










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.40 supl.1 Santa Marta Dec. 2011
COMPOSICIÓN Y ESTRUCTURA TRÓFICA DE LOS MACROINVERTEBRADOS TERRESTRES DE LA ISLA MALPELO, PACÍFICO COLOMBIANO*
COMPOSITION AND TROPHIC STRUCTURE OF TERRESTRIAL MACROINVERTEBRATES OF MALPELO ISLAND, COLOMBIAN PACIFIC
David Calero1, Mateo López-Victoria2,3 y Patricia Chacón de Ulloa1
1 Universidad del Valle, Facultad de Ciencias Naturales y Exactas, Departamento de Biología, Grupo de Investigación Biología, Ecología y Manejo de Hormigas, A.A. 25360 Cali, Colombia. davidrc114@hotmail.com (D.C.), pachacon@univalle.edu.co (P.C.U.).
2 Universidad Justus-Liebig, Departamento de Ecología Animal, Giessen, Alemania. gf1617@uni-giessen.de. (Dirección actual).
3 Instituto de Investigaciones Marinas y Costeras-INVEMAR , Cerro Punta Betín, Santa Marta, Colombia.
RESUMEN
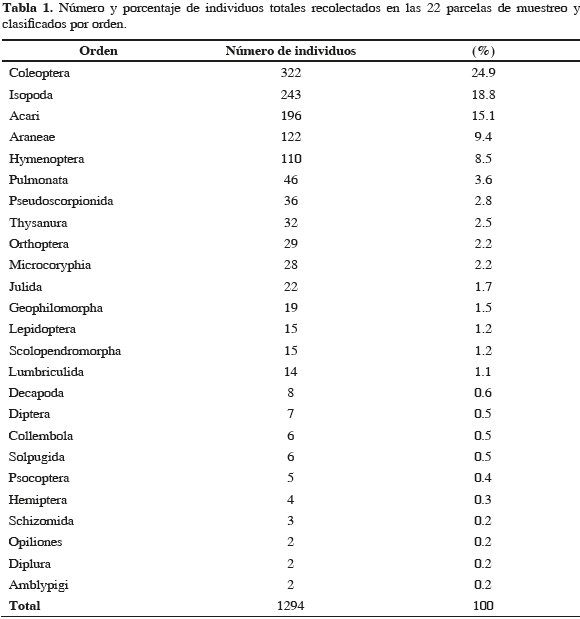
Las islas oceánicas poseen ecosistemas con dinámicas especiales debido a su aislamiento y condiciones climáticas influenciadas por el mar. Es el caso de la isla Malpelo, localizada en el Pacífico colombiano, en donde estudios previos mostraron una composición de invertebrados fuertemente dominada por depredadores. Con el propósito de contribuir al conocimiento de estos organismos en la isla, se realizaron muestreos en 22 parcelas, abarcando tres tipos de microhábitat, clasificados de acuerdo al tipo de suelo, sombra y vegetación. Las colectas se realizaron entre mayo y junio de 2006, mediante captura directa. Se obtuvieron 1294 especímenes, pertenecientes a siete clases y 25 órdenes. La diversidad estuvo determinada por la complejidad en la estructura del microhábitat, siendo más diverso el que presentó un mayor grado de heterogeneidad. El orden Aranae presentó el mayor número de especies, Coleoptera la mayor abundancia y Staphylinidae la mayor abundancia relativa. Los depredadores presentaron la mayor riqueza (45.3 %) y abundancia (40.6 %) de todos los gremios tróficos. Estos resultados refuerzan la idea de que el sistema terrestre de Malpelo es dependiente del mar.
PALABRAS CLAVE: Isla oceánica, Invertebrados terrestres, Gremios tróficos, Riqueza, Abundancia.
ABSTRACT
Oceanic islands have special ecosystem dynamics due to their isolation and weather conditions influenced by the sea. This is also the case for Malpelo Island, located at the Colombian Pacific, where previous studies showed an invertebrate composition strongly dominated by predatory species. With the aim of contributing to the knowledge of these organisms on the island, 22 sample plots were surveyed, covering three types of microhabitat, classified according to the type of soil, shadow, and vegetation. Sampling took place between May and June 2006, by direct catches. A total of 1294 specimens from seven classes and 25 orders were obtained. Diversity was determined by microhabitat structure complexity, being more diverse the one that presented the greatest degree of heterogeneity. The order Araneae presented the highest number of species, Coleoptera the highest abundance and Staphylinidae the highest relative abundance. Predators showed the highest richness (45.3 %) and abundance (40.6 %) among all trophic guilds. These findings lent support to the idea that the terrestrial system of Malpelo depends on the sea.
KEY WORDS: Oceanography, Water mass, Eastern Tropical Pacific, Malpelo Island, Colombia.
INTRODUCCIÓN
Las islas oceánicas tienen su origen en los fondos marinos y son el resultado de la actividad volcánica y la dinámica de la tectónica de placas. La gran mayoría se encuentra situada lejos de los continentes y separada de éstos por grandes profundidades (MacArthur y Wilson, 1967; Whittaker y Fernández-Palacios, 2007). Este tipo de islas exhiben ecosistemas con dinámicas especiales, por su grado de aislamiento y condiciones climáticas influenciadas por los cuerpos de agua circundantes (Carlquist, 1965, 1974; Whittaker y Fernández-Palacios, 2007). Se caracterizan por presentar una fauna y flora particular, poco diversa y con altos niveles de endemismo (Carlquist, 1974; Burggren y McMahon, 1988; Whittaker y Fernández-Palacios, 2007).
Los invertebrados terrestres son un grupo interesante para estudiar en islas oceánicas, ya que son organismos que presentan una alta diversidad y se encuentran en todos los ecosistemas del mundo (Borror et al., 1992; Fernández-Palacios y Morici, 2004). Incluso en la isla más remota y poco diversa habitan al menos algunos invertebrados, que juegan un papel clave en la formación del suelo, en la transformación de la fase orgánica (Fitzpatrick, 1992), en la dinámica de los perfiles del sustrato (Chamorro y Romero, 1990) y en las redes tróficas y la conversión de energía (Ulrich, 2004).
El conocimiento de la diversidad y ecología faunística terrestre de la isla Malpelo ha sido escaso, ya que la mayoría de investigaciones se han enfocado en inventarios y descripciones diagnósticas de vertebrados terrestres y acuáticos. En cuanto a los invertebrados terrestres se destaca el trabajo de Wolda (1975), quien registró 40 especies de 24 familias y 21 órdenes, además de proponer un modelo simplificado de red trófica para mostrar la estructura del ecosistema. Ese investigador encontró que la diversidad de depredadores era mayor que la de los herbívoros y descomponedores, de lo que dedujo que entre las especies depredadoras el mayor porcentaje debía corresponder a especies generalistas, en lugar de a especialistas.
Posteriormente al trabajo pionero de Wolda (1975), Álvarez-Rebolledo et al. (1999) publicaron un nuevo inventario de invertebrados terrestres, a partir de colecciones realizadas por el Instituto Alexander von Humboldt en 1998. En ese compendio se registran cinco clases, 13 órdenes y 25 familias, con 42 especies.
El presente trabajo es la tercera iniciativa dedicada exclusivamente a los macroinvertebrados terrestres presentes en Malpelo, y tuvo como objetivos realizar un inventario actualizado, estudiar su diversidad en función del tipo de sustrato y elaborar un modelo preliminar de la red trófica. Al margen del aporte científico, este trabajo cumple la función de ser punto de referencia para futuras comparaciones y para adecuados programas de manejo y prevención en la isla.
ÁREA DE ESTUDIO
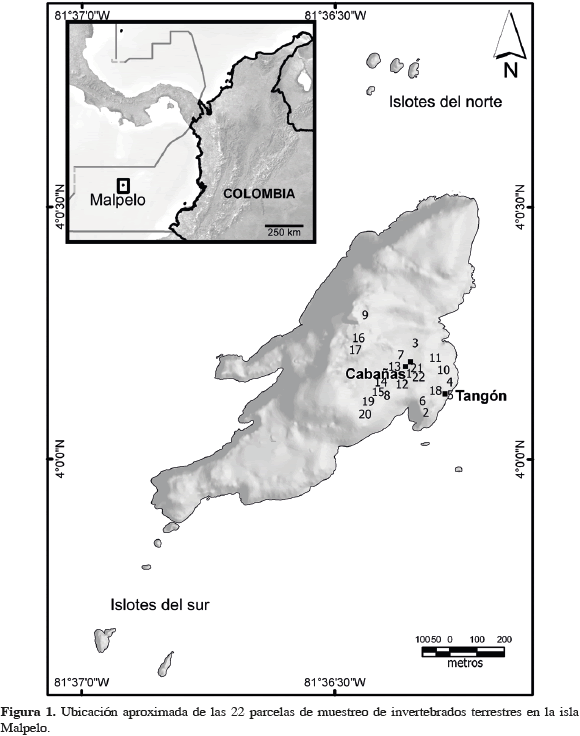
Malpelo está ubicada en el Pacífico colombiano, 500 km al oeste de Buenaventura (Valle del Cauca) (04°0'00" N, 81°3'30" W). Es la cúspide de una cordillera sumergida que se levanta paralela a la costa continental, desde una profundidad de 4000 m. La isla está rodeada por once pequeños islotes y presenta alturas máximas cercanas a 300 m. Es de origen volcánico, exhibe acantilados muy escarpados en casi todos sus márgenes y tiene 1.2 km2 de superficie emergida (Chase, 1968; Stead, 1975; Prahl, 1990; López-Victoria y Rozo, 2006) (Figura 1). El clima se encuentra bajo la influencia de la franja ecuatorial de la Zona de Convergencia Intertropical (ZCI), es predominantemente cálido, húmedo y con precipitaciones durante la mayor parte del año (Prahl, 1990; Brando et al., 1992; López-Victoria y Estela, 2007).
La vegetación en Malpelo es escasa, y está representada por un helecho rastrero (Pteridophyta: Ptyrogramma dealbata), cuatro géneros de líquenes (Calopaca, Candelaria, Lecidea y Pyxine), microalgas y musgos (Wolda, 1975; Prahl, 1990; Brando et al., 1992). Los principales exponentes de la macrofauna terrestre son tres lagartos endémicos, Diploglossus millepunctatus, Phyllodactylus transversalis y Anolis agassizi (Graham, 1975; Prahl, 1990; Brando et al., 1992), más de 60 especies de aves (López-Victoria y Estela, 2007) y un interesante ensamblaje de invertebrados, representado por anélidos, moluscos, crustáceos, insectos y arácnidos entre otros (Wolda, 1975; Álvarez-Rebolledo et al., 1999). A pesar de que existen registros de visitas a la isla desde 1542 (Gorman y Chorba, 1975), solamente a partir de 1986 ha estado permanentemente ocupada por infantes de marina. Además, es visitada esporádicamente por turistas e investigadores, la mayoría de ellos interesados principalmente en actividades de buceo.
MATERIALES Y MÉTODOS
Fase de campo
Los muestreos fueron realizados entre mayo y junio de 2006, en 22 parcelas ubicadas en el costado oriental de Malpelo (Figura 1). Cada parcela, de 50 x 50 cm, fue demarcada en el suelo con una cuerda extendida. Al interior de cada parcela, tres personas recolectaron simultáneamente todos los macroinvertebrados, durante un lapso de tiempo de 70 minutos en promedio, empleando pinzas y aspiradora bucal; sólo se capturaron cangrejos terrestres con ancho de caparazón <15 mm. El muestreo en cada parcela se dio por terminado cuando no hubo más macroinvertebrados visibles, hasta una profundidad de 5 a 7 cm por debajo de la superficie; las piedras en la superficie o parcialmente enterradas fueron removidas y revisadas. Todos los organismos fueron preservados en alcohol etílico al 70 %. La ubicación de las parcelas obedeció al tipo de sustrato (microhábitat), en un ámbito de altura de 20 a 270 msnm. A partir de los criterios "tipo de suelo" (tierra o roca), "sombra" (con o sin) y "presencia-ausencia de vegetación", se establecieron los siguientes tres microhábitat: Sustrato S1, compuesto por piedras muy pequeñas (<20 mm) a medianas (10-20 cm), sobre suelo descubierto (roca), sin sombra (incluidos los nidos de Sula granti); Sustrato S2, compuesto por piedras pequeñas (2-10 cm) a medianas (10-20 cm), sobre suelo de tierra, con sombra (cuevas); Sustrato S3, compuesto por piedras medianas (10-20 cm), sobre suelo de tierra, entre vegetación (helecho rastrero).
Fase de laboratorio
La determinación taxonómica de las muestras se realizó hasta el mejor nivel de resolución posible, utilizando un microscopio estereoscópico (Nikon K-891), con base en las guías de identificación de Borror et al. (1992), De la Fuente (1994), Flórez (1996), Fernández (2003) y Ubick et al. (2005). Ya que la mayoría de organismos se determinó hasta el nivel de familia, para efectos prácticos, en adelante el término "especie" se utilizará indistintamente para referirse tanto a los organismos que fueron completamente identificados, como a aquellos que quedaron hasta el nivel de morfoespecie.
Para estimar la biomasa de los distintos gremios tróficos, evitando en el proceso el deterioro de las muestras por desecación, se utilizó el peso húmedo, adaptado a partir del método empleado por Rodrigues et al. (2009). Las muestras se dejaron secar del exceso de alcohol durante cinco minutos, a temperatura ambiente (aprox. 24 ºC), sobre un papel absorbente, y luego se pesaron en una balanza analítica (Ohaus ± 0.0001 g). Todos los especímenes se encuentran depositados en el Museo de Entomología de la Universidad del Valle, Cali, Colombia (MUSENUV).
Análisis de los datos
Para evaluar el esfuerzo de muestreo se utilizó Chao1, basado en la abundancia, mediante el programa EstimateS 8.1 (Colwell, 2005). Los gremios tróficos fueron asignados con base en la biología de los distintos grupos taxonómicos y en el reconocimiento de los distintos tipos de estructuras bucales de los organismos. Se estimó la biomasa total que aporta cada gremio al sistema, a partir de la superficie obtenida del modelo tridimensional de Malpelo (ver López-Victoria y Rozo, 2006). Para caracterizar una parcela "típica" de la isla, se analizaron los datos de riqueza, abundancia y biomasa promedio de las 22 parcelas de muestreo. Para comparar la diversidad de los microhábitat se utilizó el índice de Shannon y una prueba t de Student. El modelo preliminar de red trófica fue construido a partir de la biomasa estimada para cada gremio trófico.
RESULTADOS
Riqueza y abundancia
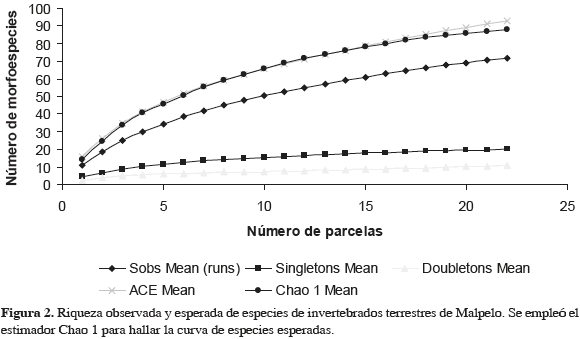
En las 22 parcelas se capturó un total de 1294 invertebrados terrestres, correspondientes a siete clases, 25 órdenes y, al menos, a 23 familias y 72 morfoespecies (Anexo 1). Según el estimativo Chao 1 se capturó el 82 % de las especies esperadas, faltando por capturar 16 especies más (Figura 2). Las clases encontradas y su representación porcentual en cuanto al número de órdenes fueron: Hexapoda (48 %), Arachnida (28 %), Malacostraca (8 %), Mollusca (4 %), Chilopoda (4 %), Diplopoda (4 %) y Oligochaeta (4 %). Los hexápodos y los arácnidos fueron los grupos más diversos, con doce y siete órdenes, respectivamente. El orden con mayor riqueza fue Araneae con 20 especies, seguido por Acari (11), Coleoptera (8) y Diptera (5). En los otros 22 órdenes se encontraron las restantes 28 especies (38.9 %), pero ninguno de éstos aportó más del 7 % a la riqueza total.
Los órdenes con mayor abundancia fueron Coleoptera (25 %), Isopoda (18.9 %), Acari (15.2 %) Araneae (9.2 %) e Hymenoptera (8.6 %), que totalizan el 76.7 % de todas las muestras (Tabla 1). Los coleópteros de la familia Staphylinidae (Aleocharinae) presentaron la mayor abundancia (23 %), seguidos por las cochinillas terrestres del suborden Oniscidoidea (18.9 %) y las arañas Salticidae (2.4 %), Linyphiidae (2 %) y Pholcidae (1.78 %). La abundancia de himenópteros se debió a la gran cantidad de hormigas del género Pheidole (7 % del total de especímenes capturados).
Estructura y redes tróficas
Los depredadores presentaron la mayor riqueza, constituyendo el 45.3 % de todas las especies y la mayor diversidad por parcela muestreada (Anexo 1, Figura 3). Después de los depredadores, el gremio más diverso fue el de los saprófagos, pero con la mitad de especies menos. En cuanto a la abundancia, se repitió el mismo patrón anterior, con los parásitos como el tercer gremio con el mayor número de individuos por parcela. No obstante su relativa abundancia, debido a su pequeño tamaño, la biomasa aportada por los parásitos fue la menor de todas. En este sentido los saprófagos fueron el gremio con el mayor aporte, seguidos de lejos por los depredadores (Figuras 3 y 4). En síntesis, una parcela promedio estuvo constituida por un total de 10.5 especies, 58.5 individuos y un aporte de biomasa de 642.5 mg.
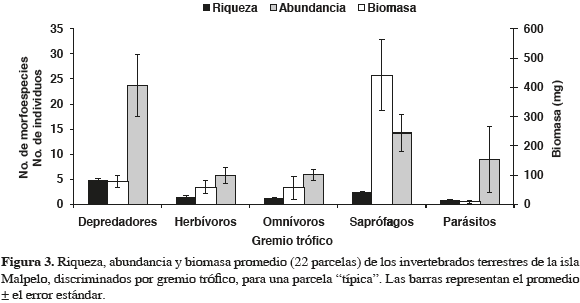
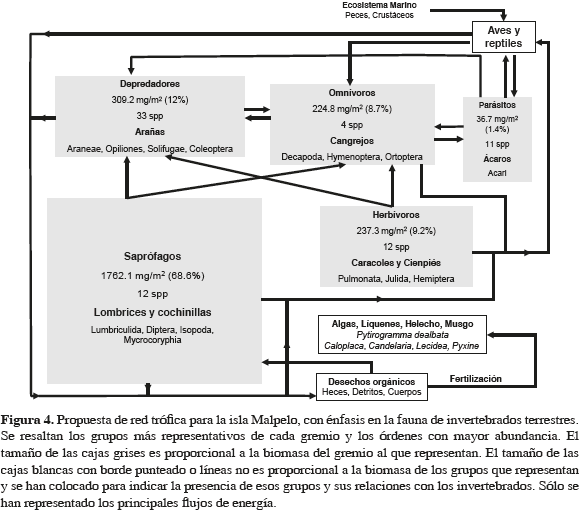
En el gremio de los saprófagos, con el 68.6 % de biomasa promedio por parcela, se destacaron principalmente las lombrices y las cochinillas (Figura 4). Los depredadores, a pesar de ser el gremio con mayor riqueza y abundancia, aportaron solamente el 12 % de la biomasa total. Los herbívoros, caracoles, milípedos y microlepidópteros, ocuparon el tercer lugar con 9.2 %, mientras que los omnívoros, principalmente cangrejos (juveniles con ancho de caparazón <15 mm) y grillos, ocuparon el cuarto lugar (8.7 %). Por último se encontraron los parásitos, que aportaron tan sólo el 1.4 %. Teniendo en cuenta el promedio de la biomasa, 2570 mg/m2, y la superficie de la isla habitada por macroinvertebrados terrestres, 0.813 km2, la biomasa total de estos organismos fue de 2.1 tons aproximadamente (entre 1.7 y 2.5 tons, al incorporar el error estándar al cálculo de los valores mínimo y máximo, respectivamente).
Estructura del microhábitat
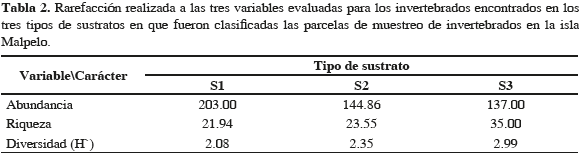
El sustrato S3 presentó la menor abundancia, pero la mayor riqueza de especies y el valor más alto en el índice de diversidad (Tabla 2). Los sustratos S1 y S2 mostraron valores similares de riqueza y diversidad, aunque el S1 tuvo la mayor abundancia. Las diferencias entre microhábitat en cuanto a la diversidad fueron significativas entre los sustratos S1-S3 y S2-S3 (S1 vs S2, t= 11.525, p >0.05; S1 vs. S3, t= -25.551, p <0.05; S2 vs S3, t= 30.808, p <0.005).
DISCUSIÓN
Riqueza y abundancia
Wolda (1975) basó sus registros en capturas directas en la parte sur de la isla y en revisiones de contenidos estomacales y de excrementos de lagartos. Por su parte Álvarez et al. (1999) no mencionan el método de muestreo utilizado (pudo tratarse de capturas esporádicas). El estimador de riqueza indica que durante el presente estudio se habrían podido encontrar hasta 16 especies más de invertebrados terrestres en Malpelo. Ese resultado, sumado a la aparición de especies representadas por un solo individuo, sugieren que el inventario fue incompleto, ya que en la naturaleza no existen individuos "solos", sino poblaciones (Villareal et al., 2004). No obstante, ya que el método de muestreo es el utilizado para capturar especies raras o muy escasas (Villareal et al., 2004), el nivel de inventario alcanzado es aceptable. Otros métodos complementarios de captura (p.ej., sacos Winkler, trampas de caída y cebos atrayentes) permitirían complementar el inventario y capturar otros invertebrados terrestres que son esquivos a la captura directa.
Al igual que el patrón general observado en la naturaleza alrededor del mundo (Ruppert y Barnes, 1996), el mayor número de órdenes en Malpelo lo presentaron las clases Insecta y Arachnida. Este resultado, guardadas las proporciones en cuanto al área emergida y la cobertura vegetal, fue similar en magnitud a lo encontrado en otros estudios en islas oceánicas del Pacífico Oriental: en las islas Galápagos el número de especies de insectos registrados fue superior a 2000, mientras que unas 350 especies adicionales correspondieron a otros invertebrados terrestres (Peck, 2005); en la isla de Cocos (Costa Rica) más del 50 % de las especies de invertebrados terrestres identificadas correspondieron a insectos (Hogue y Miller, 1981); en el archipiélago de Revillagigedo (México) se registraron 119 especies de invertebrados en la isla Socorro, de las cuales 74.9 % correspondieron a insectos y 22.7 % a arácnidos (Turnbull, 1973). Estos altos porcentajes se deben principalmente a que los insectos son el grupo animal con mayor diversidad y capacidad de dispersión, ya que poseen características morfo-fisiológicas que les han permitido colonizar prácticamente todos los ambientes de la tierra (Ruppert y Barnes, 1996).
En el caso particular de los arácnidos, su abundancia en las comunidades es suficientemente grande como para efectuar análisis numéricos que permitan investigaciones sobre estructura de comunidades, estratificación y sucesión (Turnbull, 1973). Tanto en ambientes naturales como intervenidos las arañas presentan los mayores índices de captura (Buddle et al., 2006), en parte debido a su facilidad para dispersarse y colonizar nuevos hábitats (Halaj et al., 1998). En este sentido, Malpelo tampoco es la excepción, ya que, en cuanto a riqueza, este grupo presentó el mayor número de especies, pertenecientes principalmente a la familia Salticidae, que a su vez presentó el mayor número de individuos del grupo. Resultados similares fueron obtenidos por Rico et al. (2005) al estudiar la diversidad de arañas en la isla Gorgona (Pacífico colombiano), donde esta familia también fue la más diversa con 40 morfoespecies, representando el 16.2 % de todas las morfoespecies capturadas. Este patrón no solamente se presenta en islas. Los resultados obtenidos en bosques continentales en Perú (Silva, 1996; Silva y Coddington, 1996), en Panamá (Nentwig, 1993) y en Colombia (Flórez y Sánchez, 1995) apuntan en la misma dirección, ya que la familia Salticidae ocupa los primeros lugares en riqueza y abundancia. Las especies de esta familia de arañas se caracterizan por cazar al acecho, y hay más de 500 géneros y 5000 especies de esta familia en el mundo (13 % de Araneae; Peng et al., 2002).
Dentro del grupo de los quelicerados, a nivel mundial el primer lugar en riqueza y abundancia lo ocupan los ácaros, que son la biota dominante tanto en suelos como en hojarasca (Graham, 2001). Sin embargo, en Malpelo este grupo no fue tan abundante, quizá porque la cobertura vegetal es muy reducida (<0.02 km2; López-Victoria y Rozo, 2006) y no se forman suelos con hojarasca que puedan albergar este tipo de invertebrados.
Los coleópteros y los isópodos, grupos con mayor abundancia en Malpelo, también son los grupos más diversos y abundantes en las islas Galápagos (Peck, 2005). En Malpelo, el grupo más dominante dentro de Coleoptera fue el de los estafilínidos de la subfamilia Aleocharinae que, además de ser muy abundantes y con frecuencia dominantes en la mayoría de microhábitat del mundo, se encuentran entre los depredadores generalistas más abundantes de las comunidades de hojarasca y del suelo (Navarrete-Heredia et al., 2002).
En cuanto a las hormigas recolectadas en Malpelo se destaca la subfamilia Myrmicinae, que es el grupo más importante de hormigas por su gran número y variabilidad de especies (alrededor de 1000 solo en el neotrópico), y se caracteriza por poseer las especies con el mayor grado de complejidad social conocido de Hymenoptera (Jaffé, 2004). Por su parte Pheidole es catalogado como un género muy rico, además de ser dominante en número de obreras y colonias (Wilson, 2003). Se piensa que, debido a su comportamiento social y forma de forrajeo, tienen una alta incidencia de captura cuando se hace monitoreo de insectos, lo cual parece haberse presentado también en este estudio.
Estructura y redes tróficas
En los bosques continentales e islas grandes que poseen abundante vegetación, la energía disponible y la cantidad de individuos va disminuyendo a medida que sube el nivel trófico (Morin, 2003). Además, una teoría general postula que el número de especies depredadoras no debe exceder el número de especies presa (Levins, 1968), luego los consumidores primarios tienden a concentrar más biomasa que los secundarios, y así sucesivamente. No obstante, existen varias excepciones (Stewart y Levins, 1973), y Malpelo es un claro ejemplo de ello, ya que los depredadores presentaron la mayor riqueza y abundancia, y los saprófagos aportaron casi siete veces la biomasa de los otros gremios tróficos. Kotse y Samways (1999) también encontraron grupos de arañas y estafilínidos como los más diversos entre los invertebrados del suelo y Camero (1999), en un estudio comparativo de la fauna de coleópteros en bosques húmedos de Colombia, encontró una dominancia de la familia Staphylinidae. Otra explicación estaría en el método de captura empleado, ya que se podría tender a capturar especies conspicuas y llamativas, así como aquellas que tienen un comportamiento social o gregario. Sin embargo, Wolda (1975) obtuvo resultados proporcionalmente similares a los de este estudio, a pesar de haber combinado otros métodos para la recolecta de sus datos. Wolda clasificó los estafilínidos como herbívoros debido a su estrecha relación con los musgos, pero esto no aplica para este trabajo, ya que a pesar de que existen algunas especies de Staphylinidae con hábitos saprófagos (no herbívoros), la gran mayoría son depredadoras, en especial la subfamilia Aleocharinae (de igual forma la familia Paederinae, depredadora tanto en su estado inmaduro como adulto; Navarrete- Heredia et al., 2002). Cabe anotar que tanto Aleocharinae como Paederinae fueron encontradas en los sustratos descritos anteriormente por Wolda (1975).
En cuanto a la biomasa de los gremios tróficos, trabajos como el de Lee (1985) sugieren que la alta biomasa de saprófagos en este tipo de estudios está dada por la presencia de lombrices, organismos que tienen un papel muy importante en los procesos físicos, químicos y biológicos del suelo. La presencia de lombrices en Malpelo podría indicar que se están dando estos procesos, y que estarían contribuyendo a la formación de suelo, ya que al ingerir de manera selectiva gran cantidad de material orgánico y mineral, su actividad conduce a la formación de estructuras que contribuyen directa e indirectamente con el aumento de la porosidad, aireación y mejor estabilidad de los suelos (Lee, 1985). A las lombrices se les suma la abundancia de cochinillas, que presentaron pesos relativamente altos respecto a otros invertebrados terrestres (ver más adelante la estructura del microhábitat).
El modelo de red trófica se propone con el ánimo de tratar de explicar algunas de las interacciones que se podrían estar presentando en Malpelo, a partir de lo encontrado en las parcelas muestreadas. En este sentido se presentan a continuación posibles explicaciones para el modelo de red trófica propuesto. Referente a la coexistencia de tantas especies de depredadores respecto al resto de gremios, posiblemente sucede que los depredadores se están comportando como generalistas en vez de especialistas. Es decir que están depredándose entre sí (interacciones intragremiales) y a todos los gremios de niveles inferiores. Esta teoría ya ha sido planteada con base en redes tróficas estudiadas en islas, en particular para el caso de los arácnidos (ver Polis y Hurd, 1995; Polis et al., 1998), y fue también una de las conclusiones a las que llegó Wolda (1975) en su trabajo sobre los invertebrados de Malpelo. Esta tasa de consumo permitiría un balance que haría posible la coexistencia de varias especies de este mismo gremio. Sin embargo, si los depredadores no están consumiendo exclusivamente a los herbívoros, la diversidad de estos tendría que ser mucho mayor, y como este no es el caso, una posible explicación a su baja diversidad debe estar relacionada con la cobertura vegetal, pues esta corresponde a menos del 0.02 % del área total de la isla (López-Victoria y Rozo, 2006), lo que estaría restringiendo la posible diversificación y colonización de nuevas especies de herbívoros, porque no habría nichos disponibles para ellos.
Wolda (1975) propuso que los lagartos podrían estar consumiendo a estos depredadores en algún momento del año a una tasa muy alta, lo cual haría que la competencia disminuyera, de manera que estas especies podrían coexistir. Si esto fuera así, las poblaciones de invertebrados terrestres dependerían de la densidad de lagartos, luego éstos dependerían a su vez de quién controle sus poblaciones. Esta hipótesis es viable si se tiene en cuenta que los invertebrados terrestres en Malpelo no solo son consumidos por los lagartos (principalmente por Anolis agassizi y Phyllodactylus transversalis; López-Victoria, 2006), sino además por algunas especies de aves migratorias (López-Victoria y Estela, 2007), en particular durante las fuertes lluvias, cuando muchos invertebrados salen de sus refugios para no ahogarse, o son simplemente arrastrados por las escorrentías (MLV, obs. pers.).
Es claro que los herbívoros no están manteniendo en forma exclusiva, y mucho menos soportando los requerimientos energéticos de los niveles superiores, aunque se supone que éstos son el segundo eslabón de la gran mayoría de cadenas tróficas basadas en productores primarios (Odum, 1957). Surge entonces la pregunta de cómo puede mantenerse este sistema. La respuesta que se propone en este trabajo es que buena parte de la energía que ingresa al sistema proviene de otra parte, es decir no se genera in situ, combinado con el hecho de que los saprófagos son en este caso los principales agentes "transportadores" de esa energía generada ex situ. Esto explicaría por qué son estos organismos los que están acumulando la mayor cantidad de biomasa entre los macroinvertebrados y porque son los más abundantes. Estos resultados refuerzan la idea de que es el ecosistema marino la fuente principal de esa energía extra. Al respecto ya se ha planteado la importancia de los aportes, provenientes del mar, que hacen las aves marinas al sistema terrestre de Malpelo (Wolda, 1975; Prahl, 1990). Incluso se ha demostrado que esa energía proveniente del mar viene dada en forma de guano, cadáveres de piqueros adultos, polluelos, huevos, restos de pescado y plumas, entre otros (López-Victoria et al., 2009), a lo que habría que sumarle la materia orgánica que se genera in situ, como resultado del metabolismo de la fauna y flora terrestres.
A todo lo anterior se suma que, desde 1986, los infantes de marina apostados en la isla suministran constantemente comida en forma de sobrados (López-Victoria, 2006). Esta carga "extra" de alimento seguramente está siendo aprovechada por saprófagos y especies oportunistas (p. ej., Johngarthia malpilensis; López-Victoria y Werding, 2008), lo que podría significar mayores abundancia y riqueza de invertebrados alrededor de las cabañas, además de una alteración de los ciclos biológicos y reproductivos de algunas especies. En este sentido, cabe la posibilidad de que originalmente hubiese una carencia de especies redundantes o equivalentes ecológicos (Lawton y Brown, 1994), atribuible al grado de aislamiento de la isla, lo que con el tiempo se podría ver afectado de manera considerable, debido al suministro de fuentes de alimento adicionales y a la introducción de especies.
Estructura del microhábitat
El sustrato S3 presentó mayor diversidad, seguramente debido a que se trata del hábitat estructuralmente más complejo. En este sentido, existe una relación directa entre la diversidad y abundancia de especies terrestres y la complejidad estructural del hábitat (Schoener, 1974; Mangee y Sutherland, 1976; M'Closkey, 1978; Armbrecht et al., 2004), ya que un hábitat heterogéneo puede albergar más nichos que un hábitat homogéneo (MacArthur et al., 1962; Levins, 1968). Por otra parte, la gran abundancia de invertebrados presentes en el S1, y la baja diversidad que presenta este microhábitat, sugieren que posiblemente hay dominancia de algunas pocas especies, y que esta dinámica se ve reflejada en los gremios tróficos, pues los nichos de las especies que los representan están definidos por la función de utilización (MacArthur et al., 1962). Sin embargo, las relaciones ecológicas intra e interespecificas también juegan un papel importante en la estructura de las comunidades (Schoener, 1974; Mangee y Sutherland, 1976), y es aquí donde el microhábitat y el grado de complejidad juegan un papel preponderante, pues sus características estructurales afectan la distribución de las especies, permitiendo la diferenciación de nichos y la coexistencia dentro de la comunidad (Schoener, 1974).
El presente es el primer trabajo donde se hace un muestreo utilizando un método estándar e intensivo para recolectar invertebrados terrestres en Malpelo, y el primero en estimar datos de abundancia y biomasa. Por tratarse de un estudio puntual, limitado en el tiempo, el inventario fue incompleto y no refleja las variaciones estacionales a que están sometidos estos organismos. Los resultados obtenidos refuerzan la idea de Malpelo como un sistema subsidiado desde el mar, y en donde los depredadores son el grupo más diverso y abundante.
AGRADECIMIENTOS
Agradecemos al Instituto de Investigaciones Marinas y Costeras-INVEMAR por su apoyo financiero y logístico; al Instituto Colombiano para el Desarrollo de la Ciencia y Tecnología - COLCIENCIAS, en el marco del proyecto #2105-09-13527- 03, por el apoyo financiero; a la UAESPNN por facilitar el trabajo de investigación en Malpelo; a la Fundación Malpelo por su apoyo logístico; a los biólogos Pilar Herrón, Juan Carlos Botello, Catalina Sanabria, Rafael Achury y Jimy Cabra por sus aportes a este trabajo, y al personal de la Armada Nacional por su hospitalidad en la isla y su ayuda en las faenas de campo.
BIBLIOGRAFÍA
1 Álvarez-Rebolledo, M., F. Gast y S. Krieger. 1999. La fauna terrestre de la isla Malpelo. Instituto Humboldt. Biosíntesis, 12: 1-4. [ Links ]
2 Armbrecht, I., I. Perfecto y J. Vandermeer. 2004. Enigmatic biodiversity correlations: Ant diversity responds to diverse resources. Science, 304: 284-286. [ Links ]
3 Borror, D. J., A. Triplehorn y N. F. Johnson. 1992. An introduction to the study of insects. Sexta Edición, Saunders College Publishing, Nueva York. 805 p. [ Links ]
4 Brando, A., H. von Prahl y J. R. Cantera. 1992. Malpelo: isla oceánica de Colombia. Banco de Occidente, Santiago de Cali. 195 p. [ Links ]
5 Buddle, C. M., D. W. Langor, G. R. Pohl y J. R. Spence. 2006. Arthropod responses to harvesting and wildfire: Implications for emulation of natural disturbance in forests and managements. Biol. Conserv., 1 (28): 346-357. [ Links ]
6 Burggren, W. W. y B. R. Mcmahon (Eds.). 1988. Biology of the land crabs. Cambridge University Press, Cambridge. 327 p. [ Links ]
7 Camero, E. 1999. Estudio comparativo de la fauna de coleópteros (Insecta: Coleoptera) en dos ambientes de bosque húmedo tropical colombiano. Rev. Colomb. Entomol., 25 (3): 131-135. [ Links ]
8 Carlquist, S. 1965. Island life: A natural history of the islands of the world. Natural History Press, Nueva York. 200 p. [ Links ]
9 Carlquist, S. 1974. Island biology. Columbia University Press, Nueva York. 250 p. [ Links ]
10 Chamorro, C. y M. Romero. 1990. Biología del suelo. 75-106. En: Aguirre, J. C. y O. Rangel (Eds.). Biota y ecosistemas de Gorgona. Fondo FEN, Bogotá. 303 p. [ Links ]
11 Chase, R. E. 1968. Sea floor topography of the central eastern Pacific Ocean. U.S. Fishand Wildlife Service, BCF Circular 291. 33 p. [ Links ]
12 Colwell, R. K. 2005. User's guide to Estimate S8. Statistical estimation of species richness and shared species from samples. User's guide, ver. 8, and application. http://viceroy.eeb.uconn.edu/estimates. 07/08/2008. [ Links ]
13 De la Fuente, J. A. 1994. Zoología de artrópodos. Interamericana McGraw-Hill, Madrid. 805 p. [ Links ]
14 Fernández, F. 2003. Subfamilia Myrmicinae. 307-330. En: Fernández, F. (Ed.). Introducción a las hormigas de la región Neotropical. Instituto de investigación de recursos biológicos Alexander Von Humboldt, Bogotá. 398 p. [ Links ]
15 Fernández-Palacios, J. M. y C. Morici (Eds.). 2004. Ecología insular. Asociación Española de Ecología Terrestre, Cabildo Insular de la Palma, Santa Cruz de La Palma. 450 p. [ Links ]
16 Fitzpatrick, E. A. 1992. An introduction to soil science. Longman Scientific y Technical, Essex, Inglaterra. 255 p. [ Links ]
17 Flórez, E. 1996. Las arañas del departamento del Valle: Un manual introductorio a su diversidad y clasificación. Inciva-Colciencias, Santiago de Cali. 89 p. [ Links ]
18 Flórez, D. E. y H. Sánchez. 1995. La diversidad de los arácnidos en Colombia, aproximación inicial. 327-372. En: Rangel, O. (Ed.). Colombia, diversidad biótica. Instituto de Ciencias Naturales, Universidad Nacional, Inderena, FES, FEN, Bogotá. 254 p. [ Links ]
19 Gorman, G. C. y T. L. Chorba. 1975. Terrestrial biology of Malpelo island: A historical review. 9-12. En: Graham, J. B. (Ed.). 1975. The biological investigation of Malpelo island, Colombia. Smithsonian Contributions to Zoology No. 176. Smithsonian institution press, Washington, D.C. 98 p. [ Links ]
20 Graham, H. R. 2001. Contributions of oribatid and mesostigmatid soil mites in ecologically based estimates of global species richness. Austral. Ecol., 26: 70-79. [ Links ]
21 Graham, J. B. (Ed.). 1975. Introduction. The biological investigation of Malpelo Island, Colombia. Smithsonian Contr. Zool., 176, Washington, D.C. 98 p. [ Links ]
22 Halaj, J., D. W. Ross y A. R. Moldenke. 1998. Habitat structure and prey availability as predictors of the abundance community organization of spiders in western Oregon forest canopies. J. Arachnol., 26: 203-220. [ Links ]
23 Hogue, C. L. y S. E. Miller. 1981. Entomofauna of Cocos Island, Costa Rica. Atoll Res. Bull., 250:1-29. [ Links ]
24 Jaffé, C. K. 2004. El mundo de las hormigas. Equinoccio Ediciones de la Universidad Simón Bolívar, Valle de Sartenejal, Baruta. 148 p. [ Links ]
25 Kotse, D. J y M. J. Samways. 1999. Support for the multi-taxa approach in biodiversity assessment, as shown by epigaeic invertebrates in an Afromontane forest fragment archipelago. J. Insect. Conserv., 3: 125-143. [ Links ]
26 Lawton, J. H. y V. K. Brown. 1994. Redundancy in ecosystems. 255-270. En: Schulze, E. D. y H. A. Mooney. (Eds.). Biodiversity and Ecosystem Function. Springer-Verlag, Nueva York. 521 p. [ Links ]
27 Lee, K. E. 1985. Hearthworms: their ecology and relations with soils and land use. Academic Press, Sidney. 411 p. [ Links ]
28 Levins, R. 1968. Evolution in changing environments. Princeton University Press, Princeton, New Jersey. 120 p. [ Links ]
29 López-Victoria, M. 2006. Los lagartos de Malpelo (Colombia): Aspectos sobre su ecología y amenazas. Caldasia, 28 (1): 129-134. [ Links ]
30 López-Victoria, M. 2009. Island ecology: trophic and breeding behaviour of land crabs and seabirds determined the terrestrial community of Malpelo. Disertación de Doctorado, Facultad de Biología y Química, Universidad Justus-Liebig, Giessen, Alemania. 144 p. [ Links ]
31 López-Victoria, M. y F. A. Estela. 2007. Una lista anotada de las aves de la isla Malpelo. Ornitología Col., 5: 40-53. [ Links ]
32 López-Victoria, M. y D. M. Rozo. 2006. Model-based geomorphology of Malpelo island and spatial distribution of breeding seabirds. Bol. Invest. Mar. Cost., 35: 111-131. [ Links ]
33 López-Victoria, M. y B. Werding. 2008. Ecology of the endemic land crab Johngarthia malpilensis (Decapoda: Brachyura: Gecarcinidae), a poorly known species from the Tropical Eastern Pacific. Pac. Sci., 62: 483-493. [ Links ]
34 López-Victoria, M., V. Wolters y B. Werding. 2009. Nazca Booby (Sula granti) inputs maintain the terrestrial food web of Malpelo Island. J. Ornithol., 150: 865-870. [ Links ]
35 MacArthur, R. H. y E. O. Wilson. 1967. Island biogeography. Princenton University Press, Princeton, Nueva York. 224 p. [ Links ]
36 MacArthur, R. H., J. W. MacArthur y J. Preer. 1962. On bird species diversity. II. Predictions of bird census from habitat measurements. Am. Nat., 96: 167-164. [ Links ]
37 Mangee, B. A. y J. P Sutherland. 1976. Species diversity gradients: Synthesis of the roles of predation, competition, and temporal heterogeity. Am. Nat., 110: 351-369. [ Links ]
38 M'Closkey, R. T. 1978. Niche separation and assembly in four species of Sonoran deserts rodents. Am. Nat., 112: 683-694. [ Links ]
39 Morin, P. J. 2003. Comunnity ecology and the genetics of interacting species. Ecology, 84 (3): 577-580. [ Links ]
40 Navarrete-Heredia, J. L., A. F. Newton, M. K. Thayer, J. S. Ashe y D. S. Chandler. 2002. Guía ilustrada para los géneros de Staphylinidae (Coleoptera) de México. Universidad de Guadalajara y CONABIO, México, D. F. 401 p. [ Links ]
41 Nentwig, W. 1993. Spiders of Panama. Fauna y flora Handbook. The Sandhill Crane. Inc, Leiden Press, Gainesville, Estados Unidos. 274 p. [ Links ]
42 Odum, H. T. 1957. Trophics estructure and productivity of Silver Springs. Florida. Ecol. Monogr., 27: 55-112. [ Links ]
43 Peck, S. B. 2005. The beetles of the Galápagos Islands, Ecuador, evolution, ecology, and diversity (Insecta: Coleoptera). NRC Research Press, Ottawa. 302 p. [ Links ]
44 Peng, X. J., I. M. Tso y S. Q. Li. 2002. Five new and four newly recorded species of jumping spiders from Taiwan (Araneae: Salticidae). Zool. Stud., 41 (1): 1-12. [ Links ]
45 Polis, G. A. y S. D. Hurd. 1995. Extraordinarily high spider densities on islands: flow of energy from the marine to terrestrial food webs and the absence of predation. PNAS, 92: 4382-4386. [ Links ]
46 Polis, G. A., S. D. Hurd, C. T. Jackson y F. Sánchez-Piñero. 1998. Multifactor population limitation: variable spatial and temporal control of spiders on Gulf of California islands. Ecology, 79: 490-502. [ Links ]
47 Prahl, H. von. 1990. Malpelo la roca viviente. Editorial Presencia, FEN, Bogotá. 105 p. [ Links ]
48 Rico, G. A., J. P. A. Beltrán, A. D. Álvarez y E. Flórez. 2005. Diversidad de arañas (Arachnida: Araneae) en el Parque Nacional Natural isla Gorgona, Pacífico colombiano. Biota Geotrópica, 5: 99-110. [ Links ]
49 Rodrigues, C. A., I. Muñoz, N. Bonada, A. Gaudes y S. Tomanova. 2009. La biota de los ríos: los invertebrados. 253-270. En: Elosegi, A. y S. Sabater (Eds.). Conceptos y técnicas en ecología fluvial. Fundación BBVA, Bilbao. 470 p. [ Links ]
50 Ruppert, E. E. y R. D. Barnes. 1996. Zoología de los invertebrados. McGraw-Hill Interamericana, México D.F. 1114 p. [ Links ]
51 Schoener, T. W. 1974. Resourse partitioning in ecological communities. Science, 185: 27-39. [ Links ]
52 Silva, D. 1996. Species composition and community structure of Peruvian rainforest spiders: a case study from a seasonally inundated forest along the Samiria River. Rev. Suisse Zool., 597-610. [ Links ]
53 Silva, D. y J. A. Coddington. 1996. Spiders of Pakitza (Madre de Dios, Perú): species richness and notes on community structure. 253-311 En: Wilson, D. E. y A. Sandoval (Eds.). The biodiversity of Southeastern Perú. Smithsonian Institution, Washington D. C. 679 p. [ Links ]
54 Stead, J. A. 1975. Field observations on the geology of Malpelo island. Smithsonian Contr. Zool., 176: 17-20. [ Links ]
55 Stewart, F. M. y R. Levins. 1973. Partitioning of resources and the outcome of interspecific competition: A model and some general considerations. Am. Nat., 107:171-198. [ Links ]
56 Turnbull, A. L. 1973. Ecology of the true spiders. Annu. Rev. Entomol., 18: 305-348. [ Links ]
57 Ubick, D. P., P. Paquin, P. E. Cushing, y V. Roth. 2005. Spiders of North America: An identification mannual. American Arachnological Society, Portland, Estados Unidos. 377 p. [ Links ]
58 Ulrich, W. 2004. Soil living parasitic Hymenoptera: Comparison between a forest and an open landscape habitat. Pedobiologia, 48: 59-69. [ Links ]
59 Villareal, H., M. Álvarez, S. Córdoba, F. Escobar, G. Fagua, F. Gast, H. Mendoza, M. Ospina y A. M. Umaña. 2004. Manual de métodos para el desarrollo de inventarios de biodiversidad, Programa de inventarios de biodiversidad. Instituto de recursos biológicos Alexander von Humboldt, Bogotá. 236 p. [ Links ]
60 Whittaker, R. J. y J. M. Fernández-Palacios. 2007. Island Biogeography: ecology, evolution and conservation. Segunda edición, Oxford University Press, Nueva York. 401 p. [ Links ]
61 Wilson, E. O. 2003. La hiperdiversidad como fenómeno real: el caso de Pheidole. 363-369. En: Fernández, F. (Ed.). 2003. Introducción a las hormigas de la región neotropical. Instituto de Investigación de Recursos Biológicos Alexander Von Humboldt, Bogotá. 424 p. [ Links ]
62 Wolda, H. 1975. The ecosystem on Malpelo island. Smithsonian Contr. Zool., 176: 22-26. [ Links ]
FECHA DE RECEPCIÓN: 21/09/2010 FECHA DE ACEPTACIÓN: 12/10/2011
*Contribución No. 1097 del Instituto de Investigaciones Marinas y Costeras-INVEMAR.

