










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.41 no.2 Santa Marta July/Dec. 2012
EL COLAPSO DE LA PESQUERÍA DE LA MOJARRA RAYADA EUGERRES PLUMIERI (PISCES: GERREIDAE) EN LA CIÉNAGA GRANDE DE SANTA MARTA: ¿CAUSAS PESQUERAS, AMBIENTALES O BIOLÓGICAS?*
THE COLLAPSE OF THE STRIPED MOJARRA EUGERRES PLUMIERI (PISCES: GERREIDAE) FISHERY IN THE CIÉNAGA GRANDE DE SANTA MARTA: FISHING, ENVIRONMENTAL OR BIOLOGICAL CAUSES?
Efraín Viloria Maestre1, Arturo Acero P.2 y Jacobo Blanco3
1 Instituto de Investigaciones Marinas y Costeras -Invemar-, Cerro Punta Betín, Santa Marta, Colombia. efrain.viloria@invemar.org.co.
2 Universidad Nacional de Colombia sede Caribe, Centro de Estudios en Ciencias del Mar -Cecimar-. Invemar, Cerro Punta Betín, Santa Marta, Colombia. aacero@invemar.org.co.
3 Universidad del Magdalena, Programa de Ingeniería Pesquera, Santa Marta, Colombia. jablarce@gmail.com.
RESUMEN
Se analiza el colapso de la pesquería de la mojarra rayada (Eugerres plumieri), recurso tradicional de la Ciénaga Grande de Santa Marta, en 1994-1996 y 2000-2007. El enfoque BACI (Before and After Control Impact) relaciona aspectos pesqueros, biológicos y ecológicos con la abundancia de mojarra rayada. Los resultados indican que la pesca no fue determinante para disminuir la abundancia de E. plumieri, sino una serie de eventos distintos relacionados en cascada. Aparentemente, en corto tiempo la mojarra se vio privada de su alimento preferido Mytilopsis sallei, pequeño bivalvo sésil antes muy abundante sobre bancos de ostra (Crassostrea rhizophorae). Estos fueron sepultados por los sedimentos aportados por tributarios de la Sierra Nevada de Santa Marta, inicialmente, y luego también por las obras de reconexión del sistema lagunar con río Magdalena desde 1996. Se presentan modelos que explican el cambio en la abundancia de mojarra ante la variación de captura y esfuerzo pesquero, así como la disminución de la capacidad de carga para mojarras y presas, por el impacto de los sedimentos provenientes de ecosistemas vecinos. Se enfatiza la necesidad y utilidad de un enfoque ecosistémico al considerar el manejo de recursos pesqueros.
PALABRAS CLAVE: Colapso de pesquerías, mojarra rayada, enfoque ecosistémico, BACI, regresión segmentada.
ABSTRACT
The collapse of the striped mojarra (Eugerres plumieri) fishery, a traditional resource in Ciénaga Grande de Santa Marta, from 1994-1996 to 2000-2007, is analyzed. The BACI (Before and After Control Impact) approach used relates fishery, biological and ecological aspects with striped mojarra abundance. The outcome point out that fishing was not determinant of E. plumieri abundance diminution, but a series of different related and cascading events. Seemingly, mojarras were deprived in short time of their preferred food (Mytilopsis sallei), a small sessile bivalve very abundant at one time on oyster banks (Crassostrea rhizophorae). These were buried by sediments delivered initially by tributaries from Sierra Nevada de Santa Marta, and then, since 1996, by works connecting lagoon to Magdalena River. Models are proposed, explaining abundance variation confronted with catch and effort, as well as reduction of carrying capacity for mojarras and their prey, due to impact of sediments proceeding from neighboring ecosystems. The need and usefulness of an ecosystem approach when considering fish resource management is emphasized.
KEY WORDS: Fishery collapse, striped mojarra, ecosystem approach, BACI, piecewise regression.
INTRODUCCIÓN
La Ciénaga Grande de Santa Marta (CGSM) es un área de interés social y económico por su producción pesquera, que suple parte del mercado del norte de Colombia. De allí se extraían ostras y caracoles, ecológica y socioeconómicamente importantes (Carranza et al., 2009) que eran cerca de la mitad de la biomasa desembarcada (Santos-Martínez y Viloria, 1998). Los bancos de ostras servían como hábitat para una variada fauna (crustáceos, moluscos y poliquetos) que utilizaba el sustrato duro y que era alimento de varias especies ícticas. Otros recursos han sido principalmente peces y crustáceos, como la mojarra rayada (Eugerres plumieri), que, junto con Mugil incilis y Cathorops mapale, representaba entre los años 1994- 1996 alrededor del 80% de la captura total de pescado en la CGSM (Santos-Martínez y Viloria, 1998). Sin embargo, de 2000 a 2007 la ostra prácticamente desapareció como recurso y la extracción de las tres especies de pescado descendió a 33% del total, con mayor disminución de E. plumieri, que pasó de un 23% del total en el período 1994-1996 a menos de 1% en 2007, de su pesquería, aunque no desapareció la especie. Siguió una reducción del esfuerzo pesquero en los artes que capturaban mojarra rayada (atarrayas y boliches) por desmotivación de pescadores ante la baja producción.
Lo anterior originó preocupación local e incertidumbre en cuanto a las probables causas de la reducción en la abundancia de esta especie, lo que llevó a este trabajo. Existen estudios adicionales relativos a esta especie, dinámica poblacional (Rueda y Santos-Martínez, 1999; Olaya-Nieto y Appeldoorn, 2004), distribución y abundancia espacio temporal (Arenas-Granados y Acero, 1993, Sánchez y Rueda, 1999; Rueda, 2001; Rueda y Defeo, 2001) y otros relacionados (Santos-Martínez y Acero, 1991; Arenas-Granados y Acero, 1992; Rueda y Defeo, 2003). Pero no se habían estudiado las causas probables de este descenso, ni el o los factores limitantes para una futura recuperación de la mojarra rayada.
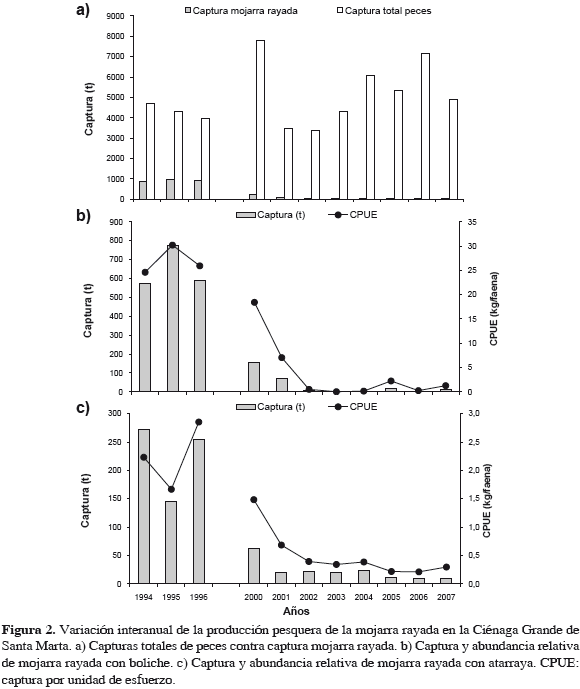
La mojarra rayada (Figura 1) es un gerreido, grupo de peces eurihalinos de gran importancia económica y ecológica que cumplen una función definida en la red trófica de las lagunas costeras (Aguirre-León et al., 1982; Cervigón et al., 1992). Randall (1967) define las mojarras como consumidores bentónicos, con una boca muy protráctil con la que se alimentan de invertebrados de la infauna. Esta especie crece hasta 40 cm y 600 g, común hasta 25 cm (Cervigón et al., 1992). Para la CGSM, se ha estimado una longitud asintótica (Lµ) de 41.9 cm y una talla media de madurez de 20 cm (Rueda y Santos-Martínez, 1999). Se distribuye en el Atlántico occidental: de Carolina del Sur (EE. UU.), el golfo de México hasta Brasil; ausente en Bahamas y otras pequeñas islas en las Antillas (Cervigón et al., 1992). El declive de la captura total anual de la especie en la CGSM y la variación de su abundancia relativa (CPUE) para boliches y atarrayas, los principales artes de pesca de este recurso (Figuras 1 y 2), justifican la denominación "colapso pesquero" utilizada en este trabajo, con el sentido que tiene la palabra de desplome, caída, ruina y que ha sido aplicado en economía y pesquerías (Pauly y Tsukayama, 1987; Boreman et al., 1997; Hutchings, 2000, 2004; Overholtz, 2002; Burgos y Defeo, 2004; Imarpe, 2004, 2005; Stone et al., 2004; Mullon et al., 2005; García et al., 2007; Klanjscek y Legovic, 2007; Longhurst, 2007) y hasta a los ecosistemas (Jackson et al., 2001).

ÁREA DE ESTUDIO
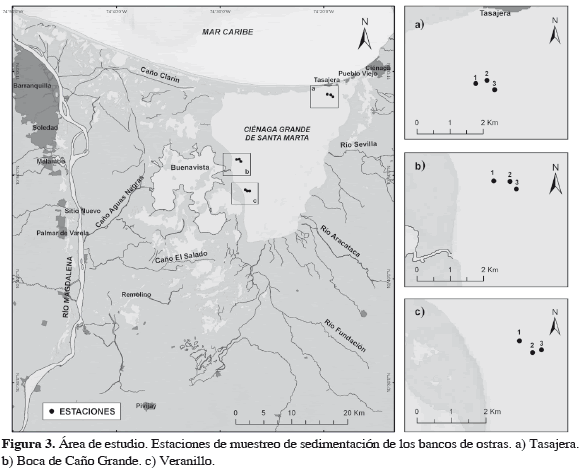
La CGSM se localiza en el litoral del departamento del Magdalena, al norte del país (Figura 3). Descrita en la literatura (Kaufmann y Hevert, 1973; Wiedemann, 1973; Botero y Salzwedel, 1999, Rivera-Monroy et al., 2001), recientemente se conoce la vinculación de su hidrología con la variabilidad climática global (El Niño Oscilación Sur o ENOS) y su importancia para las pesquerías (Blanco et al., 2006, 2007). En las últimas décadas ha sufrido serias perturbaciones ambientales, de magnitud relacionada con cambios del medio físico, aún poco conocidos. Clima, hidrología y obras civiles han contribuido a estas perturbaciones. También es afectada por la erosión y la contaminación en la Sierra Nevada de Santa Marta (SNSM).
La CGSM está ligada a los medios con los que está conectada, como los ríos de la SNSM y el río Magdalena. Además, la actividad antropogénica afecta el aporte de agua dulce y marina, controlando la salinidad y el ingreso de sedimentos que influyen en la columna de agua de la ciénaga (Botero y Mancera-Pineda, 1996). Por aumentos drásticos de la salinidad en 1993-1995, se reconectó al complejo lagunar de Pajarales (CP) con el Magdalena mediante tres caños (Botero y Salzwedel, 1999, Rivera-Monroy et al., 2006). La CGSM recibió estos cambios a través de Pajarales en 1996 y 1998, manifestados en la composición de la comunidad (Blanco et al., 2006). La pesquería de la CGSM, una de las más tradicionales del país, tenía entre los principales recursos en 1994 y 1995 la ostra (50% de la biomasa anual extraída) y la mojarra rayada (Santos-Martínez y Viloria, 1998). Los bancos de ostra (Crassostrea rhizophorae) (Figura 1), hoy sepultados, eran extensos a principios de los años 90 del siglo pasado (Hernández y Márquez, 1987; Mancera y Mendo, 1996). Sobre las conchas de estos bancos, crecía la presa principal de las mojarras, el bivalvo dreisénido Mytilopsis sallei (Arenas-Granados y Acero, 1992), con una densidad estimada de 5000 individuos/m2 (Puyana, 1995).
MATERIALES Y MÉTODOS
Se compararon gráficamente las variables pesqueras (esfuerzo con boliche -arte cuyo objetivo era la mojarra rayada-); físicas (sedimentos -como sólidos suspendidos totales (SST)- y salinidad, como probables tensores sobre la mojarra o su alimento, Mytilopsis sallei (Arenas-Granados, 1992)); biológicas (alimento como M. sallei, cuya abundancia está asociada a la capacidad de carga del sustrato duro -los bancos de ostra (Puyana, 1995)- en escenarios antes (1994-1996) y después (2000-2007)). Teniendo en cuenta lo anterior, se evaluaron los efectos sobre la especie y su relación con el ambiente (Walters y Martell, 2004) buscando una valoración objetiva, a través de la aplicación del enfoque denominado Before and After Control Impact (BACI), de las condiciones en escenarios antes (1994-1996) y después (2000-2007) (Bernstein y Zalinski, 1983; Underwood, 1991; Smith, 2002); aquí el "antes" puede considerarse también como el control o el punto de referencia.
La mojarra rayada y su pesquería en la CGSM
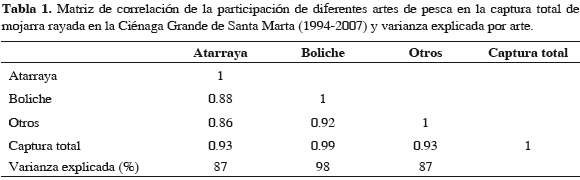
Siendo un recurso tradicional, implicaba dilucidar la responsabilidad de la pesca en la disminución de su abundancia. Los datos de capturas y esfuerzo de los principales artes de pesca, se calcularon a través del Sistema de Información Pesquera del Invemar (Sipein) (Narváez et al., 2005). Se obtuvieron estimaciones mensuales y luego anuales de las capturas por arte de pesca; de igual manera para el esfuerzo (faenas). Para determinar cuál arte de pesca ejercía mayor presión pesquera se realizó una regresión lineal entre captura total anual y capturas con las principales artes para pescar mojarra, boliche y atarraya (Narváez et al., 2008) y como criterio se tomó la varianza explicada. Se calculó el esfuerzo anual del arte de mayor incidencia de 1994 a 2007 y se hizo un análisis de correlación y de regresión lineal para estimar la influencia del esfuerzo sobre las capturas totales anuales (abundancia) de la mojarra en CGSM.
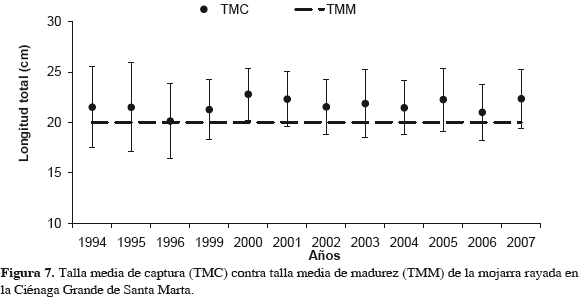
Para ver la variación de la mojarra, en cada escenario, con los promedios de cada variable, se calculó la variación (%) antes-después para detectar correspondencias en los cambios de las variables esfuerzo de boliche (ESFBOL), captura total de ostra (OSTRA), logaritmo natural de sólidos suspendidos totales acumulados (SST AC), salinidad (SALIN) y CPUE de la mojarra (CPUEAN), como indicadores de abundancia. Para evaluar el impacto de la pesca sobre este recurso en las tallas, se estimó la talla media de captura (TMC) anual, indicador de la pesquería en términos de sobrepesca por reclutamiento, comparada anualmente con talla media de madurez sexual (TMM), como punto de referencia límite biológico (PRL).
La mojarra rayada y su alimentación
Se analizó la composición de contenidos estomacales de mojarras, para determinar la importancia de sus componentes en 132 ejemplares (Arenas-Granados y Acero, 1992). Por la asociación M. sallei-bancos de ostra se analizan los cambios ocurridos de antes a después. Los valores fueron comparados con los de principio de los años 90 (Arenas-Granados y Acero, 1992). Como indicador de abundancia de dreisénidos en "antes" se utilizó el área de bancos de ostra, disponibles como sustrato y la biomasa anual de ostra (t) extraída de dichos bancos, registrada en Sipein. Se estimó la biomasa total de dreisénidos en "antes" con base en datos de densidad y biomasa (Puyana, 1995), comparada con su ausencia después.
Aspectos ambientales: la sedimentación en la CGSM
Los dreisénidos, como las ostras, son sésiles, sensibles a factores como salinidad y sedimentación, asociados a que la CGSM estuvo sometida a serios cambios hidráulicos e hidrológicos en décadas pasadas. Se estimó la influencia de dichos factores en la abundancia del alimento (dreisénidos) y de mojarra rayada. Se partió del esquema de Wiedemann (1973), que señala como origen de sedimentos en la CGSM al mar adyacente, los ríos de la SNSM y las comunicaciones con el río Magdalena. No se contó con una secuencia temporal de la sedimentación en la CGSM, aunque sí de los sólidos suspendidos totales (SST).
Se utilizó información de muestreos en estaciones prefijadas, como Boca del Caño Grande, La Rinconada, Rincón del Jagüey, centro CGSM y frente a boca de los ríos tributarios Aracataca, Fundación y Sevilla, con frecuencia trimestral, en superficie, media agua y fondo, registrados en la base de datos físicoquímicos del monitoreo ambiental de Invemar en la CGSM; de igual manera se realizó la variación contemporánea de los SST y el caudal de los ríos de la SNSM entre 1993 y 2007. Se analizó gráficamente la variabilidad anual de los SST, para definir tendencias. Como los datos de fondo corresponden a la fracción más densa y prontamente sedimentable, ellos se escogieron para los análisis, como indicativos de la tendencia de la sedimentación ocurrida de 1993 a 2007. Se supone que SST son sólidos suspendidos antes de depositarse como parte de los sedimentos de un cuerpo de agua. Es claro que los conceptos de sólidos suspendidos sedimentables (SSS) y SST son diferentes y no son intercambiables (Gray et al., 2000). Los SST no incluyen la fracción suspendida más densa de las arenas y las gravas, que decantan rápidamente por lo que sus datos estarían subvalorando la concentración real de los sólidos sedimentables presentes (APHA et al., 2005). Como el depósito de los sedimentos es un proceso acumulativo en el tiempo, los valores de SST corresponden a concentraciones instantáneas (mg L-1) en diferentes tiempos. Sin embargo, para tener una idea aproximada de la tendencia de mayor o menor influencia de los sedimentos en la laguna, se han utilizado los promedios anuales de SST de forma aditiva, para representar (Proxy) una acumulación progresiva en el tiempo. Los promedios de SST acumulados de 1993 a 2007, transformados a Ln para suavizarlos (y = Ln x), para describir así una tendencia clara y, tratándose de valores adimensionales, no entra en conflicto con unidades de datos originales.
Para dimensionar el impacto de la sedimentación sucedida hasta el presente sobre los bancos de ostra, se realizaron muestreos en abril de 2008 con sus réplicas sobre tres de los principales bancos de la CGSM (Figura 3) con un tubo corazonador plástico (1.2 m de longitud, 4 cm de diámetro). La columna de sedimento se midió en campo en corazonador, se descartó el exceso de agua y se almacenó en bolsas plásticas rotuladas para muestra y réplicas en cada banco. La columna de agua se midió sobre cada banco de ostras. Luego, se hizo el análisis granulométrico por tamizado (Holme y McIntyre, 1971).
Análisis de las relaciones entre las variables
La abundancia de mojarra como CPUE estimada anualmente se seleccionó como variable dependiente. Como predictores se tomaron ESFBOL, esfuerzo pesquero del arte selectivo de pesca boliche (faenas año-1), especie objetivo mojarra rayada, salinidad media anual en el sistema, SALIN; SST AC promedio anual acumulado 1994-2007; y OSTRA (t año-1). Inicialmente se hizo un análisis gráfico de dispersión de datos de las variables en tiempo 1994-2007. Ante una evidente discontinuidad en las variables del escenario "antes" al "después", se utilizó una metodología que operara con estos cambios: regresión por segmentos o segmentada (McGee y Carleton, 1971; Toms y Lesperance, 2003; Harper et al., 2005); aplicada para estimar cambios relacionados con pesquerías por autores como Maceina (2007), Brenden y Bence (2008), Maceina y Hunter (2008) y Holley et al. (2009). En este caso, la abundancia anual de mojarra (CPUEAN) es la variable dependiente, que varía distintamente con variables independientes, como ESFBOL, SST AC, OSTRA y SALIN. Los datos no siguen ninguna combinación lineal definida o con respecto a captura; por tanto, es difícil modelar una relación tan dinámica (antes-después) usando métodos lineales convencionales, como regresión múltiple multivariada. Se plantearon y resolvieron ecuaciones empíricas de dos segmentos ("antes", "después") usando método no lineal Quasi-Newton (Prasad et al., 2006). El programa iterativo Statistica (StatSoft, 2003) calcula dos ecuaciones por separado:
y = (b01 + b11 *x1 +...+ bm1 * xm) * (y ≤ bn) + (b02 + b12 * x1 +...+ bm2 * xm) * (y > bn)
Para optimización multivariada se usa el método de Quasi-Newton que, siendo no lineal, sirve para minimizar la función de ajuste (loss) mediante convergencia iterativa de una ecuación predefinida. Para la aceptación de modelos empíricos resultantes se tuvieron como criterios máxima verosimilitud (Hudson, 1966), bondad del ajuste con mayor correlación, mayor varianza explicada y menores residuos para cada ecuación.
Inicialmente se estimaron modelos de regresión segmentada con CPUEAN como variable dependiente y en el ámbito de cada variable independiente. En este análisis se permitió que el programa asumiera automáticamente un punto de quiebre, buscando un mejor ajuste. Con efecto combinado de variables se estimó un modelo múltiple de regresión segmentada explicativo dentro del ámbito de variable dependiente (CPUEAN). Además, se estimaron modelos de regresión segmentada con otras variables, como con la abundancia de la ostra (dependiente) y teniendo como independientes SST y SALIN.
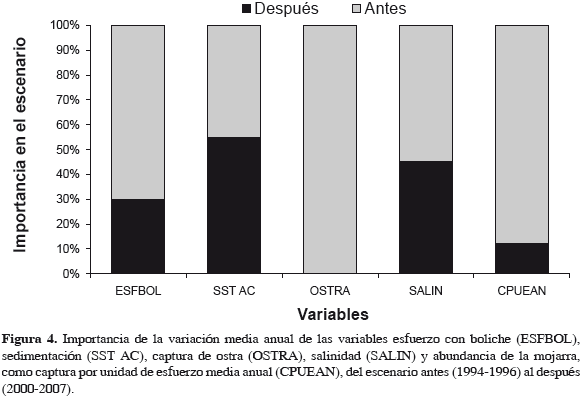
RESULTADOS
De un escenario a otro, las variables de mayor cambio fueron OSTRA y CPUEAN; ESFBOL bajó 57.0% y SALIN 18.2%, pero sedimentos (SST AC) mostraron incremento de 22.0% (Figura 4).
La mojarra rayada y su pesquería en la CGSM
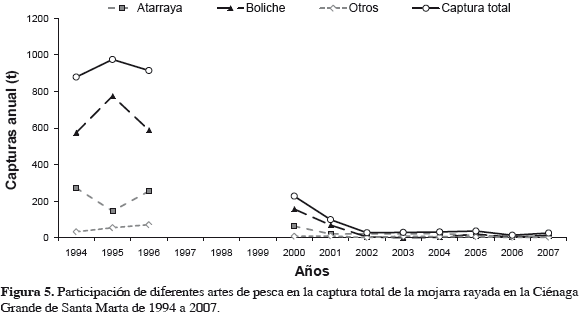
En cuanto a capturas por arte de pesca (Figuras 5 y 6, Tabla 1), destaca el boliche, cuya especie objetivo fue la mojarra. La CPUE con este arte tuvo su máximo en 1995; mientras que con atarraya la CPUE máxima sucedió en 1996 (Figura 2). La Figura 6 muestra que, mientras el esfuerzo que disminuyó en "después", oscila alrededor de 10000 faenas anuales, las capturas disminuyeron sin recuperarse. Aplica aquí CPUE de la mojarra con boliche como indicativo de la abundancia del recurso.
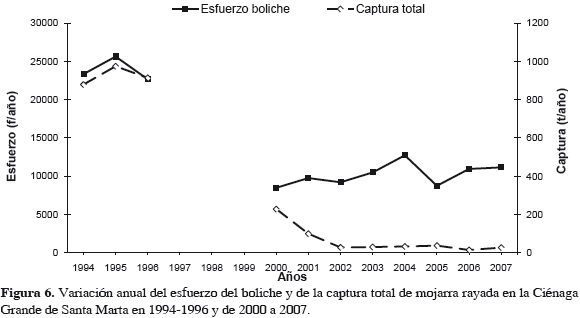
Aparentemente las mojarras se concentraron para alimentarse donde quedaban bancos de ostra parcialmente descubiertos en 1995-1996, siendo más fácilmente capturadas por pescadores con boliches y atarrayas, lo que explicaría los altos valores de CPUE esos años (Figura 2), pero no después. En lo relativo a tallas de captura, la Figura 7 muestra que no hubo mayor efecto reductor en estas tallas, porque aunque en algunos años las TMC se acercaron al punto de referencia límite biológico, TMM, siempre lo sobrepasaron.
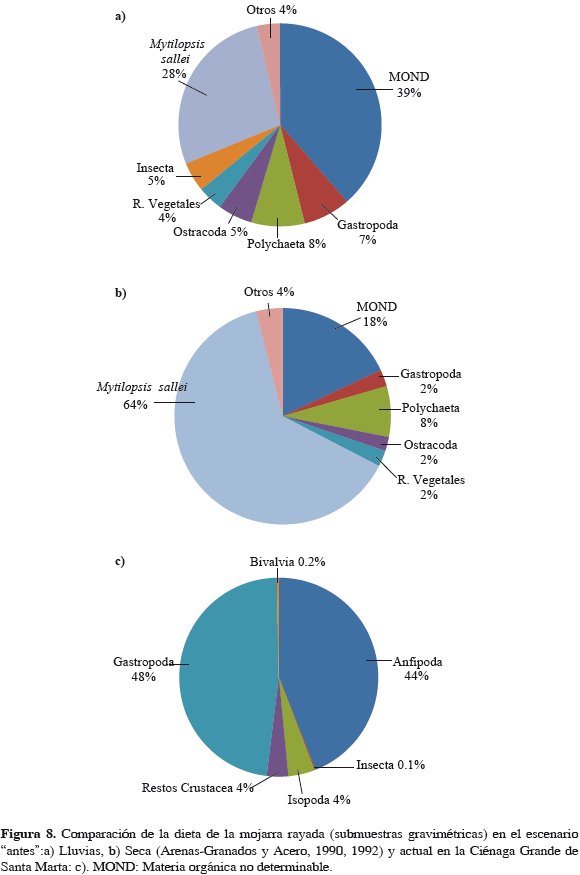
La mojarra rayada y su alimentación
La Figura 8a y b muestra la importante participación de M. sallei en la dieta en "antes", mientras que la Figura 8c evidencia su ausencia en el escenario actual. Su disponibilidad en los bancos de ostra en el escenario antes (Tabla 2), estimada de datos publicados de área de bancos (Mancera y Mendo, 1996) y de biomasa por metro cuadrado (Puyana, 1995), indican que esta varió entre 490 y 1140 t de dreisénidos en peso húmedo, importante cantidad para las mojarras, que rápidamente desapareció. La disponibilidad de dreisénidos cayó prácticamente a cero al quedar sepultados los bancos a comienzos del "después".

Aspectos ambientales: la sedimentación en la CGSM
Variación contemporánea entre SST y caudal de los ríos de la SNSM
El principal aporte de sedimentos en el escenario "antes" provino de SNSM acarreado por los tributarios. Los caudales de ríos de SNSM aumentaron de 1993-1996, pero concentraciones de SST disminuyeron, como también en 1999- 2000; mientras que en 1997-1998 al disminuir caudales, los SST de estos ríos se incrementaron, alcanzando sus máximos relativos, excepto los del Aracataca que bajaron (Figura 9). En 1993-1995 los ríos de SNSM mostraron incremento importante en contenido de SST y se infiere de sedimentos totales, pero los niveles de 1997-1998 seguramente fueron suficientes para causar un impacto decisivo sobre bancos de ostra y comunidad asociada. La aparente relación inversa caudal contra SST se explica porque a mayor volumen (caudal), menor es la densidad de partículas por unidad de volumen (mg L-1). También debido a resuspensión de partículas sedimentables por acción del viento, que es mayor en la época seca.
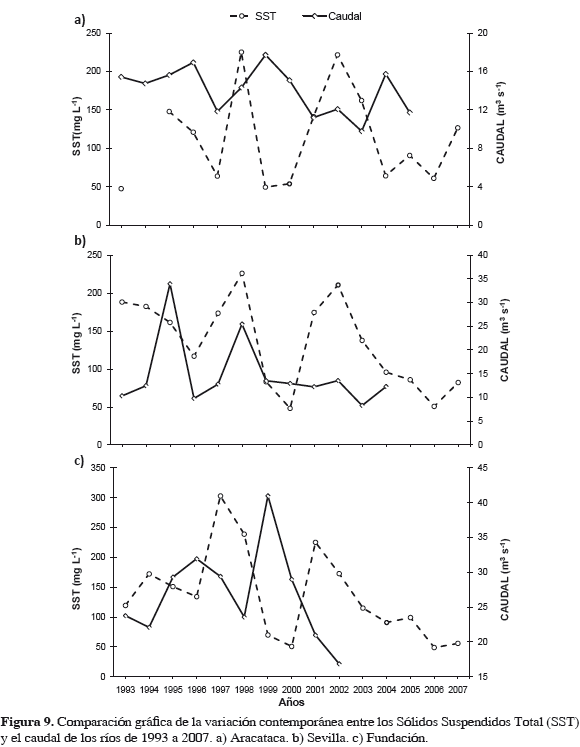
Proceso de sedimentación progresivo en la CGSM
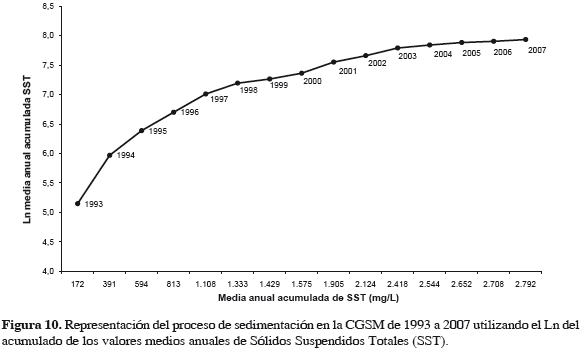
El resultado de la transformación logarítmica de valores acumulados de SST en el tiempo, indica una función de tipo exponencial creciente del proceso de acumulación de sólidos depositables en CGSM (Figura 10). El esquema muestra cómo el incremento de los sedimentos es mayor de 1993 a 1999 que de 2000 a 2007; aumentando siempre de año en año, lo cual es preocupante por su carácter acumulativo. La relativa disminución en 1999-2000 se explica por el mayor volumen de agua que ingresó al sistema en esos años, disminuyendo la densidad de SST en muestras (mg L-1), aunque no necesariamente la cantidad de sedimentos que penetró al sistema.
Sedimentos actuales sobre los principales bancos de ostra en la CGSM
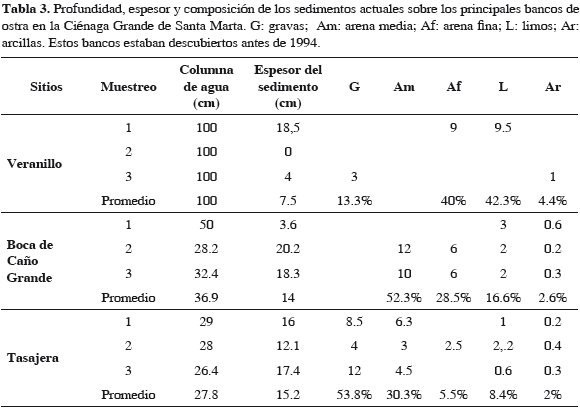
Actualmente la altura de los sedimentos sobre los bancos sepultados es inversamente proporcional a la altura de la columna de agua en cada sitio (Tabla 3). Las fracciones de sedimentos más gruesos, gravas y arenas medias, fueron mayores en Tasajera y frente a Boca de Caño Grande; mientras que en Veranillo predominan los sedimentos más finos, como arenas finas, limos y arcillas.
Análisis de las relaciones entre las variables
Relación entre la abundancia de la ostra, la salinidad y el proceso de sedimentación en CGSM
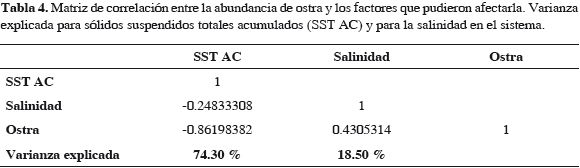
El 74.3% de la variación en la abundancia de la ostra es explicada inversamente por el cambio en la sedimentación (SST AC). Sin embargo, el impacto biológico se dio tan pronto los bancos fueron cubiertos. El cambio en la salinidad apenas explica directamente el 18.5% de dicha variación (Tabla 4) (Figura 11).
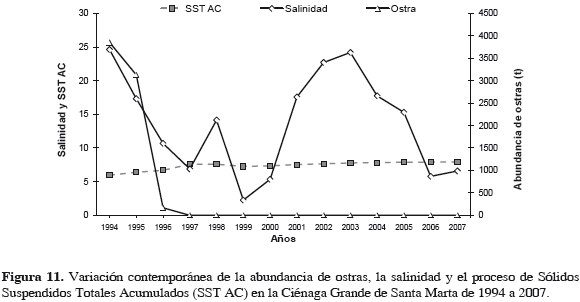
Modelos explicativos
La variación anual de la abundancia de la mojarra rayada en función de cada predictor y en el ámbito de cada uno, se representa en Figura 12. Los coeficientes calculados para cada modelo segmentado resultante se relacionan en Tabla 5. El punto de quiebre de la abundancia de la mojarra estimado fue 10.07 kg f-1 en todos casos. Las varianzas explicadas por modelos fueron superiores a 94%, indicando alto grado de correspondencia entre la variación de valores observados y estimados por modelos en los escenarios, antes y después. Sin embargo, la expresión multivariada de regresión segmentada, mostró un mejor ajuste al actuar conjuntamente variables predictivas sobre la abundancia de la mojarra rayada en ambos escenarios (Figura 13).
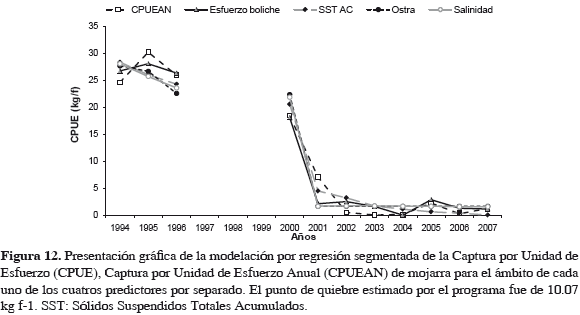
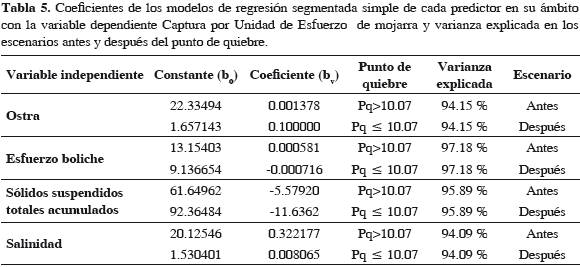
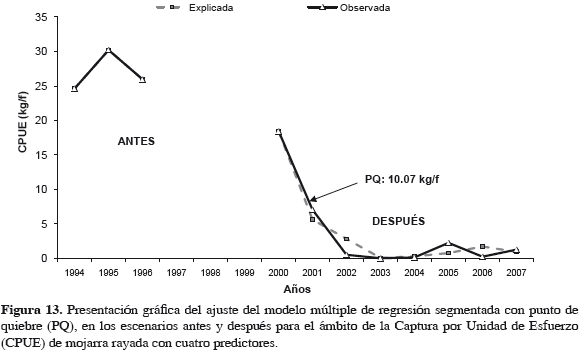
El punto de quiebre calculado por programa fue una abundancia de 10.07 kg f-1, como en modelos anteriores. La correlación fue alta (0.99) (Tabla 6) y la varianza explicada por modelo fue 99.17%, indicando que el modelo corresponde de cerca con variación observada en la abundancia de la mojarra, en los dos escenarios. Cuando la abundancia de la mojarra estuvo debajo del punto de quiebre (10.07 kg f-1) para 2001, en escenario después, ya no se recuperó. Lo que se muestra en los residuos resultantes (Tabla 7). Al ser la relación intrínsecamente no-lineal, los estimativos de los parámetros (coeficientes de regresión) y los errores típicos de esos estimativos pueden parecer anómalos, menores o inadecuados, pero hay que tener en cuenta que estos coeficientes corresponden a segundas derivadas de la función (Statsoft, 2003). El esfuerzo de boliche (b12 y b11) del escenario antes al después bajó a la mitad, probablemente desmotivado por bajas de CPUE, pero en escenario después (2000-2007) se nota una ligera disminución, indicativa de cambio lento (Tabla 6). Esto sucede en una magnitud tan baja, que ya es poco probable su efecto sobre la abundancia de la mojarra.
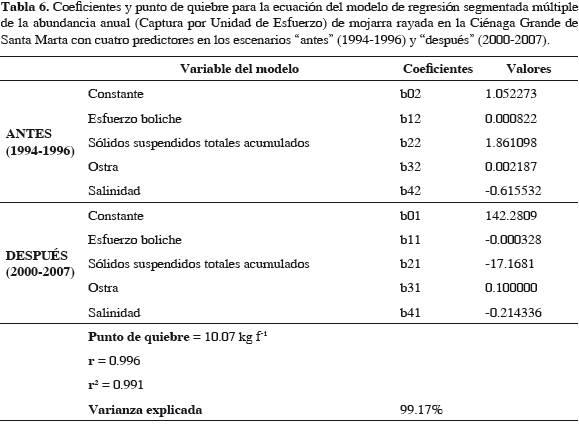
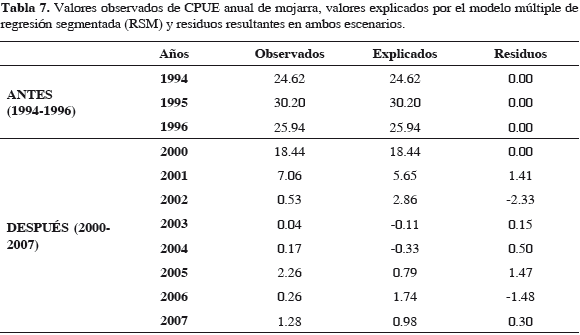
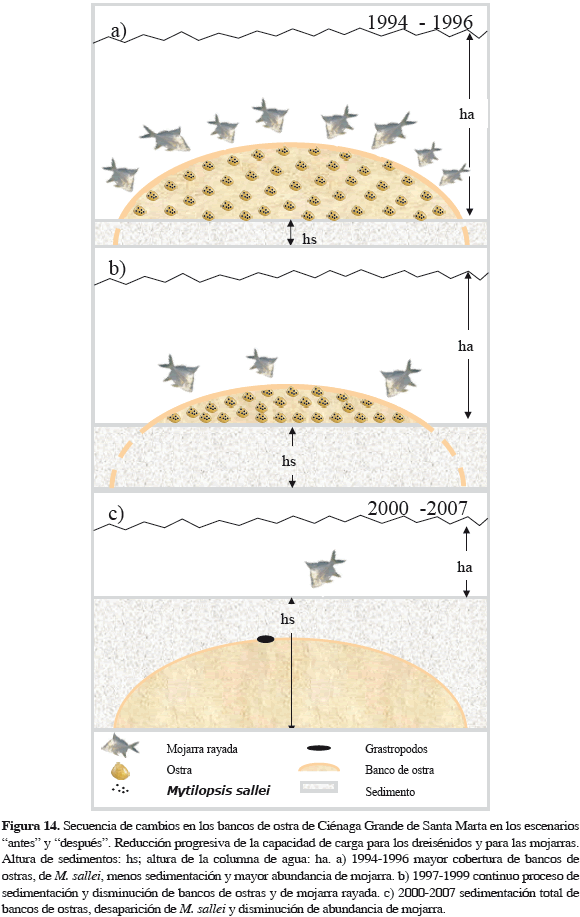
Tampoco fue rápido el cambio de pendiente con la salinidad, b42 a b41 de un escenario al siguiente. Sin embargo, este cambio no llega ser de un orden de magnitud, por lo que no se considera significativo. Con SST, el cambio en pendiente del escenario "antes" al "después", muestra una dimensión de cambio significativa, de un orden de magnitud. Es decir que se aceleró diez veces más que en escenario antes, sugiriendo la celeridad con que ocurrió el cambio en las variables (Tabla 6). Los coeficientes de regresión con ostra (b32, b31) muestran (Tabla 6) que de "antes" a "después" hubo un cambio acelerado en pendiente, unas 100 veces más rápido. Esto sugiere que el cambio sucedió en corto tiempo. El signo de coeficientes se mantuvo positivo, implicando relación positiva (directa) en ambos escenarios; así, a menos abundancia de ostras, menos de mojarra, ya que lo importante para el pez era el dreisénido de su dieta, fijado a las ostras. De 1994 a 1997 la abundancia de ostra cayó de unas 4000 t anuales a cero (Figura 11), sin recuperarse y correspondientemente la abundancia de mojarra disminuyó drásticamente hasta 2007, como se ilustra en la Figura 14.
DISCUSIÓN
Hay cierta analogía entre estudio de colapsos pesqueros y el análisis forense; el investigador se enfrenta a hechos consumados y en autopsia (Smith y Link, 2005) debe caracterizar el agente fatal, para conocer por qué se produce y para evitar que vuelva a suceder. Por tratarse de un recurso pesquero era previsible enfocar el colapso de la pesquería de la mojarra desde el punto de vista pesquero clásico de esfuerzo y captura, la presión pesquera, entre otros. Existen serias críticas al uso extendido de correlaciones simples y de asumir causalidad sin discutir explicaciones alternativas, ni mecanismos para su interpretación. Se desecha la posibilidad de que una tercera variable pueda explicar la relación observada (Longhurst, 2007). Además, teniendo en cuenta los cambios recientes en la CGSM, cabía pensar un impacto ambiental sobre el recurso y por ello el enfoque debía ser fundamentalmente ecosistémico, incluyendo elementos cuyo efecto era probable (Jackson et al., 2001). El enfoque BACI permitió determinar causas probables del colapso de la pesquería de E. plumieri, entre 1994 y 2007, relacionando aspectos pesqueros, biológicos y ecológicos con la abundancia de la especie en estudio, enfatizando la necesidad de un enfoque ecosistémico.
La discontinuidad en valores de variables de uno a otro escenario pudo absolverse mediante una metodología que opera en tales condiciones, como regresión segmentada (McGee y Carleton, 1971; Toms y Lesperance, 2003; Harper et al., 2005), usada para estimar cambios en pesquerías (Maceina, 2007; Brenden y Bence, 2008; Maceina y Hunter, 2008; Holley et al., 2009). La situación anterior es intrincada, más si se tiene en cuenta que entre los sistemas más complejos conocidos están los ecosistemas (Maurer, 1998). Las interacciones entre sus componentes son difusas en tiempo y espacio haciendo que no sea posible delinear límites de mayoría de ecosistemas. Esto hace difícil establecer relaciones causa-efecto entre los componentes observables de ecosistema; como la CGSM, en donde la estadística convencional necesita suposiciones sobre la naturaleza de datos que no se cumplen fácilmente en investigación ecológica. Aunque las estadísticas sean robustas ante desviaciones de supuestos, el mayor problema es intentar inferir causalidad a partir de resultados de pruebas formales. La teoría presume que los ecosistemas pueden modelarse con ecuaciones diferenciales que describen desviaciones del equilibrio de densidades poblacionales de una especie (Pielou, 1977).
Los intentos de modelación han sido simplistas, frecuentemente ajustados estrictamente a mecánica estadística desarrollada para situaciones de equilibrio en termodinámica (Maurer, 1998). El pensamiento linear en ecosistemas, con suposición de que están en equilibrio o que son estables, se reemplaza por la visión de que los ecosistemas están en cambio constante y que esos cambios dependen de condiciones experimentadas por el ecosistema antes de las mediciones. Además, los procesos que afectan la colecta de datos en un ecosistema, operan con frecuencia a escalas tiempoespacio distintas y rara vez sus relaciones son lineales. De otra parte, inferencias sobre causalidad presumen finalmente conocimiento implícito de un modelo del proceso, pero con frecuencia tales modelos son difíciles de formular y de analizar (Maurer, 1998).
Un enfoque precautorio del manejo demanda mejor conocimiento de la dinámica de colapsos pasados y detección de señales de alerta temprana (Mullon et al., 2005). La teoría convencional pesquera (Beverton y Holt, 1957; Ricker, 1975) se basa en el principio del equilibrio, que garantiza que, si el esfuerzo es constante y no muy alto, se logre la estabilización del stock y la captura y nunca habría colapso; pero sí los hay (Mullon et al., 2005). En concordancia, la forma de ver un ambiente lo asumía estable e infinitamente resiliente, en equilibrio general. La nueva perspectiva reconoce que la resiliencia puede ser o ha sido degradada y que no se debe dar por sentada la capacidad propia de reparación de ecosistemas (Folke et al., 2005). Aquí se considera la resiliencia como la capacidad del sistema para absorber la perturbación y reorganizarse al sufrir el cambio, de modo que retenga su misma función, identidad y retroalimentación (Folke et al., 2005).
Dada la importancia de E. plumieri se suponía una causa pesquera para el colapso (Santos-Martínez y Viloria, 1998; Rueda y Santos-Martínez, 1999; Sánchez y Rueda, 1999; Rueda y Defeo, 2003; Olaya-Nieto y Appeldoorn, 2004; Rueda, 2007). Los resultados indican que la pesca no fue determinante en la disminución de la abundancia, sino eventos distintos relacionados en cascada. Independientemente de los cambios en el esfuerzo, las capturas continuaron disminuyendo, sin embargo, la relación no es lineal y se comportó diferentemente entre "antes" y "después". El efecto sobre tallas de captura tampoco fue definitivo en la declinación de los rendimientos de la especie. Aunque en algunos años se acercó al punto de referencia limite (TMM), se supone que el riesgo de sobreexplotación fue moderado, ya que TMC superó a TMM en todos los años, en gran parte por selectividad del arte de mayor eficiencia de captura, el boliche (Rueda y Santos-Martínez, 1997; Rueda et al., 1997; Narváez et al., 2008).
Del escenario antes (1994 -1996) al después (1999-2007) la CGSM sufrió considerables cambios de salinidad y de aporte de sedimentos (Blanco et al., 2006). Siendo un pez eurihalino, no era previsible que lo afectara mucho la salinidad del sistema, que no fue significativa para explicar el cambio en abundancia de la mojarra. Tampoco era previsible un efecto directo de la sedimentación sobre las mojarras, pero sí sobre su dieta preferencial, los dreisénidos asociados a los bancos en la CGSM. Los organismos del bentos estuarino están sometidos a eventos perturbadores causados por procesos hidrodinámicos que revuelven y mueven los sedimentos donde o hacia donde los animales viven. Sin embargo, son poco conocidos los mecanismos de la perturbación que afectan a las poblaciones de la infauna y la epifauna (Hinchey et al., 2006).
Teniendo en cuenta la importancia del alimento preferencial de las mojarras en el antes (M. sallei) y su asociación con los bancos de ostra, la dieta natural de la mojarra es un aspecto influyente en su abundancia. El análisis mostró la ausencia del dreisénido (M. sallei) en el escenario después, produciendo variaciones de comportamiento, como lo que actualmente ocurre con la especie. Nikolsky (1963) menciona que ocurren variaciones dentro de una especie de un año a otro, con relación a sus condiciones de alimentación. Comparando con otros hábitats afines, se encontró que en el área deltaica estuarina del río Sinú, aledaña a la bahía de Cispatá, al igual que en el antes de la CGSM, la especie también consumía mayoritariamente M. sallei, registrándose este ítem alimentario en un 50.5% (Parra, 1996).
La sedimentación causa reducción en la columna de agua y por tanto habrá menos volumen para seres acuáticos, así como menor capacidad de carga del ecosistema, alteraciones en la composición, geometría, calidad y cantidad del fondo y del bentos que lo habita, sean arrecifes o bancos de ostras (Jackson et al., 2001; Jha, 2007). Los procesos de sedimentación dependen de dinámica de masas de agua, cuando es baja se rigen por la ley de Stokes, siendo su deposición proporcional al tamaño de la partícula y a su densidad (Holme y McIntyre, 1971). Así, en sitios con mayor dinámica por corriente u oleaje, el depósito será menor y viceversa.
Las perturbaciones ambientales influyen en estructura y función de comunidades acuáticas. Debido a su poca profundidad, el acarreo de los tributarios e ingreso de energía asociada con mareas, oleaje, tormentas y corrientes, la perturbación física de los organismos bentónicos de una laguna como la CGSM, a causa de la sedimentación, es un rasgo común de ambientes costeros (Jackson et al., 2001). Estas perturbaciones en el medio físico pueden llegar a ser fuerzas que afectan a distintos niveles de organización, sea individual, poblacional o de la comunidad. Sin embargo, los efectos de una perturbación pueden diferir en el caso de organismos móviles y en el de sésiles, debido a su distinta capacidad de escape y de recolonización a partir de sobrevivientes (Breitburg, 1992). En este caso, las especies más afectadas por la perturbación fueron las ostras y los dreisénidos asociados, ambas especies sésiles que sufrieron un impacto de tipo físico al ser sepultadas por los sedimentos, mientras que el efecto sobre la mojarra fue de tipo trófico, al desaparecer sus presas.
Se sabe que Crassostrea virginica es capaz de sobrevivir hasta siete días a hipoxia causada por enterramiento bajo 0.2-0.5 cm de sedimentos, antes de que ocurra 100% de mortalidad (Hinchey et al., 2006). Un espesor de menos de un centímetro de sedimentos sobre un banco es suficiente para determinar muerte de las ostras y su epibiota sésil, y más si el grosor de la capa actual oscila entre 7.5-15.2 cm, como en la CGSM. Se confirma así que el proceso de sedimentación continuó después de que los bancos de ostra quedaron sepultados, impidiendo recuperación de los mismos. Esto reafirma el supuesto de que la mortalidad en los bancos locales ocurrió de una manera rápida como replica el modelo propuesto. Cada vez hay más evidencia que sugiere que con frecuencia los ecosistemas no responden al cambio gradual de manera uniforme (Folke et al., 2005), menos aún si el cambio es repentino, como parece que sucedió en CGSM.
Los sedimentos producidos por las obras de los caños también tuvieron un efecto, aunque posterior a 1996. Se removieron cerca de 3.1 x 106 m3 de tierra para hacer canales, pero esta tierra no salió del sistema, sino que la depositaron en orillas de caños. Lluvias e inundaciones hicieron que esos sedimentos fueran acarreados por los caños hacia la laguna. Cuando estos sedimentos llegaron a la CGSM en 1997, ya los bancos de ostra estaban sepultados, pero con este ingreso adicional de sedimentos, la situación se volvió definitivamente irreversible para ellos, para los dreisénidos y para sus depredadores, las mojarras.
Según Restrepo y Kjerfve (2000), la carga sedimentaria del río Magdalena a la altura de Calamar (Bolívar) es de aproximadamente 394 x 103 t día-1. Suponiendo que las trampas de sedimentos en la entrada del río a caños retuvieron 75% de esta carga, los sedimentos que ingresaron a caños constituyeron unos 98.5 x 103 t día-1, una cantidad formidable fluyendo en una dirección: el sistema lagunar. Si se usa el mismo modelo de Restrepo y Kjerve (2000) para calcular la carga de sedimento, pero con el caudal nominal de los caños Clarín, Aguas Negras y Renegado, 140 m3 s-1 , la carga para los tres caños llega a ser de 1420 t día-1, y aunque en sus lechos se depositara 75% de estos sedimentos, todavía habrían llegado al sistema lagunar cerca de 355 t día-1, un valor considerablemente alto, que sin duda agravó impacto de sedimentación en CGSM-CP.
La Figura 14 es la representación idealizada de lo que probablemente sucedió desde 1993. El colapso de la pesquería de la mojarra fue la etapa final de cascada de eventos de impacto ambiental, donde el factor desencadenante fue la carga de sedimentos producidos por erosión en la SNSM, que entró a la CGSM desde 1993 por tributarios y llegó a los bancos, sepultándolos con su comunidad asociada. Probablemente operó entonces el control trófico de abajo hacia arriba. Las mojarras sobrevivientes tuvieron que cambiar la dieta, consumiendo pequeños gastrópodos e infauna vágil asociada a sedimentos, que no son reemplazo para el alimento del que disponían, probablemente ni en calidad ni en cantidad. El cambio de condiciones debió ser rápido como sugiere la aceleración del cambio de las pendientes en el modelo segmentado. Es presumible que el impacto de los sedimentos sobre los bancos alteró la estructura de comunidad (diversidad y biomasa), las tasas de reproducción y de crecimiento (dreisénidos, ostras y otros), la eficiencia alimentaria (filtración, taponamiento), el comportamiento (competencia por espacio), la capacidad de carga de los depredadores (mojarras) y la competencia entre ellos.
No hubo evidencia de que la presencia de tilapia u otra especie eventualmente en aguas de bajas salinidades (< 10), tuviera influencia en la reducción de la abundancia de la mojarra, como también concluyeron Leal-Flórez et al. (2008) y Narváez et al. (2011). La mayor salinidad de las aguas en la CGSM siempre frenó la expansión de la tilapia a esta laguna. La caída abrupta de los niveles de captura de la mojarra, después de tener una relativa estabilidad en el escenario "antes", demuestra que esa estabilidad ocultaba un riesgo de colapso como el observado. Lo anterior contradice la suposición de que la estabilidad de capturas es garantía para la sostenibilidad de una pesquería (Mullon et al., 2005), sobre todo cuando hay una tensión ambiental. Cuando los eventos ocurren en cascada, generan cambios no lineales y con frecuencia sorprendentes en la dinámica de los ecosistemas, haciéndolos cambiar su régimen (Díaz y Cabido, 2001). Efectos combinados, a veces sinérgicos, de tales eventos hacen, más vulnerables a los ecosistemas frente a los cambios que podrían ser asimilados y en consecuencia pueden pasar de súbito de estados deseables a indeseables con respecto a su capacidad de prestar servicios ecosistémicos (Folke et al., 2005), como la pesca en la CGSM. Siguiendo la analogía de la práctica forense, los casos de colapsos pesqueros, como en autopsias, deben también estudiarse rigurosamente como formas de evaluar la asesoría en la toma de decisiones.
CONCLUSIONES
A pesar de la importancia de E. plumieri como recurso, la pesca no fue determinante en la disminución de la abundancia de la especie, sino una serie de eventos distintos en cascada: erosión en la SNSM, transporte de sedimentos por tributarios a la CGSM, enterramiento de los bancos y privación de dreisénidos para la mojarra. Dada la importancia de M. sallei como alimento para la mojarra y su fijación sobre los bancos, la suerte de la ostra la siguió M. sallei, lo que finalmente afectó la capacidad de carga de la mojarra. Probablemente competencia por alimentarse en últimos bancos de ostra descubiertos, favoreció agregación de mojarras en áreas donde fueron presa más fácil para los pescadores, contribuyendo a la reducción de su población.
Destacan la sedimentación y salinidad del agua en la CGSM, como factores naturales que afectaron los bancos. Sin embargo, mientras la salinidad puede variar, los sedimentos se acumulan y permanecen, dificultando la recuperación de las condiciones tróficas favorables para la ostra, para M. sallei y finalmente para la mojarra. No se encontró evidencia de que la tilapia u otra especie invasora afectaran de algún modo a la mojarra rayada, o que compitiera con ella por alimento o espacio.
En el análisis del colapso de otros recursos en sistemas lagunares similares a la CGSM, es necesario incluir, con visión ecosistémica, agentes bióticos y abióticos que razonablemente pueden perturbar su abundancia. El enfoque antes y después (BACI) y el uso de la regresión segmentada fueron útiles al representar lo sucedido con el recurso mojarra rayada en la CGSM, pero bien pueden aplicarse en otros casos.
AGRADECIMIENTOS
Los autores agradecen el apoyo a la realización de este trabajo al Invemar y su Programa Valoración y Aprovechamiento de Recursos Marinos y Costeros (VAR) a través de los proyectos "Evaluación de los principales recursos pesqueros de la Ciénaga Grande de Santa Marta" (1993-1996), y del proyecto "Monitoreo de las condiciones ambientales y los cambios estructurales y funcionales de las comunidades vegetales y de los recursos pesqueros durante la rehabilitación de la Ciénaga Grande de Santa Marta" (2000-2007). A la Universidad Nacional de Colombia y al Cecimar. A la Doctora G. Guzmán por sus aportes. A la Ing. Pesquera L. Vivas de Redcam, por su apoyo. A L. Ordóñez por su colaboración en el análisis de composición de contenidos estomacales de las mojarras. A los auxiliares de campo de la Ciénaga Grande de Santa Marta: H. Rodríguez, V. Carbonó, J. Suárez, Y. Palomino, A. Beleño y C. Carbonó; a la digitadora S. Chávez; al Tecnólogo en Sistemas J. Romero, por su apoyo con Sipein; a la Ing. Pesquera D. Mármol, por su colaboración, y a los pescadores de la Ciénaga Grande de Santa Marta.
BIBLIOGRAFÍA
1 Aguirre-León, A., A. Yáñez-Arancibia y F. Amezcua-Linares. 1982. Taxonomía, diversidad, distribución y abundancia de las mojarras de la laguna de Términos, sur del Golfo de México (Pisces: Gerreidae). An. Inst. Cienc. Mar. Limnol. Univ. Nal. Autón. México, 9: 213-250. [ Links ]
2 APHA, AWWA y WEF. 2005. Standard methods for the examination of water and wastewater. Edición 20, American Public Health Association (APHA), American Water Works Association (AWWA) y Water Environment Federation (WEF), Washington D. C. 1217 p. [ Links ]
3 Arenas-Granados, P. y A. Acero P. 1992. Organización trófica de las mojarras (Pisces: Gerreidae) de la Ciénaga Grande de Santa Marta (Caribe colombiano). Rev. Biol. Trop., 40 (3): 287-302. [ Links ]
4 Arenas-Granados, P. y A. Acero P. 1993. Taxonomía y distribución de las mojarras (Pisces: Perciformes: Gerreidae) de la Ciénaga Grande de Santa Marta (Colombia). Bol. Ecotrópica, 26: 1-19. [ Links ]
5 Bernstein, B. B. y J. Zalinski. 1983. An optimum sampling design and power test for environmental biologists. J. Environ. Manag., 16: 35-43. [ Links ]
6 Beverton, R. J. H. y S. J. Holt. 1957. On the dynamics of exploited fish populations. Fish. Invest. Minist. Agric. Fish. Food. G.B. (2 Sea Fish.), 19: 533 p. [ Links ]
7 Blanco, J. A., E. A. Viloria y J. C. Narváez. 2006. ENSO and salinity changes in the Ciénaga Grande de Santa Marta coastal lagoon system, Colombian Caribbean. Est. Coast. Shelf Sci., 66: 157-167. [ Links ]
8 Blanco, J. A., J. C Narváez B. y E. A. Viloria. 2007. Enso and the rise and fall of a tilapia fishery in northern Colombia. Fish. Res., 88: 100-108. [ Links ]
9 Boreman, J., B. S. Nakashima, J. A. Wilson y R. L. Kendall. 1997. Northwest Atlantic groundfish: Perspectives on a fishery collapse. Am. Fish. Soc., 242 p. [ Links ]
10 Botero, L. y J. E. Mancera-Pineda. 1996. Síntesis de los cambios de origen antrópico ocurridos en los últimos 40 años en la Ciénaga Grande de Santa Marta (Colombia). Rev. Acad. Colomb. Cienc., 20 (78): 465-474. [ Links ]
11 Botero, L. y H. Salzwedel. 1999. Rehabilitation of the Ciénaga Grande de Santa Marta, a mangroveestuarine system in the Caribbean coast of Colombia. Ocean Coast. Manag., 42: 243-256. [ Links ]
12 Breitburg, D. L. 1992. Episodic hipoxia in Cheasepeake Bay: interacting effects of recruitment, behavior, and physical disturbance. Ecol. Monogr., 62 (4): 525-546. [ Links ]
13 Brenden, T. O. y J. R. Bence. 2008. Comment: Use of piecewise regression models to estimate changing relationships in fisheries. North Am. J. Fish. Manag., 28 (3): 844-846. [ Links ]
14 Burgos, R. y O. Defeo. 2004. Long-term population structure, mortality and modeling of a tropical multi-fleet fishery: the red grouper Epinephelus morio of the Campeche Bank, Gulf of Mexico. Fish. Res., 66: 325-335. [ Links ]
15 Carranza, A., O. Defeo y M. Beck. 2009. Diversity, conservation status and threats to native oysters (Ostreidae) around the Atlantic and Caribbean coasts of South America. Aq. Cons. Mar. Freshwater Ecosyst., 19: 344-353. [ Links ]
16 Cervigón, F., R. Cipriani, W. Fischer, L. Garibaldi, M. Hendrickx, A. J. Lemus, R. Márquez, J. M. Poutiers, G. Robaina y B. Rodríguez. 1992. Guía de campo de las especies marinas y de aguas salobres de la costa septentrional de Sur América. FAO, Roma. 513 p. [ Links ]
17 Díaz, S. y M. Cabido. 2001. Vive la differénce: plant functional diversity matters to ecosystem processes. Trends in Ecology and Evolution, 16: 646-655. [ Links ]
18 FAO. 2007. El estado mundial de la pesca y la acuicultura 2006. Departamento de Pesca y Acuicultura de la FAO, Roma. 194 p. [ Links ]
19 Folke, C., S. Carpenter, B. Walker, M. Scheffer, T. Elmqvist, L. Gunderson y C. S. Holling. 2005. Regime shifts, resilience, and biodiversity in ecosystem management. Ann. Rev. Ecol. Evol. Syst., 35 (1): 557-581. [ Links ]
20 García, C. B., L. O. Duarte, J. Altamar y L. Manjarrés. 2007. Demersal fish density in the upwelling ecosystem off Colombia, Caribbean Sea: Historic outlook. Fish. Res., 85: 68-73. [ Links ]
21 Gray, J. R., G. D. Glysson, L. M. Turcios y G. E. Schwarz. 2000. Comparability of suspendedsediment concentration and total suspended solids data. U.S. Geological Survey Water-Resources Investigations Report 00-4191, Reston, EE. UU. 14 p. [ Links ]
22 Harper, K., I. Bergeron, P. Drapeau, S. Gauthier y L. De Grandpré. 2005. Structural development following fire in a black spruce boreal forest. Forest Ecol. Manag., 206: 293-306. [ Links ]
23 Hernández, C. y G. Márquez. 1987. Potencial cosechable de la ostra Crassostrea rhizophorae (Guilding), en la Ciénaga Grande de Santa Marta, Colombia. Acta Biol. Col., 1 (3): 25-40. [ Links ]
24 Hinchey, E. K., L. C. Hinchey, L. C. Schaffner, C. C. Hoar, B. W. Vogt y L. P. Batte, 2006. Responses of estuarine benthic invertebrates to sediment burial: the importance of mobility and adaptation. Hydrobiology, 556:85-98. [ Links ]
25 Holley, M. P., M. D. Marshall y M. J. Maceina. 2009. Fishery and population characteristics of blue catfish and channel catfish and potential impacts of minimum length limits on the fishery in Lake Wilson, Alabama. North Am. J. Fish. Manag., 29 (4): 1183-1194. [ Links ]
26 Holme, N. y A. McIntyre. 1971. Methods for the study of marine benthos. Blackwell Sci. Publ., Oxford. 334 p. [ Links ]
27 Hudson, D. J. 1966. Fitting segmented curves whose join points have to be estimated. J. Am. Stat. Ass., 61: 1097-1129. [ Links ]
28 Hutchings, J. A. 2000. Collapse and recovery of marine fishes. Nature, 406: 882-885. [ Links ]
29 Hutchings, J. A. 2004. Marine fish population collapses: consequences for recovery and extinction risk. BioScience, 54 (4): 297-309. [ Links ]
30 Imarpe. 2004. Anchoveta. <http://www.imarpe.gob.pe/imarpe/pelagicos/images/Anchove_170604.pdf>. 28/11/2007. [ Links ]
31 Imarpe. 2005. Seguimiento de pesquerías y evaluación de recursos pesqueros. <http://www.imarpe.gob.pe/imarpe/transparen/informe_plan_oper_ 3trim05.pdf>. 28/11/2007. [ Links ]
32 Jackson, J. B., M. Kirby, W. H. Berger, K. A. Bjorndal, L. W. Botsford, B. J. Bourque, R. H. Bradbury, R. Cooke, J. Erlandson, J. A. Estes, T. P. Hughes, S. Kidwell, C. B. Lange, H. S. Lenihan, J. M. Pandolfi, C. H. Peterson, R. S. Steneck, M. J. Tegner y R. R. Warner. 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science, 293 (5530): 629-638. [ Links ]
33 Jha, M. N. 2007. Framework for developing suspended and bedded sediment (SABS). Water Quality Criteria. Sustaining Colorado Watershed. US EPA Region-8. Breckenridge, EE. UU. 72 p. [ Links ]
34 Kaufmann, R. y F. Hevert. 1973. El régimen fluviométrico del río Magdalena y su importancia para la Ciénaga Grande de Santa Marta. Mitt. Inst. Colombo-Alemán Invest. Cient., 7: 121-138. [ Links ]
35 Klanjscek, J. y T. Legovic. 2007. Is anchovy (Engraulis encrasicolus, L.) overfished in the Adriatic Sea? Ecological Model., 201: 312-316. [ Links ]
36 Leal-Flórez, J., M. Rueda y M. Wolff. 2008. Role of the non-native fish Oreochromis niloticus in the longterm variations of abundance and species composition of the native ichthyofauna in a Caribbean estuary. Bull. Mar. Sci., 82 (3): 365-380. [ Links ]
37 Longhurst, A. 2007. Doubt and certainty in fishery science: Are we really headed for a global collapse of stocks? Fish. Res., 86: 1-5. [ Links ]
38 Maceina, M. J. 2007. Use of piecewise nonlinear models to estimate variable size-related mortality rates. North Am. J. Fish. Manag., 27: 971-977. [ Links ]
39 Maceina, M. J. y R. H. Hunter. 2008. Use of piecewise regression models to estimate changing relationships in fisheries: Response to comment. North Am. J. Fish. Manag., 28 (3): 847-848. [ Links ]
40 Mancera, J. y J. Mendo. 1996. Population dynamics of the oyster Crassostrea rhizophorae from the Ciénaga Grande de Santa Marta, Colombia. Fish. Res., 26: 139-148. [ Links ]
41 Maurer, B. A. 1998. Ecological science and statistical paradigms: at the threshold. Science, 279: 502-503. [ Links ]
42 McGee, V. E. y W. T. Carleton. 1971. Piecewise regression. J. Am. Stat. Ass., 65 (331): 1109-1124. [ Links ]
43 Mullon, C., P. Fréon, y P. Cury. 2005. The dynamics of collapse in world fisheries. Fish and Fisheries, 6 (2): 111-120. [ Links ]
44 Narváez, J. C., M. Rueda, E. Viloria, J. Blanco, J. A. Romero y F. Newmark. 2005. Manual del Sistema de Información Pesquera del Invemar (Sipein V.3.0): Una herramienta para el diseño de sistemas de manejo pesquero. Serie de documentos generales No. 18, Instituto de Investigaciones Marinas y Costeras, Santa Marta. 128 p. [ Links ]
45 Narváez, J. C., F. A. Herrera y J. Blanco. 2008. Efecto de los artes de pesca sobre el tamaño de los peces en una pesquería artesanal del Caribe colombiano. Bol. Invest. Mar. Cost., 37 (2): 163-187. [ Links ]
46 Narváez B., J. C., J. Blanco R., E. Viloria M., M. Santos A. y D. L. Gil A. 2011. La tilapia del Nilo Oreochromis niloticus (Pisces: Cichlidae) en Ciénaga Grande de Santa Marta y Complejo de Pajarales 104-117. En: Gracia, A., J. Medellín-Mora, D. L. Gil-Agudelo y V. Puentes (Eds.). Guía de las especies introducidas marinas y costeras de Colombia. Instituto de Investigaciones Marinas y Costeras (Invemar) y Ministerio de Ambiente y Desarrollo Sostenible, Santa Marta. 136 p. [ Links ]
47 Nikolsky, G. V. 1963. The ecology of fishes. Academic Press, Londres. 328 p. [ Links ]
48 Olaya-Nieto, C. W. y R. S. Appeldoorn. 2004. Edad y crecimiento de la mojarra rayada, Eugerres plumieri (Cuvier) en la Ciénaga Grande de Santa Marta, Colombia. Proc. Gulf Caribb. Fish. Inst., 55: 337-347. [ Links ]
49 Overholtz, W. J. 2002. The Gulf of Maine-Georges Bank Atlantic herring (Clupea harengus): spatial pattern analysis of the collapse and recovery of a large marine fish complex. Fish. Res., 57: 237-254. [ Links ]
50 Parra, G. J. 1996. Biología de la mojarra rayada Eugerres plumieri (Cuvier, 1830) (Pisces: Gerreidae) en la bahía de Cispatá, golfo de Morrosquillo, Caribe colombiano. Tesis Biología. Univ. Andes, Bogotá. 139 p. [ Links ]
51 Pauly, D. e I. Tsukayama (Eds.). 1987. The Peruvian anchoveta and its upwelling ecosystem: Three decades of change. Imarpe, GTZ e ICLARM. Callao, Perú. 351 p. [ Links ]
52 Pielou, E. C. 1977. Mathematical ecology. John Wiley, Nueva York. 385 p. [ Links ]
53 Prasad, A. K., L. Chai, R. P. Singh y M. Kafatos. 2006. Crop yield estimation for Iowa using remote sensing and surface parameters. Internat. J. App. Earth Observ. Geoinf., 8: 26-33. [ Links ]
54 Puyana, M. 1995. Aspectos biológicos y ecológicos de Mytilopsis sallei (Recluz, 1849) (Bivalvia: Dreissenidae) en bancos de ostras de la Ciénaga Grande de Santa Marta, Caribe colombiano. An. Inst. Invest. Mar. Punta Betín, 24: 39-53. [ Links ]
55 Randall, J. E. 1967. Food habits of reef fishes of the West Indies. Stud. Trop. Oceanogr., 5: 665-847. [ Links ]
56 Restrepo, J. D y B. Kjerve. 2000. Magdalena river: interannual variability (1975-1995) and revised water discharge and sediment load estimates. J. Hydrol., 235: 137-149. [ Links ]
57 Ricker, W. E. 1975. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Canada, (191), 382 p. [ Links ]
58 Rivera-Monroy, V. H., R. R. Twilley, E. Mancera, E. Castañeda, O. Casas, F. Daza, J. Restrepo, L. Perdomo, P. Reyes, M. Villamil y F. Pinto. 2001. Estructura y función de un ecosistema de manglar a lo largo de una trayectoria de restauración en diferentes niveles de perturbación. Informe técnico. MAVDT, Invemar y Colciencias, Santa Marta. 331 p. [ Links ]
59 Rivera-Monroy, V. H., R. R. Twilley, E. Mancera, A. Alcántara-Eguren, E. Castañeda-Moya, O. Casas, P. Reyes, J. Restrepo, L. Perdomo, E. Campos, G. Cotes y E. Viloria. 2006. Aventuras y desventuras en Macondo: Rehabilitación de la Ciénaga Grande de Santa Marta, Colombia. Ecotrópicos 19 (2): 72-93. [ Links ]
60 Rueda, M. 2007. Evaluating the selective performance of the encircling gillnet used in tropical estuarine fisheries from Colombia. Fish. Res., 87: 28-34. [ Links ]
61 Rueda, M. 2001. Spatial distribution of fish species in a tropical estuarine lagoon: a geostatistical appraisal. Marine Ecology Progress Series, 222: 217-226. [ Links ]
62 Rueda, M. y O. Defeo. 2001. Survey abundance indices in a tropical estuarine lagoon and their management implications: a spatially-explicit approach. ICES Journal of Marine Science, 58: 1219-1231. [ Links ]
63 Rueda, M. y O. Defeo. 2003. Linking fishery management and conservation in a tropical estuarine lagoon: biological and physical effects of an artisanal fishing gear. Est. Coast. Shelf Sci., 56: 935-942. [ Links ]
64 Rueda, M. y A. Santos-Martínez. 1997. Evaluación de la eficiencia y selectividad de la red bolichera en la pesquería de la Ciénaga Grande de Santa Marta. Caribe colombiano. Bol. Inv. Mar. Cost., 26: 17-34. [ Links ]
65 Rueda, M. y A. Santos-Martínez. 1999. Population dynamics of the striped mojarra Eugerres plumieri from the Ciénaga Grande de Santa Marta, Colombia. Fish. Res., 42: 155-166. [ Links ]
66 Rueda, M. E., J. E. Mancera y J. Mendo. 1997. Estimación del factor de retención de la red bolichera empleada en la pesquería de la Ciénaga Grande de Santa Marta, Caribe colombiano. Rev. Acad. Colomb. Cienc., 21 (81): 487-495. [ Links ]
67 Sánchez, C. y M. Rueda. 1999. Space-time variation of the diversity and abundance of dominant fish species in the Magdalena River Delta, Colombia. Rev. Biol. Trop., 47: 1067-1079. [ Links ]
68 Santos-Martínez, A. y A. Acero. P. 1991. Fish community of the Ciénaga Grande de Santa Marta (Colombia), composition and zoogeography. Icthyol. Explor. Freshwaters, 2: 247-263. [ Links ]
69 Santos-Martínez, A. y E. Viloria. 1998. Evaluación de los recursos pesqueros de la CGSM y CP. Caribe colombiano. Estadística pesquera nov/93-oct/96 (Reportes de la pesquería). 1-62. En: Santos-Martínez, A. y E. Viloria (Eds.). Evaluación principales recursos pesqueros CGSM, Costa Caribe colombiana Tomo I. Informe final, Colciencias, Invemar y GTZ Prociénaga, Santa Marta. 200 p. [ Links ]
70 Smith, E. P. 2002. BACI design. Encyclopedia of environmetrics, 1: 141-148. [ Links ]
71 Smith, T. D. y J. Link. 2005. Autopsy your dead and living fisheries: a proposal for fisheries science, fisheries management and fisheries. Fish and Fisheries, 6: 73-87. [ Links ]
72 StatSoft, Inc. 2003. Statistica (data analysis and Software system) version 6.1. <http://www.statsoft.com>. 28/11/2008. [ Links ]
73 Stone, H. H., S. Gavaris, C. H. M. Legault, J. D. Neilson y S. X. Cadrin. 2004. Collapse and recovery of the yellowtail flounder (Limanda ferruginea) fishery on Georges Bank. J. Sea Res., 51: 261-270. [ Links ]
74 Toms, J. D. y M. L. Lesperance. 2003. Piecewise regression: a tool for identifying ecological thresholds. Ecology, 84 (8): 2034-2041 p. [ Links ]
75 Underwood, A. J., 1991. Beyond BACI: experimental designs for detecting human environmental impacts on temporal variations in natural populations. Aust. J. Mar. Freshwater Res., 42: 569-587. [ Links ]
76 Walters, C. J. y S. J. D. Martell. 2004. Fisheries ecology and management. Princeton University Press, New Jersey, EE. UU. 399 p. [ Links ]
77 Wiedemann, H. U. 1973. Reconnaissance of the Ciénaga Grande de Santa Marta, Colombia: Physical parameters and geological history. Mitt. Inst. Colombo-Alemán Invest. Cient., 7: 85-119. [ Links ]
FECHA DE RECEPCIÓN: 16/05/2011 FECHA DE ACEPTACIÓN: 23/09/2012
*Contribución No. 1113 del Instituto de Investigaciones Marinas y Costeras -Invemar- y No. 375 del Centro de Estudios en Ciencias del Mar -Cecimar- de la Universidad Nacional de Colombia sede Caribe.
