Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.42 no.2 Santa Marta July/Dec. 2013
INVERTEBRADOS MARINOS ASOCIADOS CON LAS PLATAFORMAS DE GAS EN LA GUAJIRA (CARIBE COLOMBIANO)*
DIVERSITY OF MARINE INVERTEBRATES ASSOCIATED TO GAS PLATFORMS IN LA GUAJIRA (COLOMBIAN CARIBBEAN SEA)
Adriana Gracia C.1, Norella Cruz2, Giomar Borrero1, Diana Patricia Báez3 y Nadiezhda Santodomingo4
1 Instituto de Investigaciones Marinas y Costeras (Invemar), Museo de Historia Natural Marina de Colombia. Calle 25 No. 2-55, Playa Salguero, Santa Marta, Colombia. adriana.gracia@invemar.org.co.
2 Investigadora independiente. Transversal 34 No. 40-39, Bogotá, Colombia. norecas@yahoo.es.
3 Investigadora independiente. Bogotá, Colombia. dpbaez@yahoo.com.
4 Natural History Museum, Department of Earth Sciences, SW7 5BD Londres, Reino Unido. n.santodomingo@nhm.ac.uk, nsantodomingo@gmail.com.
RESUMEN
El desarrollo industrial y económico de Colombia se ha expandido en la búsqueda de nuevos recursos minerales hacia los límites del mar territorial. La prospección y construcción de infraestructuras en el mar plantean nuevos retos para el desarrollo sostenible del país. Este estudio de línea base se realizó con el fin de conocer las especies de invertebrados marinos que se han establecido sobre las únicas plataformas permanentes para la explotación de gas ubicadas mar afuera en La Guajira, al norte del Caribe colombiano: plataformas A y B, construidas sobre fondos de 18 y 33 m de profundidad, respectivamente. Por medio de buceo autónomo Scuba, se recolectaron e identificaron 116 especies de invertebrados marinos. Los crustáceos presentaron la mayor riqueza de especies (50 especies), con el infraorden Brachyura como el mejor representado con 10 familias y acumulando 44% de las especies. Los moluscos comprendieron el segundo grupo con 29 especies, seguidos de anélidos con 14, esponjas con 12, cnidarios con 7 y finalmente equinodermos con 4. Los organismos sésiles filtradores dominaron en número de individuos, mientras las formas móviles presentaron un mayor número de especies. Los balanos (Balanus spp.), el octocoral Carijoa riisei y las esponjas fueron los principales constituyentes de la fauna fouling asociada a las estructuras. Aunque no existen estudios previos para el área de estudio ni para el Caribe sur, se considera que la comunidad de invertebrados marinos que ha colonizado las estructuras de las plataformas de gas de La Guajira exhibe una alta riqueza de especies e individuos asociados.
PALABRAS CLAVE: Sustratos artificiales, biofouling, macrofauna, Caribe colombiano, inventarios de especies.
ABSTRACT
The industrial and economic development of Colombia has expanded its limits towards the territorial sea in the search for new resources. The prospection and construction of infrastructures in the sea challenge new alternatives for the sustainable development of the country. The main goal of this study was to build a baseline inventory of the marine invertebrates associated to two gas platforms located offshore in La Guajira, at the northeastern Colombian Caribbean Sea: platforms A and B, sitting on sandy bottoms at 18 and 33 m depth, respectively. This inventory includes a total of 116 species of marine invertebrates that were collected by Scuba diving. Crustaceans had the greatest richness with 50 species, from which the infraorder Brachyura was the best represented with 10 families including approximately 44% of species. Mollusks comprised the second largest group with 29 species, followed by annelids with 14 species, sponges with 12 species, cnidarians with 7 species, and finally echinoderms with 4 species. Filter-feeding and sessile organisms dominated in abundance, while mobile species showed a larger number of species. The barnacles complex (Balanus spp.), the octocoral Carijoa riisei and sponges were the main components of the fouling fauna on the structures. Although there are no previous studies for the study area and the southern Caribbean, it is considered that the invertebrates community associated to gas platforms in La Guajira exhibit both high species richness and a large abundance of organisms.
KEY WORDS: Artificial substrates, biofouling, Colombian Caribbean, macrofauna, inventory of species.
INTRODUCCIÓN
El desarrollo económico costero de los países ha promovido la construcción de diferentes estructuras, tales como muelles, puertos, espolones, y plataformas de gas y petróleo, que funcionan a su vez como sustratos artificiales que brindan hábitat a una gran variedad de organismos marinos (Bomkamp et al., 2004). El término biofouling se refiere al proceso de colonización de organismos sobre una superficie sólida, bien sea viva o muerta (Wahl, 1989). Aunque para su traducción al español se ha sugerido el empleo de bioincrustación o incrustación biológica como equivalentes (Steel, 1994), en la práctica estos términos no han sido difundidos ampliamente, prevaleciendo el uso de biofouling en la mayoría de referencias.
El proceso de biofouling se presenta como resultado del asentamiento y crecimiento de organismos sedentarios sobre estructuras artificiales localizadas en ambientes marinos o estuarinos (Venugopalan y Wagh, 1990; Yan et al., 2009). Se ha sugerido que la secuencia de colonización o sucesión en sustratos artificiales es similar a la que se presenta en los arrecifes naturales (Bull et al., 1997). Sin embargo, sobre estructuras de mayor escala, como son las plataformas de gas o petróleo, la información biótica relacionada disponible es escasa, principalmente debido a su acceso restringido y rutinas de seguridad de las empresas operantes. Las plataformas localizadas en aguas someras proveen sustrato para una gran variedad de organismos de carácter arrecifal, los cuales a su vez, junto a los restos de material biológico que se desprenden de las plataformas (e.g. conchas de balanos y moluscos), proporcionan el hábitat adecuado para el asentamiento de muchas otras especies epibentónicas y endobentónicas (Sammarco et al., 2012; Gupta y Smith, 2013). Las plataformas se diferencian de los arrecifes artificiales porque los ejes que conforman su estructura se encuentran en contacto con toda la columna de agua desde la zona de salpicadura hasta el fondo, mientras que los arrecifes artificiales convencionales usualmente sólo ocupan entre 10 y 40% de la columna de agua, con un efecto en la riqueza y abundancia de los organismos que sustentan (Kolian, 2011). Por lo tanto, el suministro de información básica sobre la composición y estructura de las comunidades biofouling es importante para entender los procesos de sucesión que ocurren sobre sustratos duros localizados costa afuera.
Dentro de los antecedentes de estudios realizados en comunidades asociadas a plataformas de gas y petróleo, el golfo de México representa el ejemplo más conspicuo para el Atlántico occidental, en donde desde 1940 se han introducido al mar este tipo de estructuras (Sammarco et al., 2004), de las cuales se encuentran en operación aproximadamente unas 3600 plataformas (Atchison et al., 2008; Sammarco et al., 2012; Gupta y Smith, 2013). En este archipiélago de plataformas de gas y petróleo ubicadas mar afuera, se han realizado inventarios de las especies de invertebrados, con énfasis en la fauna comúnmente asociada a ambientes coralinos (Gallaway y Lewbel, 1982; Adams, 1996; Boland, 2002; Sammarco et al., 2004; Atchison et al., 2008). Mientras que para otras áreas del mundo como el mar del Norte, golfo de Arabia, golfo de China y Australia, parte de la literatura biológica asociada a estos lugares corresponde a comunidades ícticas y algunos de los invertebrados presentes (Stachowitsch et al., 2002; Bram et al., 2005; Neira, 2005; Gass y Roberts, 2006; Yan et al., 2006; Page et al., 2007). Este es el primer estudio que documenta la riqueza de invertebrados marinos asociados a las plataformas de gas de La Guajira en el Caribe colombiano y se enmarcó dentro del proyecto de investigación "Biodiversidad asociada a las plataformas de explotación de gas natural (La Guajira, Colombia)" (Santodomingo et al., 2004).
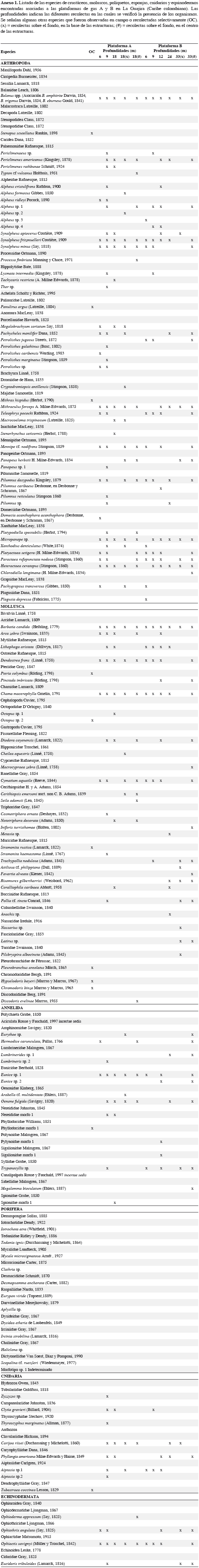
ÁREA DE ESTUDIO
Este estudio se realizó en las plataformas de gas conocidas con el nombre de Chuchupa, ubicadas en la parte media del departamento de La Guajira, al nororiente del Caribe colombiano. Para efectos de comparaciones, se denominaron plataformas A y B. La plataforma A se encuentra localizada en las coordenadas 11°47'N y 72°47'W, a 11 km de la costa y yace sobre un fondo arenoso de 18 m de profundidad. La plataforma B en las coordenadas 11°50'N y 72°50'W a 17 km de la costa y construida también sobre un fondo arenoso a 33 m de profundidad (Figura 1). Geomorfológicamente, el margen continental de este sector del Caribe colombiano está dominado por fondos arenosos con una baja pendiente y por lo tanto, es de poca profundidad sobre una gran extensión costa afuera. Aunque las descargas continentales no ejercen mucha influencia, las aguas suelen ser turbias por la continua resuspensión de sedimentos a causa del fuerte oleaje. Durante la primera época del año (diciembre-abril), este sector del Caribe colombiano es caracterizado por la presencia de surgencias (Andrade y Barton, 2005), que ocasionan un descenso de la temperatura, incremento en la salinidad y en el contenido de nutrientes de las masas de aguas superficiales (Díaz y Gómez, 2000).
MATERIALES Y MÉTODOS
La recolecta del material biológico se llevó a cabo entre octubre 28 y noviembre 5 de 2004. Las dos plataformas fueron muestreadas entre 6 y 33 m de profundidad; debido a las condiciones de oleaje fuerte no se realizó el muestreo en la zona intermareal (0-3 m). En la recolecta de organismos se empleó equipo de buceo autónomo Scuba, y los muestreos se realizaron en cada una de las estructuras a niveles distintos de profundidad: en la plataforma A a 6, 9 y 18 m, y en la plataforma B a 6, 9, 12, 24 y 33 m de profundidad. Para la obtención de las muestras se diseñó un cuadrante de 30 x 30 cm como área de referencia, en cuyo extremo se adaptó una bolsa de tela a manera de canasta de colecta. Con el cuadrante como referencia y utilizando la técnica de raspado con cincel y espátula, los organismos de la comunidad biofouling que se encontraban adheridos a las estructuras de las plataformas fueron removidos y recolectados dentro de las canastas (Figura 2). Con los mismos cuadrantes como referencia, se recolectaron muestras del fondo localizado inmediatamente debajo de las plataformas, el cual estaba compuesto principalmente por cascajo de balanos y otros organismos que, se presume, estuvieron adheridos directamente a las estructuras de las plataformas. Aunque al momento del muestreo no fue posible saber cómo y cuándo se realizó el último mantenimiento de estas estructuras, es sabido que el cuidado de estas plataformas de gas exige ejercicios de limpieza, en los cuales gran parte de la comunidad biofouling es removida para alivianar el peso y evitar el colapso de estas estructuras. El material recolectado se preservó en etanol al 70 y 96%, y se guardó en bolsas plásticas rotuladas, para su posterior separación e identificación. En adición a los cuadrantes, se hicieron recolectas selectivas de ejemplares asociados a las estructuras y se obtuvo un completo registro fotográfico en campo.

Una vez en el laboratorio, se realizó la separación por grupos mayores y cada uno fue identificado hasta el nivel taxonómico más bajo posible. Para la mayoría de las especies encontradas se obtuvieron fotografías en laboratorio empleando una cámara digital Nikon D1. El material biológico se encuentra depositado en las colecciones del Museo de Historia Natural Marina de Colombia (MHNMC), así como sus registros almacenados en el Sistema de Información sobre Biodiversidad Marina (SIBM) del Invemar y disponible en línea para su consulta en la dirección http://siam.invemar.org.co/siam/sibm/index.jsp.
RESULTADOS
En total se estudiaron seis grupos taxonómicos incluyendo esponjas, cnidarios, moluscos, crustáceos, poliquetos y equinodermos, correspondientes a más de 13321 ejemplares, de los cuales se identificaron un total de 116 especies. Los crustáceos presentaron la mayor riqueza con 50 especies, seguidos en orden descendente por moluscos con 29, anélidos con 14, esponjas con 12, cnidarios con 7 y finalmente equinodermos con 4. En el listado también se incluyeron algunos organismos que fueron observados en campo y algunas recolectas selectivas con el fin de completar el inventario de especies (esta información no se cuantifica en los resultados descritos para cada grupo). En general, se observó una alta colonización de organismos tanto incrustantes como móviles en las estructuras de las plataformas. Mientras que los organismos sésiles filtradores fueron dominantes en número de individuos, las formas móviles presentaron un mayor número de especies (Anexo 1).
Crustáceos
En el grupo de los crustáceos 40 especies fueron identificadas a nivel específico, y otras 10 lo fueron a nivel de género, con 9870 individuos recolectados. Estos organismos se agruparon dentro de las clases Maxillopoda y Malacostraca, cada una representada por un orden, Sessilia y Decapoda, respectivamente. La asociación de balanos perteneciente a la clase Maxillopoda, que incluye a las especies Balanus amphitrite, B. trigonus y B. eburneus, fue la más conspicua en número de ejemplares (88.7%), mientras que para los decápodos la especie cuantificada que presentó mayores valores de abundancia fue el camarón Synalpheus fritzmuelleri (3.2%). El orden Decapoda se encontró representado por tres de los seis infraordenes que lo componen: Caridea, Anomura y Brachyura. Aunque los decápodos no fueron muy abundantes y solo presentaron el 5.7% de los ejemplares recolectados, estuvieron representados por diez familias y constituyeron el 44% de las especies presentes. Del total de especies, 20 fueron exclusivas de la plataforma A, seis fueron exclusivas de la plataforma B y 23 fueron comunes a las dos estructuras (Anexo 1). La asociación de balanos (B. amphitrite, B. trigonus, B. eburneus), organismos que se alimentan de material en suspensión, estuvo presente en todo el gradiente batimétrico (6-33 m) y en las dos estructuras, de manera que fue el grupo sésil de organismos que dominó este hábitat artificial y dio sustento a la gran riqueza de invertebrados observados en las plataformas. Por ejemplo, los balanos vivos se observaron cubiertos principalmente por esponjas, mientras que en las conchas vacías se encontró una gran diversidad de organismos tanto sésiles como móviles que los utilizaban como refugio, tales como peces de la familia Blenniidae y crustáceos móviles. Synalpheus fritzmuelleri fue otra de las especies de crustáceos que estuvo presente en todo el gradiente batimétrico. A las especies recolectadas se sumaron las observaciones en campo del infraorden Palinura, con la langosta Panulirus argus, y por recolectas selectivas se encontraron especímenes del infraorden Stenopodidea como Stenopus scutellatus y Mithrax hispidus. En la Figura 3 se ilustran algunas de las especies recolectadas como Balanus spp. (a-b), S. fritzmuelleri (c), Petrolisthes galathinus (d), P. argus (e), Stenorynchus seticornis (f) y Mithrax hispidus (g).

Moluscos
El segundo grupo en número de especies correspondió a moluscos, con 24 especies identificadas y cinco a nivel de género (Anexo 1). De estas, 22 correspondieron a gastrópodos, seis a bivalvos y una a cefalópodos, con un total de 790 especímenes recolectados, de los cuales 77% correspondió a bivalvos, 22.9% a gastrópodos y 0.1% a cefalópodos. Con seis especies, la familia Muricidae fue la que presentó la mayor riqueza. Esta familia se encuentra caracterizada en su gran mayoría por ser de hábitos carnívoros, y algunas de sus especies se alimentan de pólipos de coral, lo cual podría relacionar su presencia con la especie dominante a mayores profundidades, el octocoral Carijoa riisei. Entre los moluscos, los bivalvos con los mayores números de individuos fueron Dendrostrea frons (47.6%), Chama macerophylla (14.9%) y Barbatia candida (9.9%), los tres reconocidos por ser organismos suspensívoros. Del total de especies, ocho solo se encontraron en la plataforma A, doce solo en la plataforma B y nueve fueron comunes a las dos estructuras. Se destacaron B. candida y C. macerophylla como las especies presentes en todo el gradiente batimétrico (6-33 m) y en las dos estructuras. Con relación a especies de hábitos herbívoros, Diodora cayennesis fue la única que se encontró en ambas estructuras, desde 9 hasta 33 m de profundidad y fue observada siempre adherida a áreas que presentaban algas coralináceas incrustantes. Adicionalmente, se observaron especies como Pleurobranchus areolatus, Hypselodoris bayeri y Chromodoris binza, entre la fauna que habita el cascajo bajo las plataformas (conformado principalmente por las conchas de balanos). En la Figura 4 se ilustran algunas especies recolectadas u observadas en campo: Pteria colymbus (a), Diodora cayenensis (b), Dendostrea frons (c), Chromodoris binza (d), Hypselodoris bayeri (e), Cymatium aquatile (f), Attiliosa cf. philippiana (g), Pollia cf. tincta (h), Stramonita haemastoma (i), Risomurex gilbertharrisi (j), Coralliophila caribaea (k), Pinctada imbricata (l), Barbatia candida (m), Lithophaga aristata (n), Chama macerophylla (o), Pleurobranchus areolatus (p) y Octopus sp. 2 (q).
Poliquetos
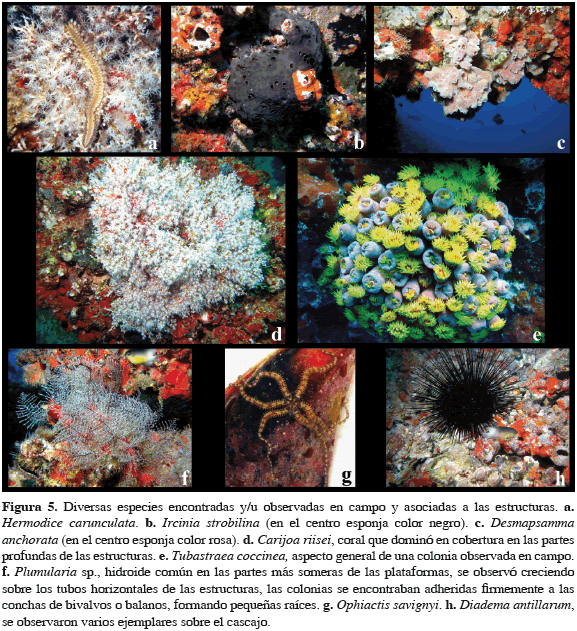
En lo que concierne a poliquetos se encontraron 14 especies de los órdenes Aciculata y Canalipalpata, con un total de 112 individuos asociados a las plataformas, cuatro fueron identificados a nivel de especie, seis a género y los restantes cuatro a familia. Los poliquetos que concentraron el mayor número de individuos para las dos plataformas fueron las especies Oenone fulgida (34.8%), Eunice sp. (19.6%) y Trypanosyllis sp. (17.9%). De las 14 especies, cuatro fueron exclusivas de la plataforma A, cinco exclusivas de la B y cinco fueron comunes a las dos estructuras (Anexo 1). Eunice sp. fue común en las dos plataformas y estuvo presente en todo el gradiente batimétrico, mientras que una de las especies visualmente más conspicua en campo fue Hermodice carunculata (Figura 5a). En general, los poliquetos asociados a las plataformas presentaron gran variedad de formas de vida y hábitos alimentarios, e.g. de vida libre y sedentarios; omnívoros, carnívoros y detritívoros, entre otros.
Esponjas
Entre las esponjas se identificaron doce especies pertenecientes a once familias y cuatro órdenes, ocho fueron identificadas a nivel de especie, tres a género y una a morfotipo (Anexo 1), todas ellas de hábitos filtradores. Estas se encontraron creciendo directamente sobre la estructura o colonizando otros organismos, construyendo tapetes de pocos milímetros de espesor (e.g. Aplysilla sp. y Mycale microsigmatosa), colonias en forma de tubos (e.g. Clathria sp. y Desmapsamma anchorata) y formas masivas (e.g. Ircinia strobilina). Aunque la plasticidad ecofenotípica de las esponjas hace que se dificulte su identificación en campo, se observó que las especies con mayores coberturas fueron M. microsigmatosa y Tedania ignis, las cuales proporcionaron a los soportes de las estructuras y a la comunidad asociada un aspecto general de color anaranjado a rojo. En la Figura 5 se ilustran las esponjas I. strobilina (b) y D. anchorata (c).
Cnidarios
Los cnidarios asociados directamente a las estructuras comprendieron siete especies, identificadas cuatro a nivel de especie y tres a género (Anexo 1). De las siete especies, dos se presentaron únicamente en la plataforma A, mientras que las demás fueron comunes a las dos estructuras. La caracterización de los cnidarios (hidroides, corales y anémonas) se hizo de manera cualitativa principalmente a través de recolectas manuales ya que, con excepción del octocoral C. riisei (Figura 5d) no representaron un elemento conspicuo en la comunidad adherida a las plataformas. No obstante, se cuantificaron aquellos organismos que se encontraron comúnmente entre las muestras recolectadas con los cuadrantes, como el coral duro Phyllangia americana, las anémonas Aiptasia sp. 1 y Aiptasia sp. 2, y el hidroide Zyzzyzus sp., de las cuales Aiptasia sp. 1 acumuló 88.9% del número de ejemplares de cnidarios. Entre los corales escleractinios, solo fueron observadas colonias de los azooxantelados P. americana y Tubastraea coccinea (Figura 5e). Phyllangia americana se encontró creciendo principalmente dentro de los tubos de balanos vacíos y en algunos casos formando pequeñas colonias de varios coralites adheridas directamente a las columnas de las plataformas, en la base de C. riisei o sobre conchas de balanos. En cuanto a la fauna de octocorales es de destacar a C. riisei, el cual está presente desde 9 m de profundidad, pero fue dominante a partir de 10 a 12 m de profundidad y hasta el fondo. La mayor parte de las colonias que crecían sobre las columnas de las plataformas se encontraron asentadas sobre pequeños balanos y, a su vez, la mayoría de las colonias (> 90%) se encontraron recubiertas de esponjas que se adhieren principalmente sobre los ejes. Sobre el cascajo bajo las estructuras, su presencia se restringió a pequeñas colonias y en las recolectas solo se encontraron fragmentos de ellas. En la Figura 5f adicionalmente se ilustra al hidroide Plumularia sp.
Equinodermos
Entre los equinodermos estudiados se identificaron cuatro especies asociadas directamente a las estructuras (Anexo 1), con un total de 1807 individuos; en ambas plataformas el ofiuro Ophiactis savignyi (Figura 5g) se observó en todo el gradiente batimétrico con el 99% del número de ejemplares. Ophioderma appressum solo se presentó en la plataforma A y las tres especies restantes fueron comunes a las dos estructuras. Sobre el fondo de las estructuras se observó al erizo Diadema antillarum (Figura 5h).
DISCUSIÓN
Aunque no se tienen antecedentes de este tipo de estudios en el Caribe colombiano o lugares cercanos en el Caribe sur, las 116 especies de invertebrados marinos asociados a las plataformas de gas de Chuchupa podrían considerarse como un alto valor en riqueza de especies, si se compara con inventarios sobre otros sustratos artificiales (e.g. hábitats y arrecifes artificiales) realizados en la bahía de Taganga y el golfo de Morrosquillo (Delgadillo-Garzón, 2009). En el primer lugar se registra la presencia de nueve especies de macroinvertebrados móviles que incluyen a los equinodermos Astropyga magnifica, Diadema antillarum e Isostichopus badionotus, y a los crustáceos Dromia erythropus, Lysmata grabhami, Mithrax sp., Panulirus argus, Stenopus hispidus y Stenorhynchus seticornis (Delgadillo-Garzón, 2009). Mientras que para el golfo de Morrosquillo se han encontrado hasta 19 especies de macroinvertebrados móviles, que incluyen un cnidario, un ctenóforo, un molusco, un poliqueto, 11 crustáceos y 4 equinodermos, adicionalmente 128 especies de organismos sésiles y epifaunales (2 cianofitas, 3 clorófitas, 2 rodófitas, 20 poríferos, 15 cnidarios, 28 moluscos, 23 anélidos, 6 sipuncúlidos, 18 crustáceos, 2 equinodermos, 8 ectoproctos y un cordado) (Delgadillo-Garzón et al., 2010).
En comparación con los estudios del golfo de México, la riqueza de especies de invertebrados marinos de las plataformas de gas de La Guajira es mayor, ya que allí solo se han registrado unas 31 especies entre invertebrados incrustantes y móviles (Carney, 2005). En plataformas de hidrocarburos localizadas fuera de zonas tropicales, como en el mar Adriático, el número de especies es menor con cerca de 40 (Relini et al., 1998), mientras que en plataformas del golfo de China se han registrado hasta 105 taxones distintos en la comunidad biofouling (Yan et al., 2006), incluyendo a otros grupos que no fueron tenidos en cuenta en el presente estudio, como briozoos y algas. En el golfo de China, a 38 m de profundidad, la comunidad biofouling está compuesta principalmente de especies litorales de ambientes tropicales y subtropicales, de los cuales los moluscos están representados por 41 especies, artrópodos 21, anélidos 14, cnidarios 9, briozoos 9, algas 7 y equinodermos 2, mientras que poríferos, turbelarios, braquiópodos y sipuncúlidos por una sola respectivamente (Yan et al., 2006). No obstante, es importante tener en cuenta las diferencias en localidad (latitud), distancia a la costa, profundidad y tamaño de las estructuras, complejidad de los microhábitats y tiempos en los procesos de colonización, que pueden no hacer comparables dichos valores.
En las plataformas de gas de Chuchupa, la comunidad estuvo dominada por balanos y bivalvos, al igual que en las plataformas de hidrocarburos del golfo de México, donde se ha documentado que las principales especies que dominan estos hábitats artificiales, entre 1 y 30 m de profundidad, corresponden a estos grupos y representan más del 99% de la biomasa (Lewbel et al., 1987; Carney, 2005). Otros grupos abundantes en las plataformas del golfo de México incluyen ofiuros, anémonas, zoantideos, caprélidos, anfípodos gamáridos, hidroides, briozoos, tunicados y esponjas cliónidas (Lewbel et al., 1987). En el mar Adriático los bivalvos y poliquetos sedentarios fueron los taxones más representativos, seguidos por los balanos (Relini et al., 1998).
Se debe tener en cuenta que, aunque muchas de las especies que se encuentran sobre las plataformas son registradas en hábitats rocosos locales submareales e intermareales, el ambiente físico y biológico de las plataformas costa afuera difiere al encontrado en dichos hábitats (Bram et al., 2005), por lo que sería un aspecto interesante de estudio y comparación para detectar tasas de crecimiento, presión por pastoreo y efectos naturales, entre otros, que podrían explicar mejor dichos procesos de sucesión en medios naturales y artificiales. Por observaciones de los autores en campo se evidenció la presencia de organismos con grandes tallas (e.g. Pteria colymbus), lo que podría deberse en primera medida a que este es un lugar de limitado acceso, con baja perturbación antropogénica y, de otra parte, podría estar relacionado con el efecto de la surgencia que se presenta en esta región y que podría manifestarse en mayores tallas de crecimiento relacionadas con una mayor disponibilidad de alimento.
Los balanos en general son característicos de la fauna fouling en cascos de barcos, pilotes de muelles, boyas, raíces de manglar y litoral rocoso, entre otros, por lo que las estructuras estudiadas constituyen un sustrato ideal para el asentamiento de este tipo de organismos. Se ha establecido que Cirripedia (balanos) son los invertebrados sésiles que dominan (58-65% de la biomasa húmeda) en estructuras ubicadas en aguas menores a 30 m de profundidad (Topolski y Szedlmayer, 2004). Por otra parte, Synalpheus fritzmuelleri, la especie de crustáceo móvil más abundante, es bastante común en una gran diversidad de hábitats, como fragmentos de coral, entre raíces de manglar, praderas de pastos con corales y cavidades de esponjas (Román-Contreras y Martínez-Mayén, 2010). En el occidente del golfo de México, esta especie detritívora es uno de los mayores componentes de la epifauna de crustáceos habitante de arrecifes sublitorales someros (Felder, 1982; Gutiérrez-Salcedo et al., 2007) y ha sido registrada igualmente en plataformas de gas (Gallaway y Lewbel, 1982). En general, la familia Alpheidae es uno de los taxa con mayor riqueza de especies (10); este es un grupo de organismos abundantes y diversos ecológicamente en la mayoría de las aguas marinas tropicales y subtropicales, también en aguas frías-templadas, agua dulce y hábitats de aguas salobres, como los manglares y estuarios (Román-Contreras y Martínez-Mayén, 2010), por lo que su presencia y alta diversidad en este estudio podría ser esperada.
En el caso de los moluscos, al comparar los resultados con estudios sobre diversidad en plataformas, se ha encontrado un número similar de especies habitando este de tipo de sustratos. Anderson (2009) en las plataformas de Luisiana encontró 27 especies de bivalvos y gastrópodos, con Isognomon bicolor y B. candida formando cerca del 90% del ensamblaje total. En general, las especies de gastrópodos encontradas en las plataformas de gas estudiadas son, en su mayoría, típicas habitantes de substratos duros o rocosos, pero es muy poco lo que se conoce en cuanto a su presencia en este tipo de estructuras en el mundo, ya que posiblemente no han sido estudiadas en detalle debido a que la mayoría de investigaciones se han concentrado en biotecnología de organismos que podrían servir como fuente de productos farmacéuticos y de los cuales los bivalvos presentan gran interés.
El poliqueto Oenone fulgida, una de las especies más representativas de este grupo en las plataformas de La Guajira, es una especie circumtropical que se encuentra asociada con arrecifes coralinos, donde ha sido registrado como perforador de coral en el Caribe (Fauchald y Jumars, 1979). Igualmente, O. fulgida ha sido encontrado en fondos lodosos, arenosos y de algas, desde el intermareal hasta 40 m de profundidad (Dean, 2004), revelando su alta plasticidad para colonizar una gran variedad de hábitats y, por lo tanto, uno de los poliquetos más conspicuos en este estudio. El género Eunice, por su parte, ha encontrado entre los balanos un sustrato alternativo a los que comúnmente se han registrado, ya que en condiciones naturales estos poliquetos habitan tanto fondos blandos como duros, muchos de ellos horadando corales duros, algas calcáreas o viviendo entre sus cavidades (Fauchald, 1992; Zanol et al., 2007). De las familias Lumbrineridae y Phyllodocidae, los ejemplares se encontraron principalmente entre balanos muertos, conchas y esponjas.
Las esponjas, uno de los grupos visualmente más conspicuos en las plataformas estudiadas, podrían ser consideradas como el grupo más exitoso en la competencia por el limitado recurso de sustrato disponible, ya que se observaron viviendo directamente sobre la estructura, o colonizando a otros organismos. Uno de los roles que desempeñan en este ecosistema artificial es el de ofrecer entre las cavidades de su propia estructura un microhábitat que sirve de refugio para un gran número de ofiuros, camarones y cangrejos. En algunas ocasiones se observaron especies que se mimetizaban con las esponjas ya fuera a través del color (e.g. cangrejo Paractaea rufopunctata nodosa), o por recubrimiento con las mismas (cangrejos Macrocoeloma trispinosum y Cryptodromiopsis antillensis). Es importante mencionar que un estudio minucioso de la composición de especies y abundancias relativas de esponjas a lo largo del gradiente de profundidad en las dos plataformas, aportaría elementos claves para el entendimiento de la dinámica de este tipo de ecosistemas y permitiría explicar con mayor evidencia el papel que juegan como refugio de pequeños invertebrados, así como en los primeros eslabones de la cadena trófica de este tipo de comunidades fouling.
La especie más conspicua de cnidarios fue el octocoral C. riisei, la cual es común en el Caribe (Bayer, 1961) y que se encuentra creciendo comúnmente en pilotes de muchos puertos y también aparece a menudo en gran abundancia en las plataformas costa afuera (Bull et al., 1997). En Colombia había sido registrada para La Guajira mediante los proyectos Macrofauna (Invemar, 2002) o asociada a pilotes de muelles en la bahía de Cartagena (Sánchez, 1994). En el resto de especies de corales se observó que las colonias fueron de bajo porte y se confundían fácilmente con algas filamentosas, a diferencia de lo registrado en algunas áreas del golfo de México, donde ha sido posible el establecimiento de corales escleractínios formadores de arrecife, tales como los corales cerebro Diploria spp. o ramificados de los géneros Porites y Madracis (Sammarco, 2003; Sammarco et al., 2004). A pesar de que T. coccinea es un coral bastante frecuente en las zonas someras de acantilados rocosos y arrecifes del Caribe (Cairns, 2000), de esta especie de coral solo se observaron dos colonias localizadas en la parte sombreada de los tubos horizontales de la plataforma B, de modo que su presencia no fue conspicua. Esta baja representatividad de corales duros pudo ser debida a que el tiempo transcurrido desde que fueron instaladas las plataformas (aproximadamente 20 años para la plataforma A y siete para la B) hasta el momento del muestreo, no fue suficiente para que las condiciones ambientales y características específicas del microhábitat (complejidad estructural) favorecieran el desarrollo coralino. Cabe anotar que, durante la salida de reconocimiento (agosto 5 de 2004) se observó una pequeña colonia del género Siderastrea (aprox. 6 cm de diámetro) creciendo sobre una roca en el fondo de la plataforma, lo cual deja abierta la posibilidad de que eventualmente y a largo plazo podría establecerse una comunidad de corales hermatípicos (que construyen arrecife) tal y como ha sido observado en el golfo de México (Sammarco, 2003; Sammarco et al., 2004). Sería interesante monitorear el ecosistema con el fin de estudiar la trayectoria de los cambios relacionados con la composición de especies y de esta manera detectar el momento, en el mejor de los casos, en que los corales arrecifales inicien eventualmente la colonización del área.
Como parte del proceso de sucesión que se desarrolla sobre estructuras artificiales introducidas en el mar, los equinodermos son uno de los grupos comunes entre las especies móviles que se asientan en las últimas etapas; sin embargo, es escasa la información específica sobre este grupo y su asociación con plataformas de producción y estructuras artificiales en general. La especie de equinodermos más común en las plataformas estudiadas fue el ofiuro Ophiactis savignyi (Figura 5g), el cual está comúnmente asociado con ecosistemas como arrecifes de coral y fondos de algas marinas (Hendler et al., 1995); también ha sido encontrado en comunidades fouling de barcos y otras estructuras artificiales oceánicas, llegando a vivir dentro de esponjas con densidades de hasta 3000 individuos por litro (Roy y Sponer, 2002). Ophiactis savignyi tiene la capacidad de reproducirse sexual y asexualmente, lo que podría explicar su abundancia y amplia distribución batimétrica en las plataformas estudiadas; ha sido definida como detritófaga, alimentándose de granos de arena, briozoos, foraminíferos y pequeños gastrópodos o también como que usa material en suspensión, lo cual le da ventajas en una comunidad como la que se desarrolla en estas plataformas, compuesta por organismos de los grupos mencionados y cuyos hábitos alimentarios se basan en material en suspensión (Stachowitsch et al., 2002).
De las especies encontradas en las estructuras se resalta la presencia de dos de ellas que se encuentran en los libros rojos de fauna amenazada de Colombia (Ardila et al., 2002). Estas son la langosta espinosa P. argus (categoría "vulnerable"), la cual por su interés comercial ha sido sobreexplotada a lo largo de la costa del Caribe colombiano, y el erizo negro D. antillarum (categoría "datos insuficientes"), especie que presentó una mortandad masiva en el Caribe en 1983. Aunque estas especies fueron observadas por los autores habitando tanto sobre las estructuras de las plataformas, como entre el cascajo al fondo de las mismas, su presencia es un buen indicador de que sustratos artificiales con una baja presión de explotación humana pueden ser fuente de refugio y/o reproducción para algunas especies que actualmente se encuentran diezmadas o sobreexplotadas en hábitats naturales.
CONCLUSIONES
En términos generales, la composición por grupos taxonómicos es similar a la que ha sido registrada para otro tipo de sustratos duros como raíces de manglar, litoral rocoso, arrecifes de coral y otras estructuras artificiales como pilotes de muelles y plataformas de gas y petróleo del Atlántico occidental, siendo las especies sésiles considerablemente más abundantes en número de individuos que las móviles. Comparativamente con los otros grupos de invertebrados presentes en este estudio, los crustáceos y moluscos se destacaron por ser los más abundantes en número de individuos y diversos en número de especies.
Estas plataformas funcionan como hábitat para una gran variedad de invertebrados sésiles y móviles, que son capaces de colonizar estas estructuras artificiales a través del desarrollo de múltiples estrategias de crecimiento. Dicha comunidad de invertebrados ha creado un sustrato con características tridimensionales tales, que puede ser utilizado a su vez por otros organismos, como la gran cantidad de peces que son atraídos hacia las estructuras (Santodomingo et al., 2004). Es importante considerar que actualmente Colombia se encuentra en una fase de intensa exploración de recursos minerales costa afuera, que de resultar positiva vislumbraría un nuevo panorama hacía la construcción de plataformas mar afuera. Dicha intervención en el ambiente oceánico traería consigo no solo cambios en las comunidades marinas al ofertar nuevos espacios para ser colonizados por numerosas especies de invertebrados marinos, como las registradas en este estudio, sino que también abriría el camino a futuras investigaciones en este campo abordadas desde diferentes disciplinas de la biología como la biodiversidad, ciclos de vida, desarrollo, conectividad y bioprospección, entre otras.
AGRADECIMIENTOS
Se agradece a la Empresa Colombiana de Petróleos Ecopetrol S. A. e Invemar por el apoyo logístico y financiero de esta investigación (proyecto No. GEA-C-RCC-3-236-04). A P. Restrepo, C. Guerrero y R. Hernández por su apoyo incondicional a esta iniciativa. A todas aquellas personas que participaron en la colecta del material biológico y en la salida de campo, especialmente a los biólogos J. Reyes, A. Rodríguez, M. López, C. Reyes y J. C. Márquez quienes además ayudaron en algunas de las fotografías en campo. También a los operarios J. Sánchez y E. Vilarete del Invemar. Sven Zea asistió en la identificación de las esponjas y F. Cortes colaboró con el componente bentónico. A N. Rangel por su apoyo en la elaboración del mapa.
BIBLIOGRAFÍA
1 Adams, C. L. 1996. Species composition, abundance and depth zonation of sponges (phylum Porifera) on an outer continental shelf gas production platform, northwestern Gulf of Mexico. Reporte final y tesis MSc, Texas A and M University-Corpus Christi, Center for Coastal Studies, Corpus Christi, EE. UU. 260 p. [ Links ]
2 Anderson, L. C. 2009. Mollusca from oil and gas platform on the Louisiana Continental shelf: A biotechnology survey. 37-42. En: Rouse, L. (Ed.). Evaluation of oil and gas platforms on the Louisiana continental shelf for organisms with biotechnology potential. U.S. Department of the Interior, Minerals Management Service, Nueva Orleans, EE. UU. 53 p. [ Links ]
3 Andrade, C. A. y E. D. Barton. 2005. The Guajira upwelling system. Cont. Shelf Res., 25: 1003-1022. [ Links ]
4 Ardila, N., G. R. Navas y J. Reyes (Eds.). 2002. Libro rojo de invertebrados marinos de Colombia. La serie Libros Rojos de Especies Amenazadas de Colombia, Invemar y Ministerio de Medio Ambiente, Bogotá. 180 p. [ Links ]
5 Atchison, A. D., P. D. Sammarco y D. A. Brazeau. 2008. Genetic connectivity in corals on the Flower Garden banks and surrounding oil/gas platforms, Gulf of Mexico. J. Exp. Mar. Biol. Ecol., 365: 1-12. [ Links ]
6 Bayer, F. 1961. The shallow water Octocorallia of the West Indian Region. A manual of marine biologist. Stud. Fauna Curacao, 12: 1-373. [ Links ]
7 Boland, G. S. 2002. Fish and epifaunal community observations at an artificial reef near a natural coral reef: nineteen years at High Island platform A-389-A, from bare steel to coral habitat. 372-392. En: McKay, M., J. Nides y D. Vigil (Eds.). Proc. Gulf Mex. Fish and Fisheries: bringing together new and recent research, October 2000. U.S. Department of the Interior, Nueva Orleans, EE. UU. 674 p. [ Links ]
8 Bomkamp, R. E., H. M. Page y J. E. Dugan. 2004. Role of food subsidies and habitat structure in influencing benthic communities of shell mounds at sites of existing and former offshore oil platforms. Mar. Biol., 146: 201-211. [ Links ]
9 Bram, J. B., H. M. Page y J. E. Dugan. 2005. Spatial and temporal variability in early successional patterns of an invertebrate assemblage at an offshore oil platform. J. Exp. Mar. Biol. Ecol., 317: 223-237. [ Links ]
10 Bull, A., L. Dauterive, D. Fertl, G. Goeke, J. Kendall, C. Langley y R. Regio. 1997. "Islands of life" A teacher's companion! Minerals Management Service, Nueva Orleans, EE. UU. 23 p. [ Links ]
11 Cairns, S. 2000. A revision of the shallow-water azooxanthellate Scleractinia of Western Atlantic. Stud. Nat. Hist. Car. Reg., 75: 1-231. [ Links ]
12 Carney, R. S. 2005. Characterization of algal-invertebrate mats at offshore platforms and the assessment of methods for artificial substrate studies: Reporte final. U.S. Department of the Interior, Minerals Management Service, Gulf of Mexico OCS, Nueva Orleans, EE. UU. 93 p. [ Links ]
13 Dean, H. K. 2004. Marine biodiversity of Costa Rica: Class Polychaeta (Annelida). Rev. Biol. Trop., 52 (Suppl. 2): 131-181. [ Links ]
14 Delgadillo-Garzón, O. 2009. Peces y macroinvertebrados móviles de hábitats artificiales en la bahía de Taganga, Caribe colombiano. Bol. Invest. Mar. Cost., 38 (1): 197-204. [ Links ]
15 Delgadillo-Garzón, O., F. Delgado-Cuadros, R. M. Henríquez, L. M. García, T. Ruiz-Álvarez, A. Zamora-Guzmán, K. Martínez, F. Blanco, G. Coronado-Luna, M. J. Mogollón, F. Puerta-Padilla, J. C. Jiménez, L. Navarro, I. M. J. Vishnoff y S. N. Perdomo-Ramos. 2010. Avances y desafíos del programa Diáspora: Arrecifes artificiales en el golfo de Morrosquillo, Caribe colombiano. 561-565. En: Invemar-Acimar (Eds.). Libro de resúmenes extendidos XIV Seminario Nacional de Ciencias y Tecnologías del Mar (Senalmar). Serie de Publicaciones Especiales de Invemar No. 21, Santa Marta. 578 p. [ Links ]
16 Díaz, J. M. y D. I. Gómez (Eds.). 2000. Programa nacional de investigación en biodiversidad marina y costera (PNIBM). Invemar, Fonade y MMA. Santa Marta. 83 p. [ Links ]
17 Fauchald, K. 1992. A review of the genus Eunice (Polychaeta: Eunicidae) based upon type material. Smith. Contr. Zool., 523: 1-422. [ Links ]
18 Fauchald, K. y P. Jumars, 1979. The diet of worms: a study of polychaete feeding guilds. Oceanogr. Mar. Biol. Ann. Rev., 17: 193-284. [ Links ]
19 Felder, D. L. 1982. Reproduction of the snapping shrimps Synalpheus fritzmuelleri and S. apioceros (Crustacea: Decapoda: Alpheidae) on a sublittoral reef off Texas. J. Crust. Biol., 2 (4): 535-543. [ Links ]
20 Gallaway, B. I. y G. S. Lewbel. 1982. The ecology of petroleum platforms in the northwestern Gulf of Mexico: a community profile. U.S. Fish and Wildlife Service, Office of Biological Services, FWS/OBS-82/27. Bureau of Land Management, Gulf of Mexico OCS Regional Office, Open-File Report 82-03. Washington. 92 p. [ Links ]
21 Gass, S. E. y J. M. Roberts. 2006. The occurrence of the cold-water coral Lophelia pertusa (Scleractinia) on oil and gas platforms in the North Sea: Colony growth, recruitment and environmental controls on distribution. Mar. Poll. B., 52: 549-559. [ Links ]
22 Gupta, B. K. y L. E. Smith. 2013. Foraminifera of petroleum platforms, Louisiana shelf, Gulf of Mexico. Mar. Micropaleontol., 101: 161-179. [ Links ]
23 Gutiérrez-Salcedo, J. M., M. I. Aguilar-Pérez, A. Bermúdez, N. H. Campos y G. Navas. 2007. Estructura de la macrofauna de invertebrados presente en los nidos del pez Malacanthus plumieri (Bloch, 1786) (Perciformes: Malacanthidae) en la bahía de Nenguange, Parque Nacional Natural Tayrona, mar Caribe colombiano. Caldasia, 29 (2): 309-328. [ Links ]
24 Hendler, G., J. E. Miller, D. L. Pawson y P. M. Kier. 1995. Sea stars, sea urchins and allies. Echinoderms of Florida and the Caribbean. Smithsonian Institution Press. Washington. 390 p. [ Links ]
25 Invemar. 2002. Caracterización de la macrofauna de la plataforma continental del mar Caribe colombiano y estado del conocimiento de la macrofauna de la plataforma continental del océano Pacífico colombiano y transferencia tecnológica y de información al SINA (UAESPNN, Corpoguajira y CVC). Informe final proyecto, Invemar, Santa Marta. 176 p. [ Links ]
26 Kolian, S. 2011. The benefits of leaving oil and gas rigs intact to serve as artificial reefs. Exploration and Production. Oil and Gas Review, 9 (2): 59-62. [ Links ]
27 Lewbel, G. S., R. L. Howard y B. J. Gallaway. 1987. Zonation of dominant fouling organisms on northern Gulf of Mexico petroleum platforms. Mar. Environ. Res., 21 (3): 199-224. [ Links ]
28 Neira, F. J. 2005. Summer and winter plankton fish assemblages around offshore oil and gas platforms in south-eastern Australia. Est. Coast. Shelf. Sci., 63: 589-604. [ Links ]
29 Page, H. M., Dugan, J. E., Scroeder, D. M., Nishimoto, M. M., Love, M. S. y J. C. Hoesterey. 2007. Trophic links and condition of a temperate reef fish: comparisons among offshore oil platform and natural reef habitats. Mar. Ecol. Prog. Ser., 344: 245-256. [ Links ]
30 Relini, G., F. Tixi, M. Relini y G. Torchia. 1998. The macrofouling on offshore platforms at Ravenna. Int. Bio. Bio., 41: 41-55. [ Links ]
31 Román-Contreras, R. y M. Martínez-Mayén. 2010. Notes on some alpheid shrimps (Decapoda: Caridea) of Thalassia testudinum meadows, from the Central-Southern Mexican Caribbean. Hidrobiológica, 20 (3): 222-229. [ Links ]
32 Roy, M. S. y R. Sponer. 2002. Evidence of a human-mediated invasion of the tropical western Atlantic by the 'world's most common brittlestar. Proc. R. Soc. Lond. B, 269: 1017-1023. [ Links ]
33 Sammarco, P. W. 2003. Coral reef communities on oil and gas platforms in the Gulf of Mexico: Where nature and industry meet. Presentation to House Sub-Committee on Energy and Natural Resources. Washington, D. C. http://republicans.resourcescommittee.house.gov/UploadedFiles/sammarco_9.17.03.pdf. 16/12/2009. [ Links ]
34 Sammarco, P. W., A. D. Atchison y G. S. Boland. 2004. Expansion of coral communities within the Northern Gulf of Mexico via offshore oil and gas platform. Mar. Ecol. Prog. Ser., 280: 129-143. [ Links ]
35 Sammarco, P. W., A. D. Atchison, G. S. Boland, J. Sinclair y A. Lirette. 2012. Geographic expansion of hermatypic and ahermatypic corals in the Gulf of Mexico, and implications for dispersal and recruitment. J. Exp. Mar. Biol. Ecol., 436-437: 36-49. [ Links ]
36 Sánchez, J. A. 1994. Presencia de los octocorales Stylatula diadema Bayer (Pennatulacea) y Carijoa riisei (Duchassaing y Mechelotti) (Telestacea) en la costa Caribe colombiana. An. Inst. Invest. Mar. Punta Betín, 23: 137-147. [ Links ]
37 Santodomingo, N., A. Gracia, A. Rodríguez, C. Reyes, J. Reyes-Forero, J. C. Márquez, N. Cruz-Castaño, P. Flórez, D. P. Báez y F. Cortes. 2004. Biodiversidad asociada a las plataformas de gas natural Campo Chuchupa: La Guajira, Colombia. Informe final proyecto, Invemar-Ecopetrol, Santa Marta. 225 p. [ Links ]
38 Stachowitsch, M., R., Kikinger, J., Herler, P., Zolda y E. Geutebruck. 2002. Offshore oil platforms and fouling communities in the southern Arabian Gulf (Abu Dhabi). Mar. Poll. B., 44: 853-860. [ Links ]
39 Steel, B. 1994. Glosario de ecología y afines: inglés-español. Ediciones Istmo S. A., Madrid. 447 p. [ Links ]
40 Topolski, M. F. y S. T. Szedlmayer. 2004. Vertical distribution, size structure, and habitat associations of four Blenniidae species on gas platforms in the northcentral Gulf of Mexico. Env. Biol. F., 70: 193-201. [ Links ]
41 Venugopalan, V. P. y A. B. Wagh. 1990. Biofouling of an offshore oil platform: Faunal composition and biomass. Indian J. Mar. Sci., 19: 53-56. [ Links ]
42 Wahl, M. 1989. Marine epibiosis. I. Fouling and antifouling: some basic aspects. Mar. Ecol. Prog. Ser., 58 (1): 175-189. [ Links ]
43 Yan, T., W. Yan, Y. Dong, H. Wang, Y. Yan y G. Liang. 2006. Marine fouling of offshore installations in the northern Beibu Gulf of China. Int. Biodeterior. Biodegrad., 58: 99-105. [ Links ]
44 Zanol, J., Fauchald, K. y P. Paiva. 2007. A phylogenetic analysis of the genus Eunice (Eunicidae, Polychaete, Annelida). Zool. J. Linn. Soc-Lon., 150: 413-434. [ Links ]
FECHA DE RECEPCIÓN: 11/01/2012 FECHA DE ACEPTACIÓN: 11/09/2013
*Contribución No. 1134 del Instituto de Investigaciones Marinas y Costeras (Invemar).