during nigh time. Further long-term studies on shark movement patterns are required for a better management of this highly mobile and vulnerable species.
KEY WORDS: Satellite telemetry, Vertical and horizontal movements, Sphyrna lewini, Malpelo, Cocos.
RESUMEN
El conocimiento de los ámbitos de distribución y los patrones diarios de actividad de peces pelágicos son aspectos clave para el establecimiento y planificación de áreas protegidas para su conservación. En este estudio los movimientos verticales y horizontales de los tiburones martillo, Sphyrna lewini, fueron registrados utilizando telemetría satelital en las islas de Malpelo y Cocos, en el Pacífico Oriental Tropical. Nueve tiburones fueron marcados con transmisores satelitales en marzo de 2006, 2007 y 2008 en la isla Malpelo, y tres tiburones martillo fueron marcados en la isla de Cocos en junio de 2008. Los tiburones se movieron entre las islas del Pacifico Oriental Tropical y realizaron movimientos regionales desde Malpelo hacia el sur de Cocos y sobre la cordillera Dorsal de Malpelo. Lejos de las islas, los tiburones realizaron buceos nocturnos poco frecuentes y cortos, hasta 1000 m de profundidad, donde las temperaturas alcanzaron hasta 4 °C. Para el tiburón que se siguió por cinco meses, los resultados indicaron diferencias significativas entre el tiempo que invirtió a diferentes ámbitos de profundidad, como también una interacción entre la profundidad, las capas de agua y el momento del día. El tiburón nadó principalmente en aguas someras (0-10 m) durante el periodo de aguas frías, e invirtió más tiempo en ámbitos de profundidad mayores (>100 m) durante la noche. Futuros estudios a largo plazo son necesarios para un mejor manejo de esta especie altamente móvil y vulnerable.
PALABRAS CLAVE: Telemetría satelital, Movimientos verticales y horizontales, Sphyrna lewini, Malpelo, Cocos.
INTRODUCTION
Apex predators have been recognized as key species within a system due to their ability to regulate species abundance, distribution and diversity, by controlling prey densities (Sergio et al., 2006) and increasing competition (Glen et al., 2007). In marine environments, apex predators are of crucial importance to maintain oceans healthy (Griffin et al., 2008). Furthermore, some marine apex predators such as sharks help maintain the complex ecosystems by providing food sources for scavengers (Sergio et al., 2006) and remove weak individuals from prey populations (Temple, 1987; Griffin et al., 2008). Thus, efforts to reduce the number of sharks captured in commercial fisheries through improved shark management are of crucial importance in conservation.
The scalloped hammerhead shark (Sphyrna lewini) is a top marine predator with a worldwide distribution. However, populations are estimated to have declined by more than 75 percent in the past few decades (Baum et al., 2003; IUCN, 2007; Camhi et al., 2008). Scalloped hammerhead sharks are frequently captured in large numbers in the bycatch of industrial as well as small-scale commercial fisheries in all three major oceans (Stevens, 2000; IUCN, 2007). Their meat, liver oil, skin and fins are well commercialized in the international trade market (Rose, 1996; FAO, 2000; Simpfendorfer and Heupel, 2004; Chapman et al., 2005; IUCN, 2007). Its late onset of sexual maturity, low fecundity and low natural mortality, result in a low intrinsic rate of population increase (Ferrari and Ferrari, 2001), making it more vulnerable to overexploitation (Camhi, 1998; Watts, 2001; Newmark and Santos-Acevedo, 2002; Baum et al., 2003; Jennings et al., 2005). For these reasons, S. lewini has been designated as globally Endangered by the IUCN Red List (IUCN, 2010).
Large aggregations of hammerhead sharks around Malpelo Island and Cocos Island have been documented during some months of the year, followed by a period (April- May) with a reduction of their densities, possibly due to extensive migrations (Sibajo- Cordero, 2008; Bessudo et al., 2011). Long-distance migrations of oceanic species have been difficult to study. However, in the last decades satellite telemetry has become a very useful technique to understand the behavior and movement patterns of some pelagic fish species (Sedberry and Loefer, 2001; Wilson et al., 2005), including sharks (Sciarrotta and Nelson, 1977; Heithaus et al., 2007; Pade et al., 2009).
A better understanding of the temporal and spatial scales at which shark species move is of paramount importance to ecologists and conservationists. This information could give insights on habitat use, key sites occupied (Schneider, 1994), as well as other essential information for the effective management and design of marine reserves that could be used to optimize the conservation of these highly mobile predators (Garla et al., 2006). Furthermore, shark movement patterns could be linked to their potential effect on prey and to the possibility that they encounter fisheries outside protected areas (Heithaus et al., 2007). In this paper we present information on the long distance movements of scalloped hammerhead sharks tagged at Malpelo Island and at Cocos Island using satellite tags.
STUDY AREA
Malpelo Island (4°00' N and 81°36'30" W) is located in the Eastern Pacific Ocean, 500 km from the port of Buenaventura in the Colombian Pacific coast (Figure 1). This 1.2 km2 island is surrounded by eleven pinnacles with its highest point at 300 m above sea level (López-Victoria and Rozo, 2006). Oceanic waters surrounding Malpelo Island are influenced seasonally by four currents: the North Equatorial Countercurrent, the South Equatorial Countercurrent, the Colombia Current, and the Panama Cyclonic Current (Rodríguez-Rubio et al., 2007). The annual mixing period of these currents depends on the migration of the Intertropical Convergence Zone (Rodríguez-Rubio et al., 2003) and the degree of mixing depends upon longer term cycles such as El-Niño Southern Oscillation (ENSO) (Devis-Morales et al., 2008). Interactions between these currents result in two distinct seasons: a cold and a warm water season. During the cold water season, when temperatures are typically of 25 °C on the surface and below the thermocline as low as 14 °C, from January through April, waters surrounding the island are full of nutrients, therefore highly productive (Rodríguez-Rubio et al., 2003).
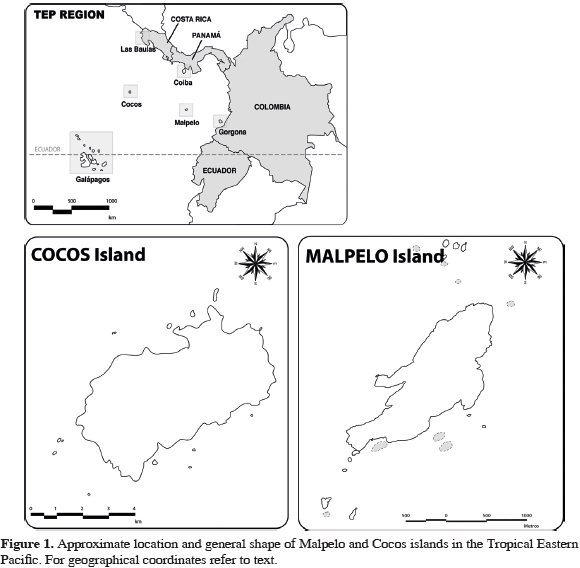
Furthermore, during this cold water season the thermocline is found fairly shallow between 10-20 m. During the warm water season, from May until the end of December, average water temperature is around 27 °C (Rodríguez-Rubio et al., 2003). Cocos Island (5°31' N and 87°04' W), located 500 km southwest of the Costa Rican coast and 627 km northeast of Malpelo Island in the Eastern Tropical Pacific, has a total perimeter of 23.3 km and an area of 47 km2 (Castillo et al., 1988). This volcanic island is the only summit of the submerged Cocos Range that reaches the surface and it is associated with the Galápagos hotspot (Castillo et al., 1988). The North Equatorial Countercurrent does not reach the island until around May. Because of this, an anticyclonic circulation is observed during the first trimester of the year at the southern part of the island. Then, during the second semester of the year when the North Equatorial Countercurrent appears, the same turns are observed but with different levels of current intensity. During the third and fourth trimesters of the year, current direction and intensity are similar, given that the North Equatorial Countercurrent is fully established, with its maximum flow to the east. Mean water temperature range between 26.8 and 28.4 °C, water being warmer during January to June and cooler during July to December (Lizano, 2008).
MATERIALS AND METHODS
Satellite telemetry
Satellite tags were affixed to sharks by free divers and SCUBA divers by inserting a stainless steel barb into the dorsal musculature of the sharks. Each tag was attached with a tether to the barb, which was placed in the inset of the tip of a spear gun. Satellite tags used in this study were pop-up archival transmitting (PAT2, PAT4 and Mk10 Wildlife Computers, Inc, Redmond, Washington, USA; Table 1) that release at a preset interval and transmit to the Argos satellites. The PATs included a battery, a microprocessor, a Random Access Memory for data storage, various environmental sensors, and a radio transmitter. Sensors measure bluegreen irradiance (~490 nm), depth, and water temperature. Each channel records measurements at regular intervals, and these measurements are stored in the tag's onboard memory. We programmed our PAT tags with a two minute interval, which permits a time-at-liberty of 12 months before filling up the memory. These tags are externally attached to the animal, using a monofilament tether severed by a cutting mechanism if the animal dives below 1500 m, which will result in the implosion of the tag's housing. After a predefined interval, current flows through a pin causing it to corrode, and the tag releases from the shark and floats to the surface.
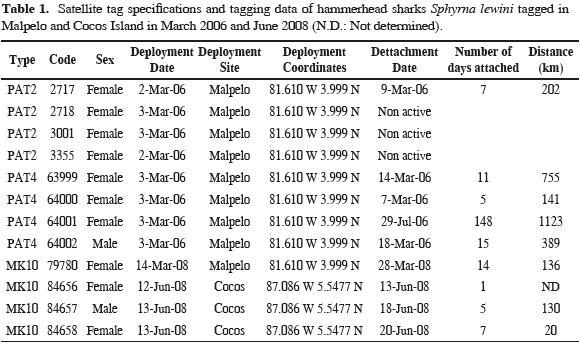
Upon retrieval of the data, through detachment of the tag, positioning the fish was done a posteriori using records of light levels at depth. The simplest method, used early in the field of marine wildlife tracking, relies on light thresholding to detect timings of sunrise and sunset events (Welch and Eveson, 1999). Alternatively, when all the light records are available, a fish can be located by finding the coordinates that best predict the observed light curve (Ekstrom, 2004). This is often called “direct light-based geolocation", in that it only uses light levels or sunrise/sunset times to estimate a daily position. Manufacturers usually provide these position estimates. For instance, Wildlife Computers, Inc. provides the software suite to decode Argos transmitted data and infer positions from the light curves. This has the advantage of simplicity, but can be error-prone, especially around the equinoxes where latitude becomes indeterminate. This well known error pattern occurs as the duration of the day does not vary much with latitude, and thus locations are highly inaccurate at this time of the year. Longitude however is not affected. To solve for this and improve geolocation accuracy, one can include additional data. This can take the form of a movement model (i.e. knowledge that the animal moves at a certain speed), or oceanographic information (e.g., satellite-derived sea surface temperature to be matched with in situ, tag-derived temperature records). To improve the geolocation quality, we applied a Kalman filter based procedure (Royer and Lutcavage, 2009). This model considers the animal behavior (mean speed by day) and oceanographic data: sea surface temperature measured by satellite compared to the sea surface temperature measured by the tag and bathymetry compared to the maximum depth measured by the tag. Instead of using the full irradiance model, we use the following astronomical equations relating longitude and latitude to a sunrise and sunset times, in minutes after 0:00 GMT.
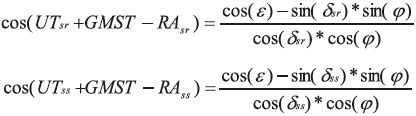
Where φ is the animal's latitude and δsr,ss and RAsr,ss are the solar declination and right angle at sunrise and sunset. ε is the sun's altitude at twilight and UT Universal Time. The function relating locations to sunrise and sunset is given by:
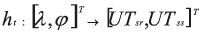
Where λ is the animal longitude and T means transpose operation. The model propagating the fish's position at time t is assumed to be a random walk, parameterized by a diffusion matrix and a bias:
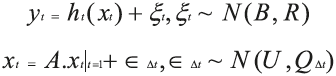
Where A is an identity matrix, U = [u, v] contains the meridional and zonal mean movement and Q is a diffusion matrix (see Royer and Lutcavage, 2009 for a complete description of the method).
Statistical analysis
In order to test whether the shark identified with the code 64001 preferred certain swimming depths (<10 m, 10-50 m, 50-100 m, >100 m), between water season (cold, warm) and time of the day (day= 06:00 to 18:00, night= 18:00 to 06:00), a three-way analysis of variance (ANOVA) with Tukey-Kramer post hoc tests (α= 0.05) was performed (JMP program, version 3.0). These analyses were only made for shark 64001 because it was the individual that retained the tag for the longest time (148 days; for sharks's codes and abbreviations refer to Table 1).
RESULTS
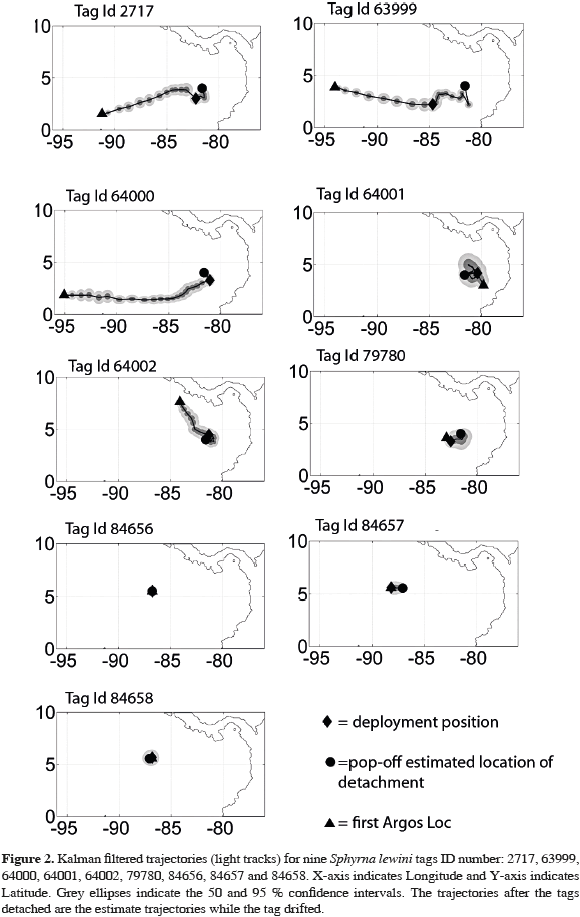
We tagged nine hammerhead sharks at Malpelo Island and three at Cocos Island. Three of the tags failed to communicate with the satellite. The remaining tags all released prematurely from the sharks between one and 148 days of being implanted (mean retention time= 23.6 days; SD= 46.9; N= 9) (Table 1). It is worth noting that for the shark that was tracked for 148 days, not all records from the tag were received, presumably due to transmission problems with the ARGOS satellite. Hence, we do not have continuous data on location, dive profiles, time spent at the different depths, or temperature bins for this individual. For the tag that remained on the shark for a day, distance traveled could not be estimated. Most of the tagged sharks at Malpelo moved in a linear fashion, although in different directions (2717, 63999, 64000 and 79780; Figure 2). The other two sharks tagged at Malpelo (64001 and 64002) did not display a clear pattern, moving east, north and west, indistinctly (Figure 2). Of the sharks tagged at Cocos, one shark (84657) moved west along a seamount around Cocos Island during five days, while the other (84658) remained near the vicinity of Cocos Island for seven days (Figure 2).
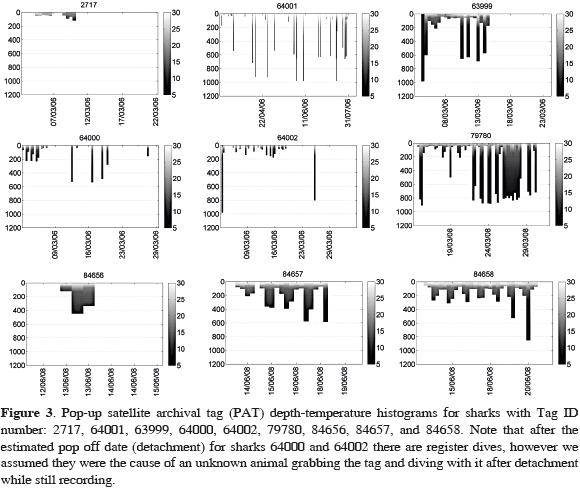
Hammerhead sharks tagged during this study showed broadly similar patterns of vertical movements, swimming primarily within the mixed layer (<100 m, >18 °C) and making short yo-yo like dives below the thermocline to depths between 100-1000 m, where water temperatures were as low as 4 °C (Figure 3).
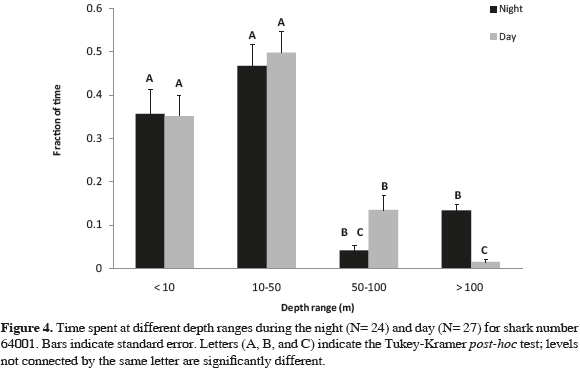
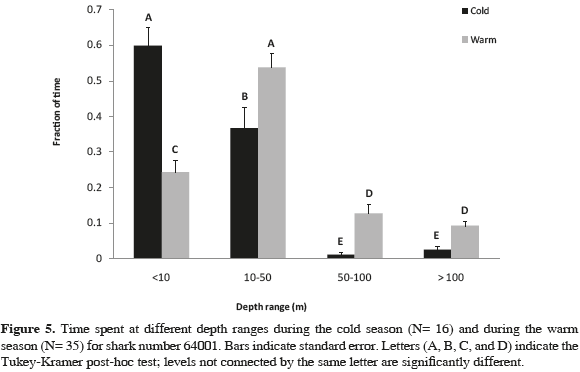
The analysis of variance (ANOVA) for data obtained from shark 64001 showed highly significant differences between the time spent at different depth ranges (p <0.001, F= 91.9, df= 3), as well as between the time spent at different depths and the water season (p <0.001, F= 20.5, df= 3), the time spent at different depths and the time of the day (p <0.001, F= 5.9, df =3), and the time spent at different depths, water season, and time of the day (p= 0.016, F= 3.5, df= 3). Tukey- Kramer test indicated significant differences in the time spent at all depth ranges comparing water seasons: this shark swam mostly at surface waters (0-10 m) during the cold water season compared to the warm water season (55 and 26 % of the time, respectively). During the later season the shark swam deeper (Figure 5), when the thermocline was deeper at Malpelo Island and the sea surface temperature was higher (around 28 °C). Significant differences were also found between the time spent at the deepest depth range (>100 m), when comparing day time with night time (Figure 4). Most of the dives registered below 100 m were almost exclusively done by the shark at night, presumably away from Malpelo Island, taken in consideration that depths below 150 m are found only further away from the island.
DISCUSSION
No clear movement patterns were observed from the tagged sharks, possibly due to the small sample size of this study and the low mean retention time (23.6 days). While sharks that were tracked for more than 11 days made round trips and remained close to the site where they were tagged, sharks tracked between five to 11 days moved with directionality suggesting high individual variability. Complex shark movements similar to those found by this study have also been observed by tagged hammerhead sharks at Galapagos Islands (Ketchum et al., 2009) and by tiger sharks in Australia (Heithaus et al., 2007). Possible explanations for this movement complexity could be the diverse habitats present, oceanographic dynamics or particular movements of sharks following prey (Heithaus et al., 2007; Ketchum et al., 2009). Therefore, tracked sharks in this study could have been either exploring the diverse seamounts found around Malpelo or Cocos Island, in the middle of a round trip following preys, or migrating to other sectors in the Tropical Eastern Pacific (Bessudo et al., 2011).
Tags in this study were inserted by expert free divers, however tag loss was considerably high due to the difficulty of the tagging operation. Other studies using PAT tags on Galapagos (C. galapagensis) and tiger sharks (G. cuvier), such as in Meyer et al. (2010), also experienced a short retention of PAT tags compared with other tagging methods (e.g., SPOT and internal implanted ultrasonic tags). Even thought Meyer et al. (2010) captured the animals and implanted the tags by the side of the boat, PAT retention times for Meyer et al. (2010) study were all less than one-third of the programmed 100-day deployment time. Apparently, tag shedding is an inherent problem of using PAT tags.
Our findings showed that depth ranges at which hammerhead sharks swam depended mostly on the season, possibly due to differences in primary productivity. For example, at Malpelo Island, during the cold water season primary productivity is higher in surface waters, principally due to the upwelling of nutrients and chlorophyll-a content (Rodríguez-Rubio et al., 2007). Consequently, during this period there is a significant increase in food availability, possibly influencing indirectly (higher primary productivity correlates with higher shark prey abundance) hammerhead sharks swimming preferences, making them spend more time at surface waters near the island. Conversely, due to the absence of upwelling during the warm water season, nutrient contents are found at around 60 m deep (Rodríguez- Rubio et al., 2007). The overall lower food availably during this period could be triggering sharks to move away from the island, swimming into deeper depths in search of other food resources. Another possible factor influencing the observed pattern is the reproduction cycle: during the cold water season large aggregations of hammerhead sharks have been observed schooling at surface waters around Malpelo Island, and then leaving the island in large groups (Bessudo et al., 2011), possibly for reproductive purposes (Hazin et al., 2001).
When sharks swam offshore, they made several short deep dives in a yoyo manner, to at least 1000 m where water temperatures reached 4 °C. These are the deepest dives reported in the literature so far for this species. Klimley (1993) and Klimley et al. (2002) also registered yo-yo dives for hammerhead sharks, using acoustic telemetry in the Bajo de Espíritu Santo, Mexico. This same behavior has also been observed for other species of sharks like the thresher shark (Nakano et al., 2003) and the white shark (Bonfil et al., 2005). Possible explanations for these deep dives into the bathypelagic zone, where there are low levels of dissolved oxygen, are to access special preys, such as deep water squids, or to avoid prey competition with other pelagic predators (Jorgensen et al., 2009).
The large amount of time spent at deep depths (>100 m) during night time found by this study is consistent with Bessudo et al. (2011) results using acoustic telemetry. It is possible that hammerhead sharks remain close to Malpelo Island during the day using it as a resting station, and during the night swim into deeper waters in search for preys.
This sharks' high mobility in and out of marine protected areas makes them extremely vulnerable to domestic fisheries as well as multinational fisheries on the high seas. Thus, future long-term monitoring of this species is required to better understand their spatial and temporal movement patterns, especially for populations inhabiting the Pacific Ocean, were the world's largest industrial tuna fleet operates (Joseph, 2003). This information is essential to provide sustainable management tools to reduce fisheries shark capture rates. Furthermore, this study confirmed the expanded vertical niche of S. lewini, which can tolerate extremely low levels of dissolved oxygen (Jorgensen et al., 2009). However, our results indicated that hammerhead sharks spent most of their time near surface waters (<100 m), a behavior that makes them very vulnerable to the fishing industry. Local and regional agreements to limit shark capture and to regulate and monitor all shark parts trade, as well as the creation of new marine protected areas, are key actions not only to enhance the protection and conservation of this species, but also because they are an important source of income for ecotourism (Orams, 1996; Davis and Banks, 1998; Landman, 2000; Norman, 2000; Pedersen, 2002; Chapman et al., 2005; Dobson, 2006; Torres et al., 2007).
ACKNOWLEDGMENTS
We thank Julia Miranda, director of the Special Administrative Unit of the National Natural Parks System of Colombia, to Guillermo Barrera and Mauricio Soto, commanders of the Colombian Navy for their permanent support. Many thanks to Jose Luis Gómez, director of the Fondo para la Acción Ambiental y la Niñez, the Whiltley Fund for Nature, the SEASCAPE project funded by GCF/UNESCO, Conservation International, the Walton Family Foundation and the Malpelo Fauna and Flora Endowment Fund for financing this project. We also thank Fabio Arjona and María Claudia Diazgranados for their great support, Fred Buyle and William Winram for tagging the sharks, Yves Lefèvre for the documentary of this project, the University of Stanford and the TOPP program, Philippe Gaspar and Francois Royer from CLS, France, and to Efraín Rodríguez-Rubio.
LITERATURE CITED
1 Baum, J. K., R. A. Myers, D. G. Kehler, B. Worm, S. J. Harley and P. A. Doherty. 2003. Collapse and conservation of shark populations in the Northwest Atlantic. Science, 299 (5605): 389-392. [ Links ]
2 Bessudo, S., G. Soler, A. P. Klimley, J. T. Ketchum, R. Arauz and A. Hearn. 2011. Residency of the scalloped hammerhead shark (Sphyrna lewini) at Malpelo Island and evidence of migration to other islands in the Eastern Tropical Pacific. Environ. Biol. Fishes, 91 (2): 165-176. [ Links ]
3 Bonfil, R., M. Meyer, M. C. Scholl, R. Johnson, S. O'Brien, H. Oosthuizen, S. Swanson, D. Kotze and M. Paterson. 2005. Transoceanic migration, spatial dynamics, and population linkages of white sharks. Science, 310 (5745): 100-103. [ Links ]
4 Camhi, M. 1998. Sharks on the line: A state by state analysis of sharks and their fisheries. Living Oceans Program, National Audubon Society, Islip, New York. 158 p. [ Links ]
5 Camhi, M. D., E. Lauck, E. K. Pikitch and E. A Babcock. 2008. A global overview of commercial fisheries for Open Ocean sharks. 166-192. In: Camhi, M. D., E. K. Pikitch and E. A. Babcock (Eds.). Sharks of the open ocean: Biology, fisheries and conservation. Blackwell Publishing, Oxford. 536 p. [ Links ]
6 Castillo, P., R. Batista, D. Vanko, E. Malavassi, J. Barqueo and E. Fernández. 1988. Anomalously young volcanoes on hot-spot traces: I. Geology and petrology of Cocos Island. Geol. Soc. Amer. Bull., 100: 1400-1414. [ Links ]
7 Chapman, D. D., E. K. Pikitch, E. Babcock and M. S. Shivji. 2005. Marine reserve design and evaluation using automated acoustic telemetry: A case-study involving coral reef-associated sharks in the Mesoamerican Caribbean. Mar. Technol. Soc. J., 39 (1): 42-55 p. [ Links ]
8 Davis, D. and S. Banks. 1998. Whale-sharks in Ningaloo Marine Park: managing tourism in an Australian marine protected area. Tourism Manag, 18 (5): 259-271. [ Links ]
9 Devis-Morales, A., W. Schneider, R. A. Montoya-Sánchez and E. Rodríguez-Rubio. 2008. Monsoon-like winds reverse oceanic circulation in the Panama Bight. Geophys. Res. Lett., 35: L20607. DOI: 10.1029/2008GL035172. [ Links ]
10 Dobson, J. 2006. Sharks, wildlife, tourism, and state regulation. Tourism Mar. Environ., 3 (1): 15-23. [ Links ]
11 Ekstrom, P.A. 2004. An advance in geolocation by light. Mem. Natl. Inst. Polar Res., Spec. Issue, 58: 210-226. [ Links ]
12 FAO. 2000. Fisheries management 1. Conservation and management of sharks. FAO Technical Guidelines for Responsible Fisheries. No. 4. Suppl. 1. Rome. 37 p. [ Links ]
13 Ferrari, A. and A. Ferrari. 2001. Guide des Requins. Plus de 120 especes du monde entier. Delachux et Niestlé, Paris. 256 p. [ Links ]
14 Garla, R. C., D. D. Chapman, B. M. Wetherbee and M. Shivji. 2006. Movment patterns of young Caribbean reef sharks, Carcharhinus perezi, at Fernando de Noronha Archipielago, Brazil: the potential of marine protected areas for conservation of a nursery ground. Mar. Biol., 149: 189-199. [ Links ]
15 Glen, A. S., C. R. Dickman, M. E. Soule and B. G. Mackey. 2007. Evaluating the role of the dingo as a trophic regulator in Australian ecosystems. Aus. Ecol., 32: 492-501. [ Links ]
16 Griffin, E., K. L. Miller, B. Freitas and M. Hirshfield. 2008. Predators as prey: Why healthy oceans need sharks. http://na.oceana.org/en/news-media/publications/reports?page=3. 03/07/2011. [ Links ]
17 Hazin, F., A. Fisher and M. Broadhurst. 2001. Aspects of reproductive biology of the scalloped hammerhead shark, Sphyrna lewini, off Northeastern Brazil. Environ. Biol. Fishes, 61 (2): 151-159. [ Links ]
18 Heithaus, M. R., A. J. Wirsing, L. M. Dill and L. I. Heithaus. 2007. Long-term movements of tiger sharks satellite-tagged in Shark Bay, Western Australia. Mar. Biol., 151: 1455-1461. [ Links ]
19 IUCN. 2007. The conservation status of pelagic sharks and rays. Report of the IUCN Shark specialist group. Pelagic sharks red list workshop. Tubney house, University of Oxford, Oxford. 92 p. [ Links ]
20 IUCN. 2010. IUCN red list of threatened species. Version 2010.4. www.iucnredlist.org. 01/02/2011. [ Links ]
21 Jennings, S., M. J. Kaiser and J. D. Reynolds. 2005. Marine fisheries ecology. Blackwell Science Ltd., Oxford. 432 p. [ Links ]
22 Jorgensen, S. J. J., A. P. Klimley and A. F. Muhlia-Melo. 2009. Scalloped hammerhead shark Sphyrna lewini, utilizes deep-water, hypoxic zone in the Gulf of California. J. Fish Biol., 74: 1682-1687. [ Links ]
23 Joseph, J. 2003. Managing fishing capacity of the world tuna fleet. FAO, Fisheries Circular No. 982, Rome. 67 p. [ Links ]
24 Ketchum, J., A. Hearn, G. Shillinger, E. Espinoza, C. Peñaherrera and P. Klimley. 2009. Shark movements and the design of protected pelagic environments within and beyond the Galapagos Marine Reserve. 127-130. In: Wolff, M. and M. Gardener (Eds.). Proceedings of the Galapagos Science Symposium, Charles Darwin Foundation, Puerto Ayora, Galápagos. 230 p. [ Links ]
25 Klimley, A. P. 1993. Highly directional swimming by scalloped hammerhead sharks, Sphyrna lewini, and subsurface irradiance, temperature, bathymetry, and geomagnetic firld. Mar. Biol., 117: 1-22 p. [ Links ]
26 Klimley, A. P., S. C. Beaversa, T. H. Curtisa and S. Jorgensen. 2002. Movements and swimming behavior of three species of sharks in La Jolla Canyon, California. Environ. Biol. Fishes, 63: 117-135. [ Links ]
27 Landman, G. 2000. Evaluación de los sitios de turismo marino en la Reserva Marina de Galápagos. Estación Científica Charles Darwin, Galápagos. 7 p. [ Links ]
28 Lizano, O. G. 2008. Dinámica de aguas alrededor de la Isla del Coco, Costa Rica. Rev. Biol. Trop., 56 (2): 31-48. [ Links ]
29 López-Victoria, M. and D. Rozo. 2006. Model-based geomorphology of Malpelo Island and spatial distribution of breeding seabirds. Bol. Invest. Mar. Cost., 35: 111-131. [ Links ]
30 Meyer, C. G., Y. P. Papastamatiou and K. N. Holland. 2010. A multiple instrument approach to quantifying the movement patterns and habitat use of tiger (Galeocerdo cuvier) and Galapagos sharks (Carcharhinus galapagensis) at French Frigate Shoals, Hawaii. Mar. Biol., 157 (8): 1857-1868, DOI: 10.1007/s00227-010-1457-x. [ Links ]
31 Nakano, H., H. Matsunaga, H. Okamoto and M. Okazaki. 2003. Acoustic tracking of bigeye thresher shark Alopias superciiliosus in the Eastern Pacific Ocean. Mar. Ecol. Prog. Ser., 265: 255-261. [ Links ]
32 Newmark, F. U. and M. Santos-Acevedo. 2002. Valoración de los recursos marinos y costeros en Colombia. 83-99. En: Ospina-Salazar, G. H. y A. Acero (Eds.). Informe del estado de los ambientes marinos y costeros en Colombia: año 2001. Serie de Publicaciones Periódicas No. 8., Instituto de Investigaciones Marinas y Costeras, Santa Marta. 178 p. [ Links ]
33 Norman, B. 2000. Rhincodon typus. 2006 IUCN Red List of Threatened Species. http://www.iucnredlist.org. 06/01/2004. [ Links ]
34 Orams, M. 1996. Using interpretation to manage nature-based tourism. J. Sustain. Tourism, 4 (2): 81-94. [ Links ]
35 Pade, N. G., N. Queiroz, N. E. Humphries, M. J. Witt, C. S. Jones, L. R. Noble and D. W. Sims. 2009. First results from satellite-linked archival tagging of porbeagle shark, Lamna nasus: area fidelity, wider-scale movements and plasticity in diel deph changes. J. Exp. Mar. Biol. Ecol., 370: 64-74. [ Links ]
36 Pedersen, B. 2002. Population trends and their impact on Western Australian beach use. Coast to Coast, Sydney. 356 p. [ Links ]
37 Rodríguez-Rubio, E., W. Schneider and R. Abarca del Río. 2003. On the seasonal circulation within the Panama Bight derived by satellite observations of wind altimetry and sea surface temperature. Geophys. Res. Lett., 30 (7): 1410. [ Links ]
38 Rodríguez-Rubio, E., J. Ortiz-Gálviz and J. Rueda-Bayona. 2007. Aspectos Oceanográficos. 29-44. In: Dirección General Marítima-Centro Control Contaminación del Pacífico y Unidad Administrativa Especial del Sistema de Parques Nacionales Naturales-Dirección Territorial Suroccidente (Eds.). Santuario de Fauna y Flora Malpelo: descubrimiento en marcha. Editorial DIMAR, Serie Publicaciones Especiales Vol. 5, Bogotá. 142 p. [ Links ]
39 Rose, D. A. 1996. An overview of world trade in sharks and other cartilaginous fishes. Traffic International, Cambridge. 106 p. [ Links ]
40 Royer, F. and M. Lutcavage. 2009. Positioning pelagic fish from sunrise and sunset times: Complex observation errors call for constrained, robust modeling. 323-341. In: Nielsen, J. L., N. Fragoso, M. Lutcavage, H. Arrizabalaga, A. Hobday and J. Sibert (Eds.). Second International Symposium on Tagging and Tracking Marine Fish with Electronic Devices. Rev. Fish Biol. Fisher., 9: 323-341. [ Links ]
41 Schneider, D. C. 1994. Quantitative ecology: spatial and temporal scaling. Academic Press, San Diego. 402 p. [ Links ]
42 Sciarrotta, T. C. and D. R. Nelson. 1977. Diel behavior of the blue shark, Prionace glauca, near Santa Catalina Island, California. U.S. Fish. Bull., 75: 519-528. [ Links ]
43 Sedberry, G. R. and J. K. Loefer. 2001. Satellite telemetry tracking of swordfish, Xiphias gladius, of the eastern United States. Mar. Biol., 139: 355-360. [ Links ]
44 Sergio, F., I. Newton, L. Marchesi and P. Pedrini. 2006. Ecological justified charisma: preservation of top predators delivers biodiversity conservation. J. Appl. Ecol., 43: 1049-1055. [ Links ]
45 Sibajo-Cordero, J. A. 2008. Tendencias espacio-temporales de los avistamientos de fauna marina en los buceos turísticos (Isla del Coco, Costa Rica). Rev. Biol. Trop., 56 (2): 113-132. [ Links ]
46 Simpfendorfer, C. A. and M. R. Heupel. 2004. Assessing habitat use and movement. 553-572. In: Carrier J. C., J. A. Musick and M. R. Heithaus (Eds.). Biology of sharks and their relatives. CRC Press, Boca Ratón. 608 p. [ Links ]
47 Stevens, J. D. 2000. The population status of highly migratory oceanic sharks. 35-43. In: Hinman, K. (Ed.). Getting ahead of the curve: Conserving the Pacific ocean's tunas, swordfish, billfishes and sharks. National Coalition for Marine Conservation, Leesburg, USA. 200 p. [ Links ]
48 Temple, S. A. 1987. Do predators always capture substandard individuals disproportionately from prey populations? Ecology, 68 (3): 669-674. [ Links ]
49 Torres, N., R. Andrade and N. Rodríguez. 2007. Community-based management through ecotourism in Bahía de los Ángeles, Mexico. Fish. Res., 84 (1): 114-118. [ Links ]
50 Watts, S. 2001. The end of the line? WildAid Report. Wendy P. McCaw Foundation. http://www.wildaid.org. 06/05/2010. [ Links ]
51 Welch, D. W. and J. P. Eveson, 1999. An assessment of light-based geoposition estimates from archival tags. Can. J. Fish. Aquat. Sci., 56 (7): 1317-1327. [ Links ]
52 Wilson, S. G., M. E. Lutcavage, R. W. Brill, M. P. Genovese, A. B. Cooper and A. W. Everly. 2005. Movements of bluefin tuna (Thunnus thynnus) in the northwestern Atlantic Ocean recorded by popup satellite archival tags. Mar. Biol., 146: 409-423. [ Links ]
FECHA DE RECEPCIÓN: 26/08/2010 FECHA DE ACEPTACIÓN: 12/10/2011
