










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.41 no.1 Santa Marta Jan./June 2012
DIFERENCIACIÓN MORFOLÓGICA DE LAS ESPECIES DE PIANGUA ANADARA TUBERCULOSA Y ANADARA SIMILIS (ARCIDAE) EN DIFERENTES BOSQUES DE MANGLAR A LO LARGO DE LA COSTA PACÍFICA COLOMBIANA MEDIANTE MORFOMETRÍA GEOMÉTRICA*
MORPHOLOGICAL DIFERENTIATION OF THE SPECIES OF PIANGUA, ANADARA TUBERCULOSA AND ANADARA SIMILIS (ARCIDAE), IN MANGROVE FORESTS THROUGHOUT THE PACIFIC COAST OF COLOMBIA USING GEOMETRIC MORPHOMETRICS
Jennee Lorena Cano-Otalvaro1, Óscar Enrique Murrillo-García2, Jaime Ricardo Cantera-Kintz2 y Diego Luis Gil-Agudelo1,3
1 Instituto de Investigaciones Marinas y Costeras, INVEMAR. Sede Pacífico. CIAT – Parque Agronatura. Kilómetro 17 recta Cali-Palmira. jenkaot@gmail.com.
2 Universidad del Valle, Facultad de Ciencias, Programa de Biología, Sede Meléndez. Calle 13 No. 100-00, Cali, Colombia. omurillo@univalle.edu.co (OEMG); jcantera@univalle.edu.co (JRCK).
3 Empresa Colombiana de Petróleos, Ecopetrol S.A. Bogotá D.C. Edificio principal, Carrera 13 No. 36-24. diego.gil@ecopetrol.com.co (dirección actual).
RESUMEN
En el presente trabajo se caracterizaron morfométricamente las especies de piangua, Anadara tuberculosa (Sowerby) y A. similis (Adams) mediante el uso de la morfometría geométrica. Se utilizaron conchas de ambas especies colectadas en nueve zonas diferentes del litoral Pacífico colombiano durante el año 2009 en el marco de un proyecto sobre potencial productivo de las poblaciones naturales de piangua. La valva izquierda de los organismos se utilizó para localizar siete puntos homólogos y evaluar la configuración geométrica de la concha. Las pianguas fueron agrupadas según la cobertura vegetal de la zona en la que fueron encontradas. Para ambas especies, la comparación intraespecífica mostró que la configuración de la valva izquierda no se ve afectada por la zona que habitan, contrario al tamaño centroide que presentó variaciones significativas entre las zonas estudiadas. Las diferencias debidas al tamaño no son suficientes para afirmar la existencia de diferentes poblaciones ya que estas variaciones pueden ser debidas a la presión pesquera que se ejerce sobre este recurso de manera diferencial en la región, causando el incremento porcentual de tamaños menores. Nuestros resultados muestran que las corrientes del océano Pacífico podrían estar favoreciendo el flujo genético entre los individuos de estas especies y limitando la posibilidad de una evolución independiente de poblaciones. Sin embargo es importante realizar estudios genéticos y de dispersión larval de estas especies a lo largo del litoral Pacífico para corroborar estos hallazgos.
PALABRAS CLAVE: Anadara tuberculosa, Anadara similis, Morfometría geométrica, Costa pacífica colombiana.
ABSTRACT
The morphological characterization of two piangua species Anadara tuberculosa (Sowerby) and Anadara similis (Adams) was performed using geometric morphometric. Shells of both species were collected in nine different areas of the Colombian Pacific coast during 2009 as part of a project on the natural populations of piangua. The left valve of the shell was used to draw seven landmarks and evaluate the geometric configuration of the shell. Pianguas were grouped based on the mangrove cover of the localities where they were collected. Intra-specific comparison showed that areas where the organisms were collected do not affect valve configuration. Contrarily, centroide size showed significant variation in organism collected in different zones. Differences due to size are not sufficient to determine the existence of different populations, mainly because these differences may happen due to differences in fishery patterns increasing the ratio of smaller sizes. Pacific Ocean current might favor the genetic flux of organisms of both species, limiting the possibility of evolution of independent populations. It is important to complement this study with other investigations in genetics and larval dispersion in order to corroborate our findings.
KEY WORDS: Anadara tuberculosa, Anadara similis, Geometric morphometric, Pacific coast of Colombia.
INTRODUCCIÓN
Las especies de piangua Anadara tuberculosa (Sowerby) y A. similis (Adams), son moluscos bivalvos que se encuentran asociados a las raíces del mangle, especialmente Rhizophora spp., en el Pacífico americano (Fischer et al., 1995). En Colombia son los bivalvos más explotados en el litoral Pacífico y de ellos se benefician las comunidades que habitan esta región. La recolección de pianguas ha venido siendo desarrollada habitualmente por mujeres sin importar la edad, siendo común que las ancianas y adultas, generalmente cabezas de hogar, colecten los individuos del sedimento que rodea las raíces de los mangles para alimentar a sus familias o comercializar estos moluscos en el mercado. Actualmente el recurso, además de ser utilizado para consumo y comercio local, está siendo llevado al vecino país del Ecuador, produciéndose así un aumento en la demanda y explotación de estos animales, lo cual ha causado la incorporación de los hombres a esta labor, especialmente cuando las otras actividades como la pesca resultan poco rentables (Espinosa et al., 2009; INVEMAR et al., 2009), lo que ha aumentado sus niveles de explotación llevándola a ser incluida en el libro rojo de invertebrados marinos de Colombia en categoría de vulnerable (Ardila et al., 2002) por la alta probabilidad de sobreexplotación. Se hace necesario entonces el establecimiento de medidas que garanticen la conservación y uso sostenible de estas especies. Una de estas medidas complementadas es el establecimiento de Áreas Marinas Protegidas (AMP); sin embargo, para un efectivo establecimiento de este tipo de áreas es importante conocer la distribución de las poblaciones y la posible detección de poblaciones fuentes y sumideros para hacerlas más efectivas (Fernández y Castilla, 2005; Pomeroy et al., 2006).
La morfometría geométrica permite comparar los tamaños entre individuos y cuantificar sus aspectos, perfiles, geometrías, conformaciones y formas de manera estable, libre de variaciones fisiológicas del tamaño (Dujardin, 2002c), lo que la hace una herramienta adecuada para el análisis poblacional y la clasificación taxonómica de las especies mediante el uso de coordenadas cartesianas (Langerhans et al., 2003; Medel et al., 2003; Rohlf y Bookstein, 2003; Aguirre et al., 2006; Ibáñez et al., 2007; Calle et al., 2008). En moluscos bivalvos se ha utilizado este método para el análisis de poblaciones de Corbicula spp. en dos estuarios de Portugal (Sousa et al., 2007). En Argentina se ha usado para caracterizar y distinguir la metapoblación de la almeja rayada, Ameghinomya antiqua, de stocks fenotípicos al norte de la Patagonia (Márquez et al., 2010) y también se ha usado en estudios paleontológicos y taxonómicos en Brachidontes (Mytilidae) (Aguirre et al., 2006). Para las especies Anadara tuberculosa y A. similis no se encuentran registros. El presente trabajo busca determinar la existencia de diferencias poblacionales de A. tuberculosa y A. similis en distintos lugares de la costa Pacífica colombiana mediante el uso de morfometría geométrica de modo que sirva para el futuro diseño de AMP u otro tipo de medidas de conservación que permitan la preservación y el uso de estas importantes especies.
MATERIALES Y MÉTODOS
Se analizaron conchas de individuos de A. tuberculosa y A. similis recolectados en diferentes localidades de la costa pacífica colombiana (Figura 1) durante 2009. El número de conchas de cada especie usada para el análisis y la cobertura vegetal de cada una de las áreas de recolección se muestra en la Tabla1.
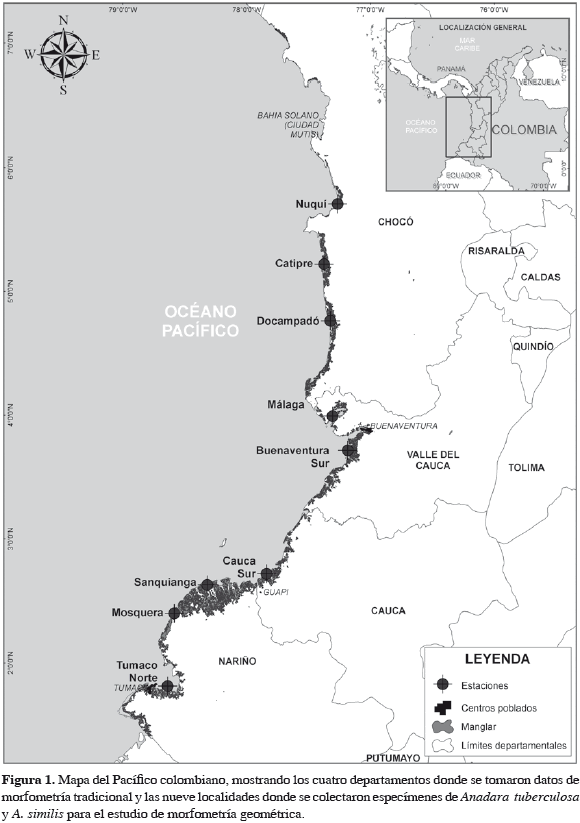
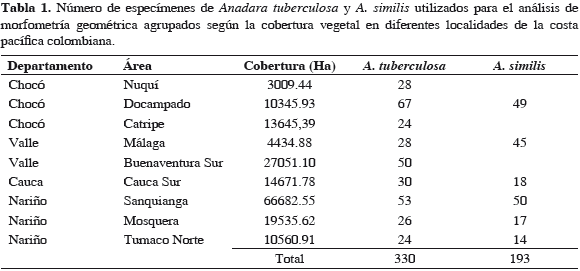
Para estudiar la configuración geométrica de la concha se utilizó la valva izquierda, que fue escaneada (lado cóncavo hacia arriba) con la ayuda de un escáner Hewlett Packard Scanjet 4070, a una resolución de 600 ppp. Posteriormente en cada concha se seleccionaron siete puntos homólogos (Figura 2), siguiendo los criterios establecidos en Zelditch et al. (2004). El programa tpsUtil, versión 1.44 (Rohlf, 2009) se utilizó para crear un archivo de imágenes Tps en las que se ubicaron los puntos homólogos con ayuda del programa tpsDig2, versión 2.12 (Rohlf, 2008a). Para identificar si los puntos homólogos en los especímenes fueron marcados correctamente, se realizó un análisis de repetitividad, que consistió en seleccionar aleatoriamente una submuestra de 30 individuos, a los que se les había digitalizado los puntos homólogos para volverlos a digitalizar, utilizando los programas TpsUtil y TpsDig. Con los programas Tetwin (Dujardin, 2002a), VARwin, versión 1.7 (Dujardin, 2003) y MOGwin, versión 0 79 (Dujardin, 2007) se obtuvieron los valores de repetitividad para los componentes X y Y de cada punto homólogo, y en Excel se calculó la repetitividad promedio de X y Y, cada punto homólogo y del análisis total, tanto de A. tuberculosa como de A. similis. A partir del análisis generalizado de Procrustes realizado con el programa tpsRelw, versión 1.46 (Rohlf, 2008b), se obtuvieron dos matrices que contenían toda la información sobre el tamaño y la configuración de la valva izquierda de las pianguas: la matriz del tamaño ("centroid size"), a la que se le realizaron análisis estadísticos (Kruskall-Wallis) para determinar si existían diferencias significativas entre los tamaños con PAST versión 1.79 (Hammer et al., 2001), y la matriz ponderada ("weight matrix"), a la que se le realizó un análisis de componentes principales (ACP) mediante el uso del programa de acceso libre BACwin (Dujardin, 2002b).

El tipo de crecimiento de las especies se determinó mediante un análisis de correlación utilizando el programa Statistica (StatSoft Inc, 2010). Donde el tamaño centroide fue usado como variable independiente, ya que es considerado un buen descriptor de forma (Bookstein, 1996), y como variables dependientes los componentes uniformes de forma UniX y UniY. El valor del índice de correlación (valor p) y la significancia indicaron el cambio en la forma, siendo alométrico cuando se encontraron índices de correlación altos (>0.5) y probabilidades significativas (<0.05) (Bermúdez, 2005).
RESULTADOS
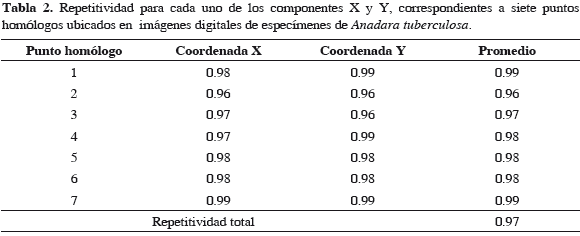
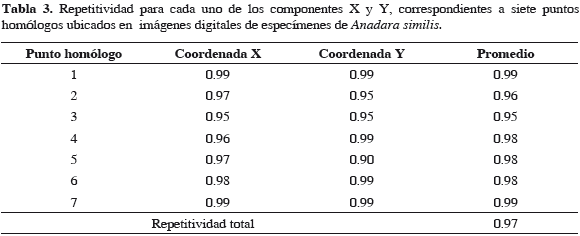
El análisis de repetitividad mostró que los puntos homólogos en las especies fueron ubicados con una precisión del 97 %. Las Tablas 2 y 3 muestran el valor de repetitividad (entre 0 y 1) para los componentes X y Y de los siete puntos homólogos, la repetitividad promedio para cada punto homólogo y la repetitividad total de cada una de las especies estudiadas.
Anadara tuberculosa
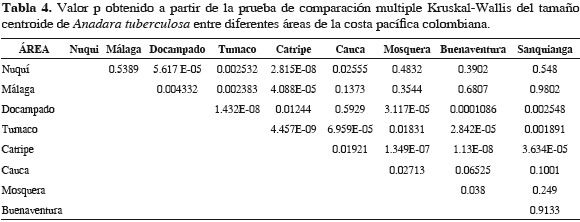
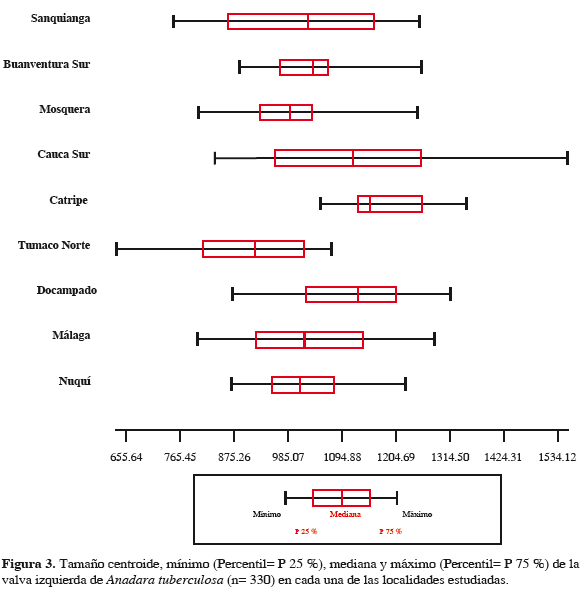
Se encontraron diferencias significativas en el tamaño de la valva izquierda de A. tuberculosa entre las localidades (Tabla 4). Los individuos más grandes se encontraron en Catripe (Chocó) y los más pequeños en el área norte de Tumaco (Figura 3). La distribución de los especímenes de A. tuberculosa a lo largo de los dos primeros factores, derivados del análisis de componente principales (CP 1 y CP 2), de la variación de la forma de la valva izquierda en cada una de las nueve localidades estudiadas, evidenció que los polígonos que delimitan las áreas que agrupan a todos los individuos de cada una de las localidades comparten áreas entre ellos, por lo que es difícil separarlas (Figura 4). La correlación entre el tamaño centroide y cada uno de los componentes de la forma UniX y UniY, muestran que no existen relación entre el tamaño y los descriptores de forma de los especímenes (centroide vs UniX: r= 0.0913, r2= 0.008336, p= 0.0978; centroide vs UniY: r= -0.007176, r2= 0.000052, p= 0.8967) lo que hace suponer crecimiento isométrico en la especie.
Anadara similis
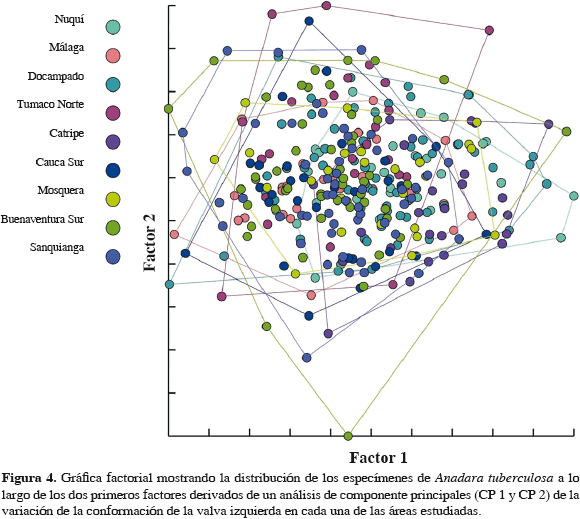
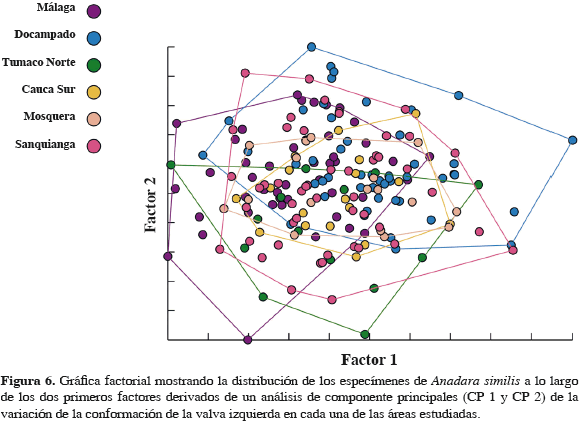
Los organismos de las diferentes áreas presentaron diferencias significativas en sus tamaños (Tabla 5), siendo los individuos muestreados en Docampado (Chocó) los más grandes mientras que los del Norte de Tumaco los más pequeños (Figura 5). El análisis multivariado mostró que la conformación de la valva izquierda de A. similis no difiere intraespecíficamente entre las seis zonas estudiadas (Figura 6). Los individuos se distribuyeron de manera similar en los dos primeros factores lo que señala una gran similitud en la conformación de la valva izquierda de esta especie entre las zonas. La correlación entre el tamaño centroide y cada uno de los componentes de la forma muestran una relación significativa no muy fuerte con los componentes de la forma (centroide vs UniX: r = 0.273977, r2= 0.075053, p= 0.000115; centroide vs UniY: r = 0.145421, r2= 0.021147, p= 0.043605), lo que hace suponer una tendencia hacia el crecimiento alométrico en A. similis.

DISCUSIÓN
La variación en la conformación de la valva izquierda de la concha de moluscos bivalvos ha sido utilizada para identificar y separar grupos intraespecíficamente de otras especies (Sousa et al., 2007; Márquez et al., 2010). Debido a esto, en el presente estudio se utilizó la valva izquierda de los especímenes recolectados, en la que se ubicaron siete puntos homólogos (Figura 2) los cuales presentaron una alta repetitividad (97 %), lo que los hizo fácilmente reproducibles de un espécimen a otro (Tabla 2 y 3).
Siendo Anadara tuberculosa y A. similis especies bastante cercanas, lo que se esperaría es que ambas tengan el mismo tipo de crecimiento; sin embargo, los resultados obtenidos no evidencian esto. La relación entre el tamaño de los especímenes (tamaño centroide) y los descriptores de forma (UniX y UniY) de A. tuberculosa no fue significativa, lo que sugiere que la forma de la valva izquierda no cambia a medida que la piangua aumenta de tamaño siguiendo un crecimiento de tipo isométrico. En A. similis la relación entre el tamaño y los componentes UniX y UniY de la forma de la valva izquierda fue significativa, lo que indica que esta especie tiene un crecimiento de tipo alométrico modificando su forma a medida que crece.
El tamaño centroide de la valva izquierda de las especies de pianguas presentó diferencias significativas entre las áreas formándose diferentes grupos. En el caso de A. tuberculosa, Catripe agrupó las pianguas de mayor tamaño, diferenciándose significativamente esta localidad de las demás estudiadas. Las pianguas de menor tamaño se encontraron en Tumaco Norte y formaron un grupo significativamente diferente al resto de las localidades que presentaron tamaños intermedios y entre las cuales no es posible evidenciar grupos claramente diferentes (Tabla 4 y Figura 3). Por su parte, A. similis formó dos grupos que presentan diferencias significativas entre ellos. El primero agrupó los mayores tamaños que se encontraron en Mosquera, Docampado y Sanquianga, estas dos últimas localidades difieren a su vez entre ellas significativamente, debido posiblemente a que Docampado, a pesar de tener un ámbito de tamaño más estrecho, registró los especímenes de mayores tamaños entre todas las áreas, mientras que en los de Sanquianga se presentaron valores intermedios de tamaño. El segundo grupo está conformado por los individuos provenientes de Cauca Sur, Tumaco Norte y Málaga, donde se encontraron los menores tamaños, registrándose diferencias significativas entre Tumaco Norte y Málaga, debidas quizá a que los tamaños de los especímenes de Málaga son mayores que los de Tumaco Norte, que fue donde se encontraron los menores tamaños entre todas las áreas (Tabla 5 y Figura 5).
Las diferencias de tamaño encontradas no son suficientes para determinar la existencia de diferentes poblaciones de pianguas, ya que estas variaciones pueden deberse a factores exógenos como salinidad, temperatura o energía de agua que afectan el crecimiento de los bivalvos (Silva y Bonilla, 2001; Aguirre et al., 2006) u otras condiciones existentes en los estuarios que habitan. Como ejemplo se puede mencionar el trabajo de Sousa et al. (2007) que muestra que conchas más alargadas de Corbicula fluminea se deben a condiciones de hacinamiento y bajas tasas de crecimiento de esta especie y no a factores endógenos (genéticos).
En mayor medida las diferencias de tamaño encontradas pueden estar influenciadas por la presión pesquera que se ejerce sobre el recurso piangua, ya que este sucede de forma diferencial en el Pacífico colombiano. Así, por ejemplo, en la ensenada de Tumaco, incluyendo la localidad Tumaco norte, se encontraron los individuos más pequeños de ambas especies, y al mismo tiempo se obtiene cerca del 60 % de la producción de la piangua en el país (Borda y Cruz, 2003), de los cuales el 55 % de los organismos comercializados presentan una talla menor a la talla mínima establecida de 5 cm, lo cual podría explicar las menores tallas de los individuos en esta zona.
El análisis de componentes principales evidenció que la forma de la valva izquierda de los especímenes no varía entre las áreas. Tal como se puede visualizar en el mapa factorial tanto para A. tuberculosa (Figura 4) como para A. similis (Figura 6), la conformación de la concha se distribuye de manera similar en el espacio multivariado. Esto mostraría que no existen diferencias poblacionales entre los individuos de las especies evaluadas. Arbeláez (2003) encontró que las poblaciones de Rhizophora mangle entre el sur de Colombia y el área de bahía Málaga no poseen una marcada diferenciación genética, mostrando como la Corriente Colombia en el Pacífico colombiano es de gran importancia para el transporte de materiales genéticos de esta y posiblemente de otras especies.
Las corrientes marinas presentes en el Pacífico colombiano, tales como la corriente Ecuatorial del Norte, la corriente del golfo de Panamá, la corriente Ecuatorial del Sur y especialmente la corriente de Colombia, que se mueve en sentido contrario a las manecillas del reloj con dirección predominante NNE y velocidades de hasta 150 cm/seg en los meses de diciembre a abril (Prahl et al., 1990), favorecerían la migración larval y el intercambio genético entre las dos especies de pianguas, limitando la probabilidad de una evolución independiente entre las diferentes áreas estudiadas. Es importante entonces realizar estudios genéticos y de dispersión larval en las especies de A. tuberculosa y A. similis, a lo largo del litoral Pacífico colombiano, que complementen la información aquí presentada.
AGRADECIMIENTOS
Agradecemos especialmente al proyecto "Potencial productivo de las poblaciones naturales de la piangua Anadara tuberculosa y A. similis dentro de una perspectiva espacio-temporal en la costa pacífica colombiana" y a cada una de las personas que hicieron posible el desarrollo de este, especialmente a Carlos Lucero, William Gualteros, Silvana Espinosa, Alex Manjarrés y María Fernanda Delgado, a la Universidad del Valle, el Instituto de Investigaciones Marinas y Costeras "José Benito Vives de Andréis"-INVEMAR, Colciencias, la Dirección Territorial Sur Occidental, UAESPNN, la Asociación de Concheras de Nariño, el WWF Colombia y al Ministerio de Agricultura y Desarrollo Rural. Igualmente damos las gracias al Doctor Joao Muñoz, profesor de la Universidad Nacional, y a los estudiantes de Maestría en esta misma institución Ludwig Jiménez y Jesualdo Fuentes.
BIBLIOGRAFÍA
1 Aguirre, L., S. Pérez y Y. Sirch. 2006. Morphological variability of Brachidontes swainson (Bivalvia, Mytilidae) in the marine Quaternary of Argentina (SW Atlantic). Palaeogeography, Palaeoclimatology, Palaeoecology, 239: 100-125. [ Links ]
2 Arbeláez, E. 2003. Análisis de diversidad y la estructura genética de dos poblaciones naturales del árbol de manglar Rhizophora mangle (Rhizophoraceae) de la costa Pacífica colombiana mediante el uso de marcadores moleculares microsatélites de ADN. Tesis de Biología, Univ. Valle, Cali. 98 p. [ Links ]
3 Ardila, N., G. Navas y J. Reyes. 2002. Libro rojo de invertebrados marinos de Colombia. Serie libros rojos de especies amenazadas de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt y Ministerio del Medio Ambiente, Bogotá. 136 p. [ Links ]
4 Bermúdez, A. 2005. Caracterización morfológica de las poblaciones de Agononida longipes y Munida flinti (Crustacea: Decapoda: Galatheidae) en el mar Caribe colombiano. Tesis de Maestría, Univ. Nacional de Colombia, Bogotá. 100 p. [ Links ]
5 Bookstein, F. 1996. Landmark methods for forms without landmarks: morphometrics of group differences in outline shape. Medical Image Anal., 1 (3): 225-243. [ Links ]
6 Borda, C. y R. Cruz. 2003. Estado de explotación y pronóstico de la pesquería de Anadara tuberculosa (Sowerby, 1833) en el Pacífico colombiano. Rev. Invest. Mar., 24 (3): 221-230. [ Links ]
7 Calle, D., M. Quiñones, H. Erazo y N. Jaramillo. 2008. Discriminación por morfometría geométrica de once especies de Anopheles (Nyssorhynchus) presentes en Colombia. Biomédica, 28: 371-385. [ Links ]
8 Dujardin, J. 2002a. TET, version 2. Institut de Reacherches pour le Développement (IRD). Montpellier. Francia. http://www.mpl.ird.fr/morphometrics/. 01/09/2010. [ Links ]
9 Dujardin, J. 2002b. BAC, version 2. Institut de Reacherches pour le Développement (IRD). Montpellier. Francia. http://www.mpl.ird.fr/morphometrics/. 01/09/2010. [ Links ]
10 Dujardin, J. P. 2002c. Introducción a la morfometría. (Con énfasis en Triatominae y Phlebotominae). IRD (Institut de Recherches pour le Développement, Paris, France). http://eclat.fcien.edu.uy/espaniol/Libros.htm. 01/09/2010. [ Links ]
11 Dujardin, J. 2003. VARwin, versión 1.7. Institut de Reacherches pour le Développement (IRD). Montpellier. Francia. http://www.mpl.ird.fr/morphometrics/. 01/09/2010. [ Links ]
12 Dujardin, J. 2007. MOGwin, versión 2. Institut de Reacherches pour le Développement (IRD). Montpellier. Francia. http://www.mpl.ird.fr/morphometrics/. 01/09/2010. [ Links ]
13 Espinosa, S., D. L. Gil-Agudelo, C. Candelo y L. A. Zapata. 2009. Las piangüeras en la Costa pacífica colombiana: investigación participativa para la conservación de la piangua y la actividad económica de este recurso biológico. 159-167. En: Vicepresidencia de la República, Comisión Colombiana del Océano, Observatorio del Pacífico colombiano (Eds.). Los pueblos del mar. Comisión Colombiana del Oceáno, Bogotá. 167 p. [ Links ]
14 Fernández, M. y J. C. Castilla. 2005. Marine conservation in Chile: Historical perspective, lessons, and challenges. Cons. Biol., 19 (6): 1752-1762. [ Links ]
15 Fischer, W., F. Krupp, W. Schneider, C. Sommer, K. E. Carpenter y V. Niem. 1995. Guía FAO para la identificación de especies para los fines de la pesca Pacífico centro-oriental: plantas e invertebrados. Volumen I, FAO, Roma. 646 p. [ Links ]
16 Hammer, O., D. Harper y P. Ryan.2001. PAST: Paleontological statistic software package for education and data analysis. http://paleo-electronica.org/2001_1/past/issue1_01.htm. 01/09/2010. [ Links ]
17 Ibáñez, A., L. Cowx y P. O'Higgins. 2007. Geometric morphometric analysis of fish scales for identifyting genera, species, and local populations within the Mugilidae. Can. J. Fish. Aq. Sci., 64: 1091-1100. [ Links ]
18 INVEMAR, Asconar, WWF Colombia, PNN Sanquianga y Universidad del Valle. 2009. Segundo Boletín: Piangüando. Ministerio de Agricultura y Desarrollo Rural. www.invemar.org.Co/redcostera1/INVEMAR/docs/7177BOLETIN_PIANGUANDO_2.pdf. 01/02/2010. [ Links ]
19 Langerhans, B., C. Layman, A. Langerhans y T. Dewitt. 2003. Habitat-associated morphological divergence in two Neotropical fish species. Biological Journal of the Society, 80: 689-698. [ Links ]
20 Márquez, F., J. Robledo, G. Escati y S. Van der Molen. 2010. Use of different geometric morphometrics tools for the discrimination of phenotypic stocks of the striped clam Ameghinomya antiqua (Veneridae) in north Patagonia, Argentina. Fish. Res., 101: 127-131. [ Links ]
21 Medel, R., C. Botto-Mahan y M. Kalin-Arroyo. 2003. Pollinator mediated selection on the nectar guide phenotype in the Andean monkey flower, Mimulus luteus. Ecology, 84 (7): 1721-1732. [ Links ]
22 Pomeroy, R. S., J. E. Parks y L. M. Watson. 2006. Cómo evaluar una AMP. Manual de indicadores naturales y sociales para evaluar la efectividad de la gestión de áreas marinas protegidas. UICN, Gland y Cambridge. 216 p. [ Links ]
23 Prahl., H.v., J. R. Cantera y R. Contreras. 1990. Manglares y hombres del Pacífico colombiano. Fondo FEN-Colciencias, Bogotá. 193 p. [ Links ]
24 Rohlf, J. 2008a. tpsDig, version 2.12. Stony Brook, NY: Department of Ecology and Evolution, State University of New York at Stony Brook. http://life.bio.sunysb.edu/morph. 01/09/2010. [ Links ]
25 Rohlf, J. 2008b. tpsRelw, version 1.46. Stony Brook, NY: Department of Ecology and Evolution, State University of New York at Stony Brook. http://life.bio.sunysb.edu/morph. 01/09/2010. [ Links ]
26 Rohlf, J. 2009. tpsUtil, version 1.44. Stony Brook, NY: Department of Ecology and Evolution, State University of New York at Stony Brook. http://life.bio.sunysb.edu/morph. 01/09/2010. [ Links ]
27 Rohlf, J. y F. Bookstein. 2003. Computing the uniform component of shape variation. Syst. Biol., 52 (1): 66-69. [ Links ]
28 Silva, M. y R. Bonilla. 2001. Abundancia y morfometría de Anadara tuberculosa y A. similis (Mollusca: Bivalvia) en el manglar de Purruja, Golfo Dulce, Costa Rica. Rev. Biol. Trop., 49 (2): 315-320. [ Links ]
29 Sousa, R., R. Freire, M. Rufino, J. Méndez, M. Gaspar, C. Antunes y L. Guilhermino. 2007. Genetic and shell morphological variability of the invasive bivalve Corbicula fluminea (Müller, 1774) in two Portuguese estuaries. Est., Coast. Shelf Sci., 74: 166-174. [ Links ]
30 StatSoft, Inc. 2010. Electronic Statistics Textbook. Tulsa, OK: StatSoft. WEB: http://www.statsoft.com/textbook/. 01/09/2010. [ Links ]
31 Zelditch, M. L., D. L. Swiderski, H. D. Sheets y W. L. Fink. 2004. Geometric morphometrics for biologists. Elsevier Academic Press, Amsterdam. 443p. [ Links ]
FECHA DE RECEPCIÓN: 22/06/2011 FECHA DE ACEPTACIÓN: 09/02/2012
*Contribución No. 1100 del Instituto de Investigaciones Marinas y Costeras-INVEMAR.
