










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.42 no.1 Santa Marta Jan./June 2013
MOLUSCOS ASOCIADOS A ENSAMBLAJES MACROALGALES EN EL LITORAL ROCOSO DE CÓRDOBA, CARIBE COLOMBIANO*
MOLLUSCS ASSOCIATED WITH MACROALGAE ASSEMBLAGES ON THE ROCKY COASTLINE OF CÓRDOBA, COLOMBIAN CARIBBEAN
Jorge Quirós Rodríguez1 y Néstor Hernando Campos2
1 Universidad de Córdoba, Facultad de Ciencias Básicas e Ingenierías, Departamento de Biología. Carrera 6 No. 76-103, Montería, Colombia. alexander_quiroz@hotmail.com.
2 Universidad Nacional de Colombia sede Caribe, Centro de Estudios en Ciencias del Mar (Cecimar), Santa Marta, Colombia. nhcamposc@unal.edu.co.
RESUMEN
La composición y abundancia de la comunidad bentónica de moluscos asociada a ensamblajes macroalgales fueron estudiadas en dos ecorregiones de la franja costera del departamento de Córdoba, Caribe colombiano. Entre septiembre de 2006 y junio de 2007 fueron realizados cuatro muestreos en seis estaciones; tres en San Antero, dos en Puerto Escondido y una en Los Córdobas. Para la recolección de los especímenes se delimitó un cuadrante de 625 cm2 con cinco repeticiones dispuestas al azar en cada estación. Se recolectaron un total de 1012 especímenes y 68 especies de moluscos fueron identificadas (46 gastrópodos, 19 bivalvos, dos poliplacóforos y un escafópodo). Las especies más abundantes en la ecorregión Morrosquillo fueron Bittiolum varium (34.95%), Parvanachis obesa (13.69%), Isognomon alatus (12.43%) y Columbella mercatoria (7.93%), mientras que en la ecorregión Darién fueron Eulithidium tessellatum (38.29%), E. affine (19.04%), C. mercatoria (7%), I. alatus (5.91%) y Tegula viridula (5.69%). Se presentaron diferencias significativas entre la cobertura macroalgal (Fs = 71.2, p < 0.001) y la abundancia de moluscos (Fs = 56.9, p < 0.001) con las estaciones de muestreo (α = 0.05). El análisis de componentes principales mostró que la abundancia de P. obesa, B. varium, I. alatus y C. mercatoria está regulada por las diferencias en cobertura de las macroalgas de la familia Gracilariaceae en la ecorregión Morrosquillo, mientras que la de E. affine, E. tessellatum y T. viridula está regulada por la cobertura de las macroalgas Sargassum polyceratium y Bryothamnion triquetrum en la ecorregión Darién.
PALABRAS CLAVE: Moluscos, ensamblajes macroalgales, franja costera, ecorregiones, Caribe colombiano.
ABSTRACT
The composition and abundance of the benthic community of mollusks associated to macroalgal assemblages was studied in two ecoregions of the coastal strip of the Córdoba Province, Colombian Caribbean. Between September 2006 and June 2007 four samplings were done in six stations; three in San Antero, two in Puerto Escondido, and one in Los Córdobas. A quadrant of 625 cm2 with five replicates was used in order to collect the material; it was arranged randomly in each sampling period. A total of 1012 specimens and 68 species of mollusks were identified (46 gastropods, 19 bivalves, two polyplacophorans, and one scaphopod). The most abundant species in the Morrosquillo ecoregion were Bittiolum varium (34.95%), Parvanachis obesa (13.69%), Isognomon alatus (12.43%), and Columbella mercatoria (7.93%), while in the Darien ecoregion they were Eulithidium tessellatum (38.29%), E. affine (19.04%), C. mercatoria (7%), I. alatus (5.91%), and Tegula viridula (5.69%). There were significant differences between the macroalgal cover (Fs = 71.2, p < 0.001) and mollusk abundance (Fs = 56.9, p < 0.001) to the sampling stations (α = 0.05). The Principal Component Analysis (PCA) showed that the abundances of P. obesa, B. varium, I. alatus, and C. mercatoria is regulated by differences in coverage of the family Gracilariaceae in Morrosquillo ecoregion, while the abundance of E. affine, E. tessellatum, and T. viridula is regulated by changes in coverage of Sargassum polyceratium and Bryothamnion triquetrum in Darien ecoregion.
KEY WORDS: Mollusks, macroalgae assemblages, coastal strip, ecoregions, Colombian Caribbean.
INTRODUCCIÓN
Los moluscos son un grupo de organismos que se encuentran asociados a gran variedad de unidades ecológicas incluyendo sustratos rocosos, formaciones coralinas, pastos marinos y ensamblajes macroalgales (García-Ríos, 1983; Márquez y Jiménez, 2002; García-Ríos et al., 2008). Estos últimos están constituidos por una comunidad heterogénea de algas filamentosas, frondosas, corticadas y coriáceas que constituyen un hábitat que alberga gran diversidad de moluscos debido a la alta producción de materia orgánica que genera y a la variedad de sustratos que brindan refugio, alimento y espacio para estos organismos (Shacklock y Doyle, 1983; Niam, 1988). El Atlántico occidental presenta alrededor de 1240 especies de algas marinas (Wynne, 1998). No obstante, la flora del Caribe colombiano está integrada por aproximadamente unas 550 especies (Díaz-Pulido, 2000), representando cerca de un 45% de la flora del Atlántico occidental tropical. Esta importante riqueza algal presente en el Caribe colombiano parece estar relacionada con la gran diversidad de hábitats marinos, presencia de surgencias e historias geológicas variadas (Quirós-Rodríguez et al., 2010). Las comunidades de moluscos asociadas a ensamblajes algales en el Caribe han sido descritas por Warmke y Almodóvar (1963) y García-Ríos (1983). En el primero de estos trabajos, se recolectaron 25 especies de algas en el suroeste de Puerto Rico, en las que estaban presentes representantes de 90 especies de moluscos. El segundo autor describió las comunidades de gasterópodos asociadas a macroalgas marinas, que se recolectaron en cuatro hábitats: costa rocosa, arrecife de coral, manglar expuesto al oleaje y canales de mangle. García-Ríos et al. (2008) describieron las comunidades de moluscos gasterópodos asociados al alga verde calcárea Halimeda opuntia, encontrando 526 gasterópodos pertenecientes a 54 especies.
En Colombia el poco conocimiento que se tiene sobre las asociaciones de grupos de invertebrados en ensamblajes algales se hace evidente por las escasas referencias en el tema. El propósito del presente estudio es determinar la composición y estructura de las comunidades de moluscos asociadas a los ensamblajes algales del litoral rocoso del departamento de Córdoba (Colombia), evaluando aspectos como riqueza específica, abundancia y diversidad, así como comparar las comunidades asociadas que se encuentran en el litoral rocoso a lo largo de 140 km de costa donde se establecen dos ecorregiones diferentes.
ÁREA DE ESTUDIO
La investigación se desarrolló en el litoral Caribe del departamento de Córdoba en las coordenadas 9°25'45"N y 75°42'21"W (corregimiento de El Porvenir, municipio de San Antero) y 8°54'86"N y 76°26'20"W (Punta Arboletes, municipio de Los Córdobas) (Figura 1). El litoral costero cordobés corresponde en general a una zona de bosque seco tropical, según el patrón de zonación de Höldridge; además presenta dos épocas climáticas al año, una seca que abarca los meses de diciembre hasta inicios de abril y otra lluviosa, la cual va desde finales de abril hasta noviembre. La extensión de la franja costera de Córdoba es de 140 km y recibe la influencia de dos ecorregiones con características propias en cuanto a su geología, hidrología, flora y fauna (Invemar, 2000). El sector correspondiente al municipio de San Bernardo del Viento no presenta una zona intermareal rocosa consolidada, por lo que en su área no se establecieron puntos de muestreo.
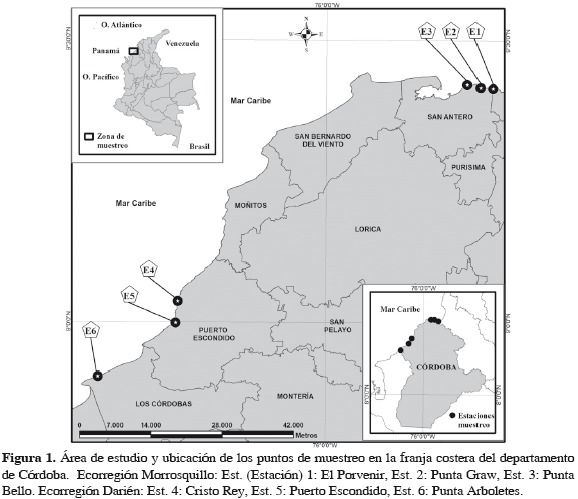
MATERIALES Y MÉTODOS
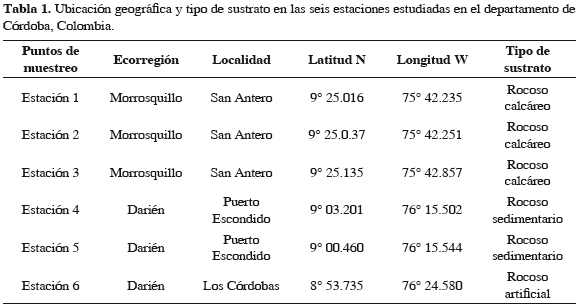
Se realizaron cuatro muestreos trimestrales entre septiembre de 2006 y junio de 2007, con el fin de abarcar los dos períodos climáticos del año que se dan en las ecorregiones de estudio. En cada punto de muestreo seleccionado se establecieron las coordenadas geográficas mediante un geoposicionador (GPS) Garmin Etrex Legend (Tabla 1). Para la selección de las estaciones de muestreo en cada área geográfica se tuvieron en cuenta varios criterios como el tipo de sustrato dominado por algas, grado de intervención antrópica, aporte de aguas continentales y vías de acceso. En cada punto de muestreo se ubicó un cinturón de cuadrantes, en cada uno de los cuales se tomaron cinco réplicas con un marco de 625 cm2 de acuerdo con lo establecido por Connor y Adey (1977) y distribuidos en seis estaciones entre 0 y 1 m de profundidad; tres estaciones se ubicaron en la ecorregión Morrosquillo y tres en Darién, cubriendo sustrato rocoso calcáreo y sedimentario. En todos los cuadrantes se evaluó la cobertura de las algas a nivel específico. Debido a la distribución de la cobertura algal sobre el sustrato rocoso, la ubicación del cinturón de cuadrantes se realizó de manera preferencial aleatoria (i.e. sobre sectores homogéneos de vegetación). El valor de cobertura obtenido corresponde a un valor relativo de la cobertura algal sobre el sustrato rocoso, puesto que se ubicaron los cuadrantes preferentemente sobre el sustrato dominado por algas. Para la separación de las macroalgas desde su disco de fijación en el sustrato se empleó una espátula metálica, teniendo cuidado de extraer solo la parte superior del disco, para de esta manera eliminar los restos de conchas que pudieran hacer parte del sustrato de fijación (Warmke y Almodóvar, 1963), siendo inmediatamente colocadas en bolsas plásticas. Posteriormente, las macroalgas se ubicaron en un recipiente plástico realizando una primera separación con pinzas de punta fina de los moluscos adheridos en su estructura vegetativa (Quirós, 2009).
En el laboratorio, las macroalgas fueron lavadas evitando que algunos ejemplares quedaran atrapados. El producto de los lavados se pasó sobre tamices de 500 y 250 μm con el fin de separar los individuos por su tamaño. Los especímenes separados se colocaron en frascos independientes según su clase taxonómica y se preservaron en etanol al 70% (Warmke y Almodóvar, 1963; Quirós, 2009). La identificación hasta especie de la mayoría de los ejemplares se basó en las características externas de la concha, utilizando como referencia la literatura de Warmke y Abbott (1975), Bullock y Franz (1994), Díaz y Puyana (1994), Abbott y Morris (1995), Lodeiros et al. (1999) y Western Atlantic Mollusks Database (Malacolog. Versión: 4.1.1 http://erato.acnatsci.org/wasp/findsnail.php). El material identificado hace parte del catálogo de moluscos de la sala de colecciones de la Universidad de Córdoba en Montería.
Para el inventario de las especies macroalgales se realizaron colectas de especímenes en frascos plásticos de 250 mL con una solución de formol al 4% en agua de mar y bórax. Los especímenes fueron procesados y montados en cartulina para su posterior identificación, la cual se realizó con base en los trabajos de Joly (1967), Bula-Meyer (1982a, 1982b) y Littler et al. (2001).
Para determinar la estructura del ensamblaje de algas y moluscos, se estimó la riqueza total (S) de los dos grupos y la abundancia absoluta como porcentaje de cobertura (%) para las macroalgas y número total de individuos (N) para los moluscos recolectados por estación. Para cuantificar la variación espacial del ensamblaje de macroalgas y moluscos se realizó un análisis de varianza de una vía, en el que los factores fueron las estaciones y los muestreos y la variable dependiente fue la cobertura algal y abundancia total promedio de moluscos, previa transformación de los datos a Log (n + 1) en función de cumplir con las suposiciones de normalidad (prueba de Kolmogorov-Smirnov) y homogeneidad de varianzas (prueba de Bartlett). En aquellos casos para los que se detectaron diferencias significativas, se aplicó la prueba de Duncan (Steel y Torrie, 1985). Estos análisis se realizaron por medio del programa Statgraphics Centurion XV (Statpoint, 2006). Para evaluar la relación entre los ensamblajes algales y la variación de la riqueza y abundancia de la malacofauna asociada, se realizó un análisis de componentes principales (ACP), mediante el programa XLSTAT versión 1.0.1 (Addinsoft, 2011), utilizando como identificadores, a las seis estaciones ubicadas en franja costera rocosa intermareal del departamento de Córdoba y como variables, el porcentaje de cobertura algal y la abundancia total promedio de la malacofauna asociada.
RESULTADOS
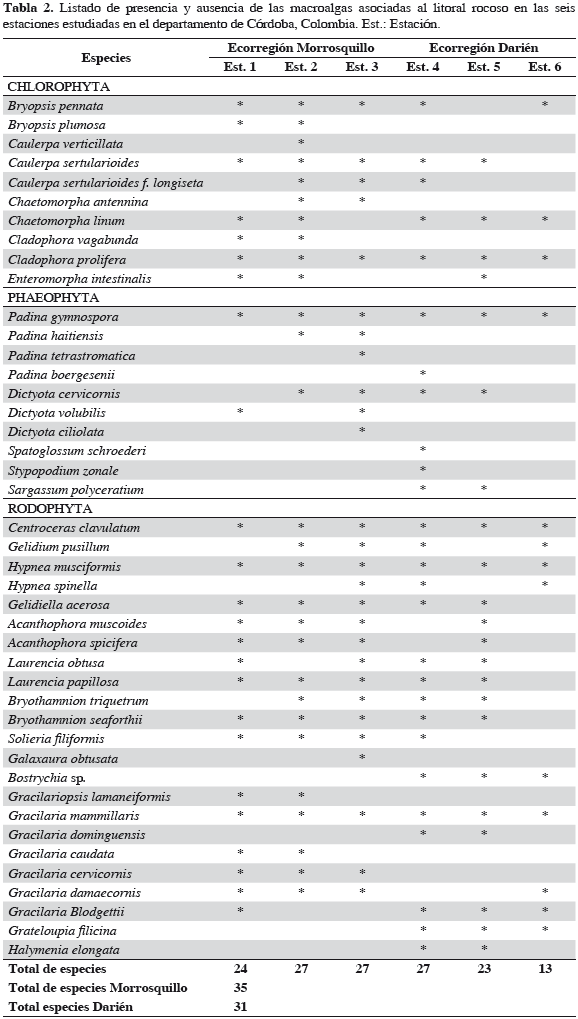
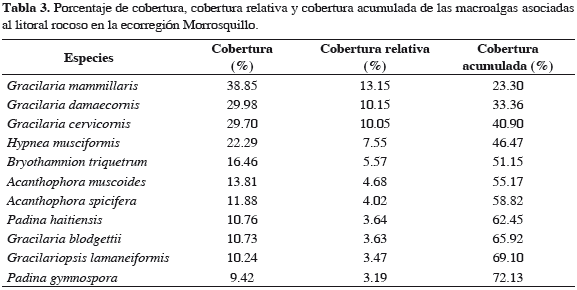
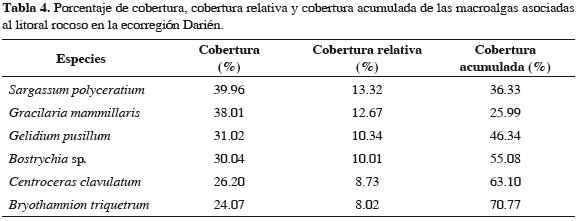
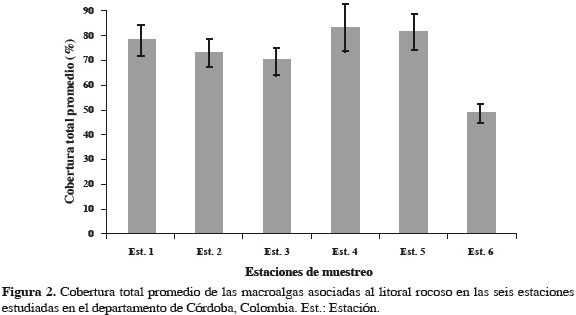
Se registraron representantes de un total de 35 especies de algas, asociadas al litoral rocoso de la ecorregión Morrosquillo; 10 son clorófitas, seis feófitas y 19 rodófitas. Los géneros con mayor número de especies fueron Gracilaria (5), Padina (3), Dictyota (3) y Caulerpa (3) (Tabla 2); Gracilaria mammillaris fue la especie más abundante (13.2%), seguida de G. damaecornis (10.2%), G. cervicornis (10.1%), Hypnea musciformis (7.6%), Bryothamnion triquetrum (5.6%), Acanthophora muscoides (4.7%), A. spicifera (4.0%), Padina haitiensis (3.6%), G. blodgettii (3.6%), Gracilariopsis lamaneiformis (3.5%) y P. gymnospora (3.2%), representando 72.1% del total de las especies registradas en el sector (Tabla 3). En la ecorregión Darién se registraron representantes de un total de 31 especies asociadas al litoral rocoso; seis son clorófitas, seis feófitas y 20 rodófitas. El género con mayor número de especies fue Gracilaria (4) (Tabla 2); Sargassum polyceratium fue la especie más abundante (13.3%) , seguida de G. mammillaris (12.7%), Gelidium pusillum (10.3%), Bostrychia sp. (10.0%), Centroceras clavulatum (8.7%) y B. triquetrum (8.0%), constituyendo el 70.8% del total de las especies registradas para el sector (Tabla 4). A nivel espacial, se establecieron diferencias significativas según el análisis de varianza a una vía (Fs = 71.2, p < 0.001), con base en la cobertura total promedio y las estaciones de muestreo. Al aplicar sobre estos resultados una prueba a posteriori de Duncan, se formaron tres grupos: la estación 6 forma un solo grupo, con la media más baja. Las estaciones 2 y 3 forman el segundo grupo, con medias relativamente altas, y el tercer grupo lo forman las estaciones 1, 4 y 5 con las medias más altas. El porcentaje más alto en la cobertura total promedio espacial de macroalgas fue para la estación 4 con 83.5%, mientras la 6 registró el menor porcentaje con 70.3% (Figura 2).
En la ecorregión Morrosquillo se recolectó un total de 555 individuos pertenecientes a 48 especies de moluscos. Se registraron 31 familias, de las cuales Columbellidae presentó el mayor número de especies (5), seguida de Cerithiidae y Mytilidae (4), y Pseudomelatomidae, Muricidae, Olividae, Phasianellidae, Isognomonidae y Tellininae (2) (Tabla 5). Las familias restantes estuvieron presentes solamente con una especie. Bittiolum varium fue la especie más abundante durante el estudio (35.0%), seguida de Parvanachis obesa (13.7%), Isognomon alatus (12.4%), Columbella mercatoria (8.0%), Eulithidium tessellatum (4.5%), Astyris lunata (3.1%), Anachis lyrata (2.9%) y Chrysallida gemmulosa (2.5%) (Figura 3a). El resto de las especies para el sector aportaron una abundancia menor de 2%.
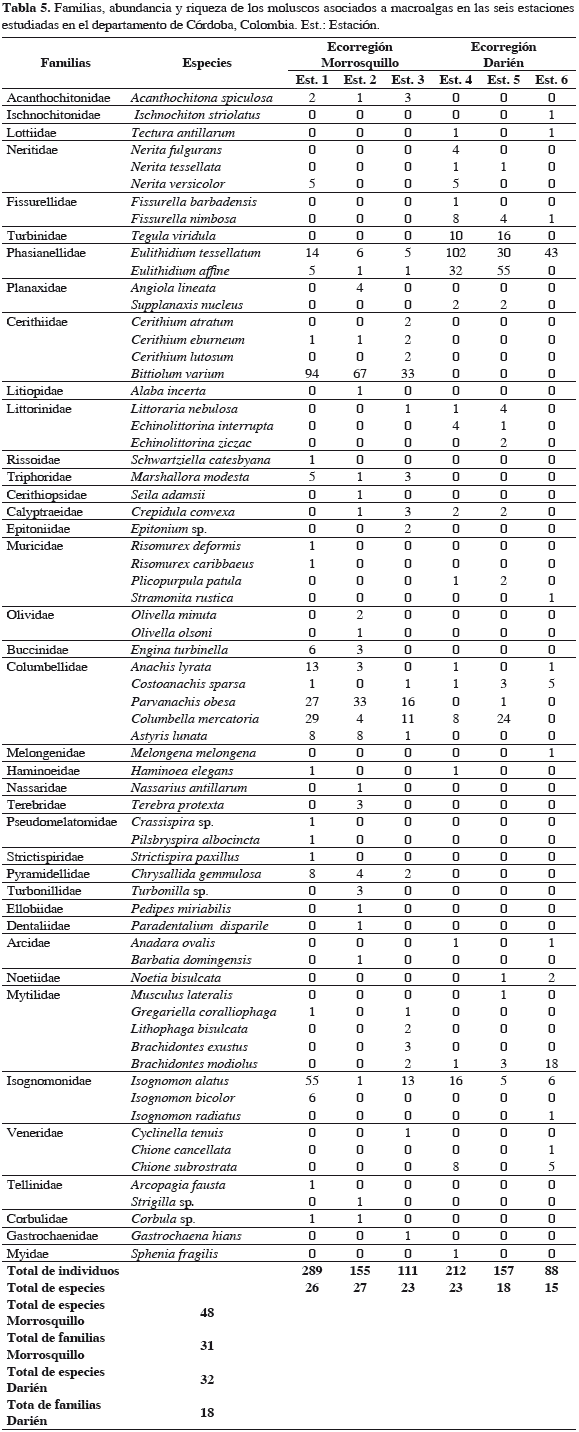
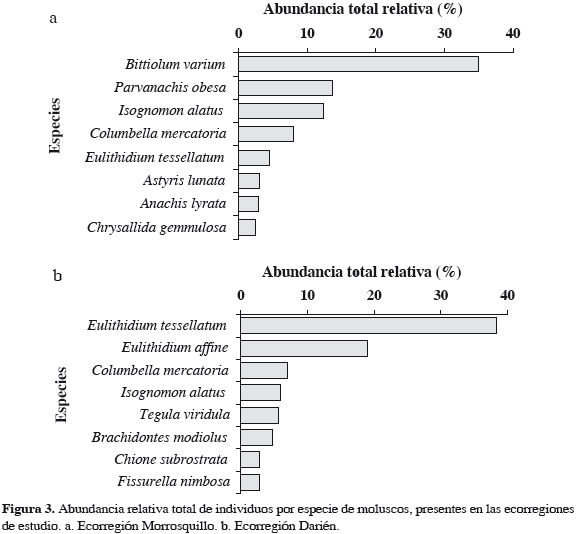
En Darién se recolectó un total de 457 individuos pertenecientes a 32 especies de moluscos. Esta ecorregión registró 18 familias, siendo igualmente Columbellidae la que presentó el mayor número de especies (4), seguida de Neritidae y Littorinidae (3) y Fissurellidae, Phasianellidae, Muricidae, Mytilidae, Isognomonidae y Veneridae (2) (Tabla 5). El resto de las especies estuvieron representadas por una especie. Eulithidium tessellatum fue la especie más abundante durante el estudio (38.3%), seguida de E. affine (19.0%), C. mercatoria (7.0%), I. alatus (5.9%), Tegula viridula (5.7%), Brachidontes modiolus (4.8%) y Chione subrostrata y Fissurella nimbosa (2.8%) (Figura 3b). El resto de las especies registraron una abundancia menor de 2%.
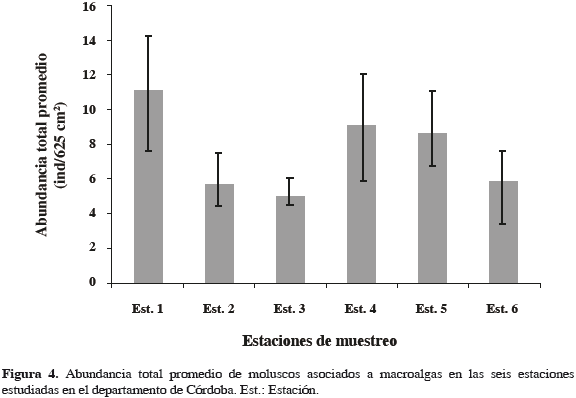
Los resultados del análisis de varianza demostraron que existen diferencias significativas entre el número de organismos promedio y las estaciones de muestreo (Fs = 56.9, p < 0.001). Al comparar las medias de la abundancia de moluscos con respecto a las estaciones de muestreo a través de la prueba a posteriori de Duncan, se presentó la formación de tres grupos. Así, las estaciones 2, 3 y 6 forman el primer grupo, con las medias más bajas, y el segundo lo forman las estaciones 4 y 5 con medias altas. El tercer grupo lo forma la estación 1 con la media más alta. La mayor abundancia total promedio, a nivel espacial, fue para la estación 1 con 11.1 ± 3.09 ind/625 cm2, y el valor más bajo fue para la estación 3, con 4.8 ± 0.53 ind/625 cm2 (Figura 4).
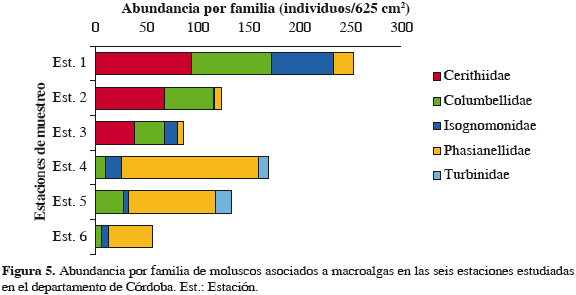
Las familias Phasianellidae, Isognomonidae y Columbellidae se registraron en todas las estaciones de muestreo. La familia Cerithiidae fue el grupo dominante en la ecorregión Morrosquillo, siendo la estación 1 la que registró el pico más alto para esta familia (95 ind/625 cm2) y la estación 3 el menor valor (39 ind/625 cm2); Phasianellidae mostró la mayor abundancia en la ecorregión Darién, siendo la estación 4 la que mostró la mayor abundancia (134 ind/625 cm2) y la 6 la que registró el menor valor (43 ind/625 cm2). La familia Turbinidae se registró únicamente en las estaciones 4 y 5 con abundancias de 10 y 16 ind/625 cm2 (Figura 5).
El Análisis de Componentes Principales (ACP), que integra las especies macroalgales de mayor cobertura y representatividad por ecorregión, muestra que G. mammillaris (1), G. damaecornis (2), G. cervicornis (3), B. triquetrum y S. polyceratium están definiendo las principales poblaciones de moluscos asociados. De acuerdo con la influencia de estos ensamblajes, se observa que la disposición de P. obesa, B. varium, I. alatus y C. mercatoria está mayormente influenciado por las diferencias en cobertura de la familia Gracilariaceae (1, 2 y 3) en las estaciones 1, 2 y 3 de la ecorregión Morrosquillo. Las especies E. affine, E. tessellatum y T. viridula conforman otro grupo que está mayormente influenciado por la cobertura de S. polyceratium y B. triquetrum en las estaciones 4, 5 y 6, pertenecientes a Darién (Figura 6).
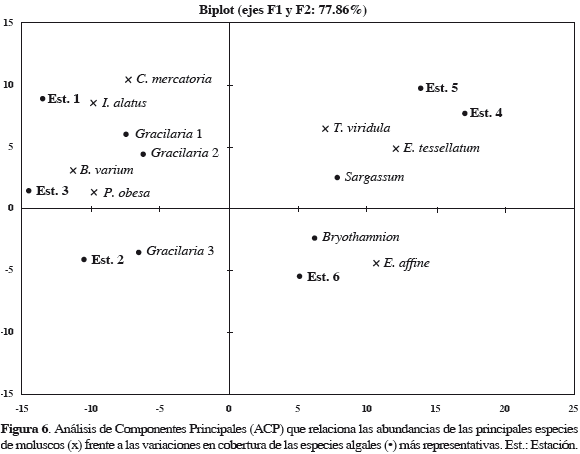
El ACP también muestra la preferencia de ciertas especies de moluscos por ocupar ambientes específicos. Se determina que las especies P. obesa, B. varium, I. alatus y C. mercatoria prefieren crecer en algas corticadas, mientras que E. affine, E. tessellatum y T. viridula prefieren algas coriáceas y en cierta manera corticadas.
DISCUSIÓN Y CONCLUSIONES
Se registraron representantes de un total de 35 y 31 especies de macroalgas asociadas al litoral rocoso de las ecorregiones Morrosquillo y Darién, respectivamente. Este número resulta relativamente bajo, comparado con inventarios conocidos en otras áreas del Caribe colombiano, como Santa Marta (309), archipiélago de San Andrés (203) y golfo de Urabá (175) (Díaz-Pulido y Bula-Meyer, 1997; Díaz-Pulido, 2000; Díaz- Pulido y Díaz-Ruíz, 2003). No obstante, es aún prematuro determinar qué tan diversa es la composición macroalgal costera de Córdoba con respecto a otras localidades, debido al escaso esfuerzo de muestreo en esta área, así como a la poca variedad de ecosistemas y microhábitats (García y Díaz-Pulido, 2006; Quirós-Rodríguez et al., 2010). Las especies registradas en el estudio presentan un amplio ámbito de distribución, encontrándose en el Caribe en general (Díaz-Pulido, 2000; Arias et al., 2006).
En la ecorregión Morrosquillo, las coberturas de Gracilaria mammillaris, G. damaecornis y G. cervicornis presentaron los mayores porcentajes. De acuerdo con Arias et al. (2006) y Quirós-Rodríguez et al. (2010), este sector a nivel de intermareal se caracteriza por presentar sustrato rocoso-coralino, importante para el establecimiento de macroalgas de características corticadas, folioso corticadas y filamentosas. Así mismo, la presencia de sedimentos provenientes de arroyos y ciénagas, así como el efecto de las descargas de materia orgánica producidas por las corrientes de arrastre del río Sinú, estimulan el crecimiento y desarrollo de comunidades de algas rojas de estas características para la ecorregión. En esta franja costera, 11 especies alcanzaron 72.1% de la cobertura total acumulada. Esta tendencia evidencia la dominancia de ciertas especies por una unidad ecológica determinada, donde la abundancia es compartida por pocas especies con adaptaciones morfológicas y fisiológicas necesarias para sobrevivir, desarrollarse y reproducirse. En Darién con solo seis especies se obtuvo un porcentaje similar, revelando una dominancia macroalgal más marcada que la ecorregión anterior. Esta condición, según Arias et al. (2006), puede ser el resultado de una escala a nivel espacial como el tipo, textura, inclinación y composición química del sustrato, y en segunda instancia puede ser regulada temporalmente por eventos climáticos como lluvias, vientos, corrientes y oleajes.
En la franja costera de Córdoba se registró un total de 1012 individuos, pertenecientes a 37 familias y 68 especies de moluscos. La gran mayoría de las especies corresponde a la clase Gastropoda (46 especies, 67.6%), seguida por Bivalvia (19 especies, 27.9%), Polyplacophora (dos especies, 2.9%) y Scaphopoda (una especie, 1.5%). Esta composición, en un área costera de 140 km, es un número relativamente bajo si se tiene en cuenta que en el Atlántico occidental tropical y el mar Caribe se encuentran entre las regiones marinas de mayor biodiversidad en el planeta debido a una alta heterogeneidad espacial y temporal en las condiciones hidrográficas, topográficas y climáticas (Díaz y Puyana, 1994; Rodríguez et al., 2003).
Este número aún resulta relativamente bajo comparado con los resultados de Díaz y Puyana (1994) para el Caribe colombiano, quienes registraron 1180 especies y el de Díaz (2004), quien registró 727 especies en Santa Marta y el Parque Nacional Natural Tayrona. Por otro lado, en el presente estudio se mostró una mayor riqueza que el trabajo realizado por García-Valencia y Díaz (2000) para el sector sur de la plataforma continental del Caribe colombiano señalando 35 especies. Sin embargo es aún anticipado determinar qué tan diversa es la malacofauna costera de Córdoba con respecto a otras localidades, debido a la escasez de estudios en el área. En la medida que se investigue la malacofauna asociada a diferentes biotopos, como formaciones coralinas, pastos marinos, litorales rocosos y fondos de arena y limo, se incrementarán los registros e información para el sector.
En el estudio se observaron cambios significativos en la cobertura macroalgal, principalmente en las estaciones 1 (El Porvenir) y 6 (Punta Arboletes). Las demás estaciones presentaron una abundancia macroalgal que se ajusta a la condición espacial de cada ecorregión. En las estaciones 2 y 3 se observó un aumento en el número de especies, especialmente debido al incremento de algas pardas.
El aumento en el número de especies en las estaciones 2 (Punta Graw) y 3 (Punta Bello) coincide con la influencia de los arroyos Bijao Chiquito y Carbonero (Molina et al., 1994). Allí también hay presencia de formaciones coralinas y praderas de fanerógamas marinas, que aportan elementos esenciales para el desarrollo de la flora algal (Quirós-Rodríguez et al., 2010). El aumento observado en estas estaciones representa los puntos de mayor diversidad macroalgal para la ecorregión. La tendencia a una disminución en el porcentaje de cobertura macroalgal en la estación 6 (Punta Arboletes) puede deberse al sustrato rocoso artificial, la alta erosión costera y a los factores dinámicos como oleaje, vientos y corrientes oceánicas.
Varias familias de moluscos (Columbellidae, Phasianellidae, Cerithiidae y Turbinidae) son características de fondos vegetales, sustrato rocoso intermareal y cascajo coralino, importantes como áreas de alimentación, protección y refugio (Houbrick, 1968; Andrews, 1977; Vokes y Vokes, 1983). Durante el estudio se registró una mayor abundancia de B. varium, P. obesa, C. mercatoria, E. tessellatum, E. affine y T. viridula, lo que sugiere que esta diversidad se puede considerar característica del sustrato macroalgal en las ecorregiones.
La presencia de sedimentos y arena en los frondes macroalgales sería una de las posibles condiciones que estarían favoreciendo la presencia de los gastrópodos, Olivella minuta y Olivella olssoni en la estación 2 de la ecorregión Morrosquillo. Edwards (1969), Hyndes et al. (2005) y Ourives et al. (2011) registraron especies de vida libre de la familia Olividae como características de fondos blandos, praderas marinas y sustratos rocosos. En el presente estudio, las macroalgas proporcionan numerosos hábitats, refugio y alimento, ofreciendo una gama de alternativas para ser colonizadas por estos gastrópodos. La presencia de sedimentos acumulados en las macroalgas, así como materia orgánica en suspensión, sería una de las condiciones que estaría favoreciendo la presencia a los bivalvos de la familia Veneridae en las estaciones 4 y 6 del Darién cordobés. Denadai et al. (2001) y Prieto et al. (2005) señalan que estos organismos son comunes en fondos blandos, ya que este tipo de hábitat les sirve como áreas de alimentación y refugio. Según McLachlan y Jaramillo (1995) y Ríos-Jara et al. (2008), además del tipo de sustrato se presentan factores de tipo biológico, como competencia por alimento, tasa de depredación y migración, las que determinan la presencia y abundancia de bivalvos en un área determinada.
Phillips y Meñez (1988) y Reise (1991) indican que las diferencias biogeográficas mediadas por los procesos abióticos y la distancia son a largo plazo; además, al aumentar la escala espacial, como lo plantean Pearson y Rosemberg (1987), las asociaciones no se repetirían a causa de los gradientes ambientales o las diferencias regionales. Esta condición se puede apreciar al comparar la composición y abundancia de la malacofauna en las seis estaciones de las dos ecorregiones de estudio.
A nivel espacial, se establecieron diferencias significativas con base en el número de organismos promedio y las estaciones de muestreo, las cuales están muy ligadas a las variaciones en la cobertura macroalgal en los puntos de muestreo. La estación 1 mostró la mayor abundancia promedio, mientras que la 6 registró la menor abundancia. Esta condición en la estación 1 muestra que cuando aumentó la cobertura en las algas rojas de la familia Gracilariaceae y Rhodomelaceae y verdes de la familia Bryopsidaceae se generaron las condiciones para un aumento en la abundancia promedio de moluscos, principalmente de las familias Columbellidae, con C. mercatoria, A. lyrata, P. obesa; Phasianellidae, con E. tessellatum y E. affine; Cerithiidae, con B. varium, e Isognomonidae, con I. alatus. En los trabajos de Ríos (1985) y Díaz y Puyana (1994) se menciona que estas especies habitan en praderas de Thalassia testudinum, entre el follaje de algas del género Sargassum, así como en algas en general. Arias et al. (2006) y Quirós-Rodríguez et al. (2010) señalan que la estación recibe la influencia del arroyo Carbonero y otros afluentes continentales, aportando una inyección de nutrientes, evento que puede estimular el desarrollo de algas rojas corticadas importantes para el establecimiento de ciertos moluscos.
La estación 6 presentó la menor abundancia de moluscos, lo cual es ocasionado por los rasgos fisiográficos del litoral y el tipo de sustrato rocoso, que favorece el aumento en cobertura de algas filamentosas, G. pusillum, C. clavulatum y Bostrychia sp., y de una corticada, G. mammillaris. Las familias más abundantes de moluscos en esta estación fueron Turbinidae, con T. viridula; Columbellidae, con C. mercatoria; y Phasianellidae, con E. tessellatum y E. affine. Houbrick (1968), Ríos (1985) y Díaz y Puyana (1994) indican que estas familias de moluscos habitan sobre fondos de T. testudinum y rocas de la zona intermareal, especialmente en agua turbias y algas. La estación se caracteriza por un litoral costero con espolones, corrientes litorales y el aporte de aguas del río Sinú y algunos tributarios secundarios como el río Jobo, que transportan un carga apreciable de sedimentos y le imprimen a estas aguas costeras un carácter turbio y reducida salinidad, propiciando las condiciones para el afloramiento de ciertas algas y con ello la aparición de moluscos que se ajustan a estas condiciones ambientales más exigentes.
De acuerdo con los criterios de Arias et al. (2006), existe una composición algal característica en la ecorregión Morrosquillo, estimulada por la presencia de formaciones coralinas, bosques de manglar y praderas de fanerógamas marinas, que aportan elementos esenciales para el desarrollo de algas rojas como G. mammillaris, G. damaecornis y G. cervicornis. Dichas algas son capaces de desarrollar hábitats de alta productividad que contribuyen con la fijación de microcrustáceos y otros pequeños invertebrados, fuente directa o indirecta de alimento para especies de moluscos carnívoros, entre los que dominan C. mercatoria y P. obesa. La presencia de B. varium significa que la ecorregión posee una alta cobertura de algas filamentosas y diatomeas epifitas que se establecen sobre las rodófitas y sirven de alimento para esta especie. Según Van Montfrans et al. (1982), este gastrópodo presenta un ámbito de distribución amplio que se proyecta desde el intermareal hasta 12 m. Además, Díaz y Puyana (1994) lo registran como un componente importante de las macroalgas; se registró a I. alatus asociado significativamente a G. mammillaris y G. damaecornis, algas que pueden acumular sedimentos y partículas orgánicas sobre su estructura vegetativa y convertirse en verdaderas zonas de alimentación para este bivalvo.
En la ecorregión Darién, T. viridula y E. tessellatum se encontraron asociados a S. polyceratium en las estaciones 4 y 5, mientras E. affine se registró en B. triquetrum en la 6. La presencia de estos gastrópodos en las algas, se fundamenta en la capacidad de colonización de extensas coberturas de macroalgas corticadas importantes para la alimentación. Según Díaz y Puyana (1994) y Ríos (1985), estas especies son raspadoras, alimentándose principalmente de la microflora epífita que se fija en los frondes de estas macroalgas. La alta abundancia de estas especies en las estaciones de muestreo respondió a eventos dinámicos propios de la zona intermareal y se ajusta a las variaciones inherentes del alga con relación a los escenarios climáticos del año. Autores como los mencionados anteriormente indican que condiciones ambientales extremas como oleajes, corrientes y ritmos mareales son adecuadas para el establecimiento de especies en el intermareal.
AGRADECIMIENTOS
Se agradece a Ana González López, Mauricio Díaz Fajardo, Emilio Isaza Vargas, Diana Herrera Gallego y Óscar Pérez por su ayuda en los muestreos y recolecta del material biológico. A Óscar Arévalo Vásquez por su colaboración con la identificación de los especímenes. Agradecemos a la Oficina de Investigación y Extensión de la Universidad de Córdoba por proveer los recursos necesarios para la realización de esta investigación. A los laboratorios de Biodiversidad y Zoología por suministrar los equipos de campo y laboratorio, y al Centro de Investigación Piscícola de la Universidad de Córdoba (CINPIC) por el apoyo con los equipos que permitieron la toma de fotografías a los especímenes.
BIBLIOGRAFÍA
1 Abbott, R. T. y P. Morris. 1995. Shells of the Atlantic and Gulf Coasts and the West Indies. Peterson Field Guides, Boston-Nueva York. 350 p. [ Links ]
2 Addinsoft. 2011. XLSTAT, Analyse de données et statistique avec MS Excel. Addinsoft, Nueva York. 117 p. [ Links ]
3 Andrews, J. 1977. Shells and shores of Texas. University of Texas, Austin, EE. UU. 365 p. [ Links ]
4 Arias, J., P. Dueñas y J. Quirós. 2006. Distribución espacio temporal de las comunidades macroalgales asociadas al litoral rocoso del departamento de Córdoba, Caribe colombiano. Univ. Córdoba, Montería, Colombia. 109 p. [ Links ]
5 Bula-Meyer, G. 1982a. Una clave para las especies del género Halimeda (Udoteaceae, Chlorophyta) del Atlántico occidental. An. Inst. Inv. Mar. Punta Betín, 12: 41-44. [ Links ]
6 Bula-Meyer, G. 1982b. Adiciones a las Clorophytas marinas del Caribe colombiano. An. Inst. Inv. Mar. Punta Betín, 12: 117-136. [ Links ]
7 Bullock, R. y C. Franz. 1994. A preliminary taxonomic survey of the chitons (Mollusca: Polyplacophora) of Isla de Margarita, Nueva Esparta, Venezuela. Mem. Soc. Cienc. Nat. La Salle, 222 (141): 9-50. [ Links ]
8 Connor, J. L. y W. H. Adey. 1977. The benthic composition, standing crop, and productivity of a Caribbean algal ridge. Atoll Res. Bull., 211: 1-15. [ Links ]
9 Denadai, M. R., A. C. Z. Amaral y A. Turra. 2001. Spatial distribution of molluscs in two anthropogenic mixed sand-rock substrates of São Sebastião Channel, São Paulo State, Brazil. Estuar. Coast. Shelf Sci., 53: 733-743. [ Links ]
10 Díaz-Pulido, G. 2000. Vegetación marina de un sector de la plataforma continental de La Guajira (Caribe colombiano). Bol. Invest. Mar. Cost., 29: 27-34. [ Links ]
11 Díaz-Pulido, G. y G. Bula-Meyer. 1997. Litorales marinos y costeros. 115-128. En: Invemar (Ed.). Informe nacional sobre el estado de la biodiversidad en Colombia: año 1997. Invemar, Santa Marta. 143 p. [ Links ]
12 Díaz-Pulido, G. y M. Díaz-Ruiz. 2003. Diversity of benthic marine algae of the Colombian Atlantic. Biota Col., 4 (2): 203-246. [ Links ]
13 Díaz, J. M. 2004. Malacofauna de la zona costera de Santa Marta y Parque Nacional Natural Tayrona, Caribe colombiano. An. Inst. Invest. Mar. Punta de Betín, 23: 15-43. [ Links ]
14 Díaz, J. M. y M. Puyana. 1994. Moluscos del Caribe colombiano. Un catálogo ilustrado. Colciencias, Fundación Natura e Invemar, Bogotá. 367 p. [ Links ]
15 Edwards, D. C. 1969. Zonation by size as an adaptation for intertidal life in Olivella biblicata. Amer. Zool., 9: 399-417. [ Links ]
16 García, C. B. y G. Díaz-Pulido. 2006. Dynamics of a macroalgal rocky intertidal community in the Colombian Caribbean. Bol. Invest. Mar. Cost., 35: 7-18. [ Links ]
17 García-Ríos, C. I. 1983. Comunidades de gastrópodos asociados a macroalgas marinas comunes en el suroeste de Puerto Rico. Tesis de Maestría, Univ. Puerto Rico, Mayagüez, Puerto Rico. 135 p. [ Links ]
18 García-Ríos, C. I., F. J. Soto-Santiago, R. J. Colón-Rivera y J. R. Medina-Hernández. 2008. Gastrópodos asociados al alga calcárea Halimeda opuntia (Udoteaceae) en Puerto Rico. Rev. Biol. Trop., 56 (4): 1665-1675. [ Links ]
19 García-Valencia, C. y J. M. Díaz. 2000. Moluscos y su taxocenosis en los fondos someros del sector sur de la plataforma continental del Caribe colombiano. Bol. Invest. Mar. Cost., 29: 73-79. [ Links ]
20 Houbrick, J. R. 1968. A survey of the litoral marine mollusks of the Caribbean coast of Costa Rica. Veliger, 11: 4-23. [ Links ]
21 Hyndes, G. A., P. S. Lavery y D. I. Walker. 2005. Does transported seagrass provide an important trophic link in unvegetated, nearshore areas? Est. Coast. Shelf Sci., 63: 633-643. [ Links ]
22 Invemar. 2000. Programa nacional de investigación en biodiversidad marina y costera (PNIBM). Invemar, Fonade y MMA, Santa Marta. 83 p. [ Links ]
23 Joly, A. 1967. Gêneros de algas marinas da Costa Atlântica Latino-Americana. Editora da Universidade de São Paulo, São Paulo. 462 p. [ Links ]
24 Littler, D. S., M. M. Little, K. E. Bucher y J. N. Norris. 2001. Marine plants of the Caribbean: a field guide from Florida to Brasil. Smithsonian Inst. Press., Washington D. C. 263 p. [ Links ]
25 Lodeiros, C., B. Marín y A. Prieto. 1999. Catálogo de moluscos marinos de las costas nororientales de Venezuela: Clase Bivalvia. Ed. Apudons, Caracas. 19 p. [ Links ]
26 Márquez, B. y M. Jiménez. 2002. Comunidad de moluscos asociados a las raíces sumergidas del mangle rojo Rhizophora mangle en el golfo de Santa Fe, Estado Sucre, Venezuela. Rev. Biol. Trop., 50: 1101-1112. [ Links ]
27 McLachlan, A. y E. Jaramillo. 1995. Zonation on sandy beaches. Oceanogr. Mar. Biol. Ann. Rev., 33: 305-335. [ Links ]
28 Molina, A., C. Molina, L. Giraldo, C. Parra y P. Chevillot. 1994. Dinámica marina y sus efectos sobre la geomorfología del golfo de Morrosquillo. Bol. Cient. CIOH, 15: 93-113. [ Links ]
29 Niam, O. 1988. Distributional pattern of mobile fauna associated with Halimeda on the Tiahura coral-reef complex (Morea, French Polynesia). Coral Reefs, 6: 237-250. [ Links ]
30 Ourives, T. M., A. E. Rizzo y G. Boehs. 2011. Composición y distribución espacial de la macrofauna bentónica en el estuario del río Cachoeira, Ilhéus, Bahía, Brazil. Rev. Biol. Mar. Oceanogr., 46 (1): 17-25. [ Links ]
31 Pearson, T. H. y R. Rosemberg. 1987. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanogr. Mar. Biol. Ann. Rev., 16: 229-311. [ Links ]
32 Phillips, R. C. y E. G. Meñez. 1988. Seagrasses. Smithson. Contr. Mar. Sci., 34: 104 p. [ Links ]
33 Prieto, A., L. Ruiz y N. García. 2005. Diversidad y abundancia de moluscos de la epifauna en la comunidad sublitoral de Punta Patilla, Venezuela. Rev. Biol. Trop., 53 (1-2): 135-140. [ Links ]
34 Quirós, J. 2009. Dinámica espacial de la composición y estructura de las poblaciones de crustáceos decápodos asociados a céspedes algales del litoral rocoso del departamento de córdoba, Caribe colombiano. Tesis de Maestría, Univ. Córdoba, Montería, Colombia. 128 p. [ Links ]
35 Quirós-Rodríguez, J., J. Arias-Ríos y R. Ruiz. 2010. Estructura de las comunidades macroalgales asociadas al litoral rocoso del departamento de Córdoba, Colombia. Caldasia, 32 (2): 339-354. [ Links ]
36 Reise, K. 1991. Mosaic cycles in the marine benthos. 61-82. En: Remmert, H. (Ed.). Ecological studies. Springer-Verlag, Berlin Heidelberg. 163 p. [ Links ]
37 Ríos, E. C. 1985. Seashells of Brazil. Fundação Cidade do Rio Grande, Fundação Universidade do Rio Grande, Museu Oceanográfico, Rio Grande, Brasil. 328 p. [ Links ]
38 Ríos-Jara, E., E. López-Uriarte y C. M. Galván-Villa. 2008. Bivalve molluscs from the continental shelf of Jalisco and Colima, Mexican Central Pacific. Am. Malacol. Bull., 26: 119-131. [ Links ]
39 Rodríguez, L., R. Vargas y J. Cortés. 2003. Biodiversidad marina de Costa Rica: Gastrópodos (Mollusca: Gastropoda) de la costa Caribe. Rev. Biol. Trop., 51 (Suppl. 3): 305-399. [ Links ]
40 Shacklock, P. F. y R. W. Doyle. 1983. Control of epiphytes in seaweed cultures using grazers. Aquaculture, 31: 141-151. [ Links ]
41 Statpoint. 2006. Statgraphics Centurion XV. Statistical exploration and modeling software. http://www.statgraphics.com. 21/10/2010. [ Links ]
42 Steel, R. y J. H. Torrie. 1985. Principles and procedures in statistics. McGraw-Hill, Nueva York. 350 p. [ Links ]
43 Van Montfrans, J., R. J. Orth y E. S. Vay. 1982. Preliminary studies of grazing by Bittium varium on eelgrass periphyton. Aq. Bot., 14: 75-89. [ Links ]
44 Vokes, H. E. y E. H. Vokes. 1983. Distribution of shallow-water marine mollusca, Yucatan Península, México. Mesoamerican Ecology Institute, Monograph 1. Middle American Research Institute, Publication 54, Tulane University, Nueva Orleans. 183 p. [ Links ]
45 Warmke, G. y R. Abbott. 1962. Caribbean seashells. Livingston, Narberth, Nueva Jersey. 384 p. [ Links ]
46 Warmke, G. L. y L. R. Almodóvar. 1963. Some associations of marine mollusks and algae in Puerto Rico. Malacología, 1: 163-177. [ Links ]
47 Wynne, M. J. 1998. A checklist of benthic marine algae of the tropical and subtropical western Atlantic: first revision. Nova Hedwigia, 116: 1-155. [ Links ]
FECHA DE RECEPCIÓN: 20/05/2011 FECHA DE ACEPTACIÓN: 02/01/2013
*Contribución No. 379 del Centro de Estudios en Ciencias del Mar (Cecimar), Universidad Nacional de Colombia sede Caribe.
