










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.43 no.1 Santa Marta Jan./June 2014
VARIACIÓN ESPACIO-TEMPORAL DE LA BIOMASA DEL ALGA CAULERPA SERTULARIOIDES EN POBLACIONES NATURALES Y EN CONDICIONES DE CULTIVO EN ESTANQUES DE CAMARÓN EN LA BAHÍA DE TUMACO, PACÍFICO COLOMBIANO
SEASONAL AND SPATIAL VARIATIONS OF CAULERPA SERTULARIOIDES FROM NATURAL POPULATIONS AND UNDER CULTURE CONDITIONS IN SHRIMP PONDS IN TUMACO BAY, COLOMBIAN PACIFIC
Hernel Marín-Salgado y Enrique Javier Peña-Salamanca
Universidad del Valle, Departamento de Biología, Calle 13 No 100-00 Cali, Colombia. A. A. 25360. hernelmarin@gmail.com, enrique.pena@correounivalle.edu.co.
RESUMEN
El presente estudio evaluó las variaciones espacio-temporales de la biomasa del alga verde Caulerpa sertularioides (Chlorophyta, Bryopsidales) en poblaciones naturales y en condiciones de confinamiento en estanques de camarón durante marzo-diciembre de 2010 y enero-junio de 2011. La biomasa de las poblaciones naturales se colectó a nivel submareal con cuadrante de 625 cm2. Simultáneamente se tomaron variables de calidad de agua (oxígeno disuelto, salinidad, temperatura, transparencia, NO3-, NH3 y PO43-). Las algas colectadas se llevaron al laboratorio para eliminar residuos. Posteriormente se obtuvo el peso fresco y seco para obtener la biomasa final. Las variables fisicoquímicas se evaluaron semanalmente. De igual manera se evaluó el crecimiento y la biomasa de plantas aclimatadas en piscinas para el cultivo de camarón, obteniendo una tasa de crecimiento relativo (TCR % d-1) promedio de 1.69% dia-1 (± 0.97). En los esteros Natal y Pajal no se registraron diferencias significativas (p = 0.315) en cuanto a biomasa de C. sertularioides, presentándose promedios entre 4.09 y 4.67 g cm-2. A nivel temporal, se encontró una diferencia significativa de la biomasa algal entre los meses de muestreo (p = 0.000039), con los mayores valores registrados en los meses de julio a septiembre. En los registros de salinidad para el estero Natal se presenta una relación directa con la variación temporal de biomasa y salinidad (p = 0.019) registrándose los mayores valores de biomasa en los meses en que se presentan salinidades altas en el área de estudio (julio-septiembre). En contraste, en el estero Pajal no se encontró relación entre biomasa y salinidad (p = 0.97). Los nitratos, nitrógeno amoniacal y ortofosfatos no fueron significativos con respecto a la variación de biomasa de la especie (p = 0.93, p = 0.33, p = 0.55, respectivamente). Se encontraron diferencias significativas (p = 0.0021) entre la biomasa del alga de los esteros y los valores promedio de la biomasa algal en las condiciones de cultivo, con un incremento de esta, tres veces mayor que la biomasa promedio de las poblaciones naturales. Este aumento significativo esta relacionado con las condiciones fisicoquímicas relativamente estables de la piscina, especialmente los valores de salinidad que oscilaron a lo largo del período de 22.6 a 27.1. A partir de la evaluación de la biomasa algal en los esteros, se recomienda desarrollar estrategias de cultivo de la especie, puesto que las poblaciones naturales no sostendrían su aprovechamiento comercial. Con el fin de incrementar la potencialidad de su cultivo se requiere desarrollar experiencias adicionales que permitan evaluar la incidencia de otros factores ambientales sobre la tasa de crecimiento relativo de la especie en condiciones de cultivo.
PALABRAS CLAVES: Acuicultura, Caulerpa sertularioides, macroalgas, crecimiento, Pacífico colombiano.
ABSTRACT
The biomass of natural populations and biomass under cultured conditions in shrimp ponds of the green alga Caulerpa sertularioides (Chlorophyta, Bryopsidales) was evaluated during March-December 2010 and January- June 2011. The algal biomass in the estuary was collected monthly at low tide using 625 cm2 quadrants. Water quality variables, such as dissolved oxygen, salinity, temperature, Secchi disk, NO3-, NH3 and PO43-, were taken simultaneously with algal samples. Algal biomass was taken to the lab to remove debris. Afterward, wet and dry weights were obtained to get final biomass. Relative growth rate (RGR% d-1) in ponds as well as physicochemical variables was evaluated weekly. Growth rates of plants registered a mean of 1.69 % day-1 (± 0.97). There were no significant differences (p = 0.315) in terms of biomass of C. sertularioides between Natal and Pajal creeks, showing averages between 4.09 and 4.67 g cm-2. Algal biomass showed significant differences between seasons (p = 0.000039), with highest values recorded between July and September. Salinity in Natal creek showed a direct relationship with seasonal variation of algal biomass (p = 0.019), with highest values of biomass in July-September related to higher levels of salinity in the study area. In contrast, salinity values and algal biomass in Pajal creek did not show significant relationship (p = 0.97). Nitrates, ammonia nitrogen, and orthophosphate were not significant with respect to changes in algal biomass (p = 0.93; p = 0.33; p = 0.55, respectively). Significant differences (p = 0.0021) between biomass of natural populations and algal biomass in shrimp ponds were reported, with higher values in cultured conditions, being those three times the biomass of natural creeks. Values of algal biomass from natural creeks suggested the importance of perform culture techniques, since natural populations would not sustain a commercial exploitation of this species. This study reports the first data of growth of a potential algal species and it is necessary to develop additional experiments to assess the impact of additional environmental factors on the relative growth rate.
KEYWORDS: Aquaculture, Caulerpa sertularioides, growth rates, Colombian Pacific, seaweeds.
INTRODUCCIÓN
La maricultura de algas se ha desarrollado en los últimos años especialmente con los géneros Kappaphycus, Lessonia, Macrocystis y Gracilaria, principalmente en Chile, China y Filipinas (Riaño y De la Ossa, 1999; Robledo, 2002; Bellorin et al., 2004; Buschmann et al., 2008). Particularmente, en América Latina, el cultivo de agarofitas, como el caso de Gracilaria y Gelidium, ha contribuido significativamente a la producción mundial de algas (Troel et al., 2003).
La creciente demanda de materia prima por parte de la industria, unida a la sobreexplotación y destrucción de praderas naturales, ha potenciado el desarrollo de sistemas de producción como alternativa al suministro de biomasa (Zertuche, 1993; Robledo, 1996; Draget et al., 2005; Buschmann et al., 2008). Los métodos comerciales empleados en el cultivo de algas tienen la característica de ser sencillos y requieren de poca inversión, y todos ellos se basan en la propagación vegetativa del alga (McHugh, 2003). En general, consisten en cultivos vegetativos intermareales o submareales donde manojos del alga son sujetados al substrato por medio de cuerdas, mangas de polietileno o directamente enterrados en el sedimento. Los principales factores que son tenidos en cuenta para las condiciones de cultivo son, entre otros, la fijación al substrato, la selección de la cepa, tipos de depredadores, los patrones de sedimentación, las densidades del cultivo y los métodos de cosecha (Hernández- Carmona et al., 2001; Wikfors y Ohno, 2001; Peña et al., 2005). El sector acuícola en Colombia cuenta con pocos actores que integren la cadena productiva de explotación de macroalgas, en especial en las fases de cultivo y procesamiento (Perry, 2002; Peña y Álvarez, 2006).
En los últimos años se han realizado trabajos en ecología y biología de algas de importancia económica, ofreciendo información básica sobre su distribución, biomasa y química de ficocoloides (McHugh, 2002; Hernández-Carmona et al., 2012). Específicamente en Colombia se han realizado diversos esfuerzos para evaluar el posible uso industrial de las algas, así como de los cultivos que pudieran suplir la cantidad necesaria de materia prima, analizando la factibilidad del cultivo de las rodófitas Grateloupia filicina, Euchema isiforme y Kappaphycus alvarezii, en asociación con las comunidades wayúu a lo largo del litoral de La Guajira en un sector de 25 km en el Cabo de la Vela (Peña y Álvarez, 2006). En años posteriores en la región de Santa Marta, Delgadillo y Newmark (2008) realizaron pruebas piloto para el cultivo de las algas rojas Gracilaria cervicornis, G. blodgettii, G. mamillaris e Hypnea musciformis obteniendo resultados importantes para el avance de esta actividad en el país.
El alga verde Caulerpa sertularioides (S. G. Gmelin) M. Howe (Chlorophyta: Bryopsidales) tiene distribución en la franja tropical y subtropical (Scrosati, 2001) y con frecuencia se encuentra en la zona intermareal alta, así como en esteros, lagunas costeras (Fama et al., 2002). Los estudios poblacionales de esta especie son una base para conocer las relaciones ecológicas de acuerdo con las condiciones ambientales de la zona de estudio. Se han llevado a cabo estudios para evaluar científicamente la relación entre esta macroalga y el desarrollo del camarón café (Porchas et al., 1999) y, a raíz de estos resultados, el alga se clasifica como un recurso promisorio para su cultivo potencial teniendo en cuenta sus principios bioactivos y amplia distribución en áreas tropicales y subtropicales. A principios de la década de los noventa, la industria camaronera en la región de Tumaco se vio afectada por el virus del taura y la presencia de la mancha blanca causando estragos en el sector, lo que ha conllevado a un deterioro y subutilización de la infraestructura instalada para el cultivo del camarón (Newmark et al., 2009). El objetivo de esta investigación fue determinar la variación espacio-temporal de la biomasa del alga verde Caulerpa sertularioides en poblaciones naturales y en condiciones de confinamiento en estanques de camarón de la ensenada de Tumaco (Nariño), con el propósito de evaluar el establecimiento potencial de su cultivo en la costa pacífica colombiana.
ÁREA DE ESTUDIO
La bahía de Tumaco está localizada en el sur de la costa pacífica colombiana, en el departamento de Nariño entre las latitudes 1°45' y 2°00'N y las longitudes 78°30' y 78°45'O (Figura 1), comprendiendo un área aproximadamente de 350 km2 con profundidades que varían entre 0 y 50 m (Garay-Tinoco et al., 2006). Una rica hidrografía caracteriza la bahía, conformada por importantes tributarios de la cuenca Pacífica, además de una extensa zona de esteros, siendo los más reconocidos: Chontal, Natal, Aguaclara, Pajal, Palmichal, Curay y Llanaje.
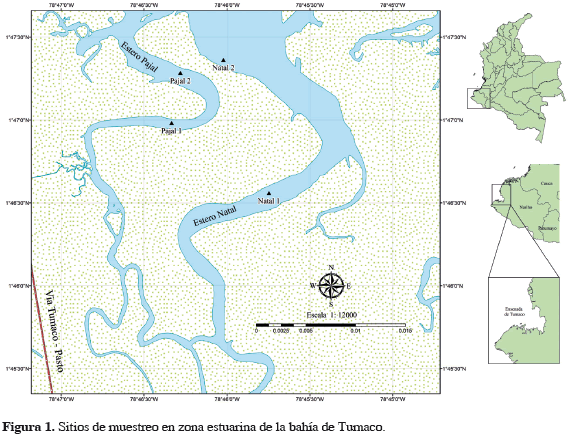
La bahía de Tumaco se comporta como un sistema estuarino presentando un intercambio continuo de aguas oceánicas con aguas continentales provenientes de la cuenca de los ríos Mira, Rosario, Mejicano y Curay. El régimen mareal semidiurno de la bahía determina las condiciones de calidad del agua, provocando la dilución o concentración de nutrientes, la variación de otros parámetros fisicoquímicos y la determinación de los procesos propios del ecosistema (Garay-Tinoco et al., 2006). Las condiciones oceanográficas del cuerpo costero están relacionadas con la composición florística de las algas en la bahía, se sabe que la presencia de ciertas especies está influenciada por la salinidad y el tipo de sustrato (Peña et al., 2005; Celis, 2008). Los esteros Natal y Pajal, donde se encontraron poblaciones naturales de Caulerpa sertularioides están bordeados por bosque de manglar dominado por Rhizophora mangle en la parte externa, Laguncularia sp. y Avicennia sp. en la zona intermareal y Conocarpus sp. en interior de bosque.
MATERIALES Y MÉTODOS
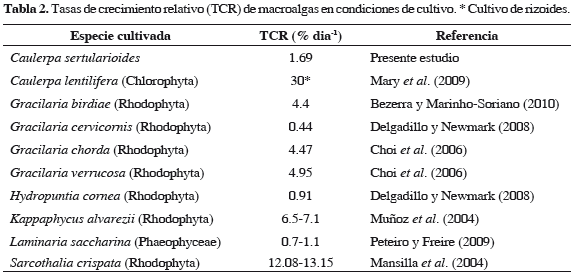
Para el monitoreo de la biomasa de las poblaciones naturales, se colectaron quincenalmente especímenes de C. sertularioides a nivel submareal en seis sitios presentes, con un cuadrante de 625 cm2 donde se midieron niveles de salinidad, oxígeno disuelto, temperatura, transparencia, NO3-, NH3, PO43- entre marzo y diciembre de 2010, en los esteros Natal (01°46'46"N y 78°45'56"O) y Pajal (01°47'08"N y 78°46'23"O) (Figura 1). La concentración de nutrientes (NO3-, NH3 y PO43-) en el agua fue registrada en el período marzo-julio y octubre-diciembre de 2010 y en el segundo trimestre de 2011. En el laboratorio las algas se lavaron con agua de mar, se limpiaron para retirar sedimento y otros organismos y posteriormente se llevaron a un tanque con agua de mar natural. El cultivo se llevo a cabo en una piscina de 0.15 ha, en la finca San Luis de la compañía Agromarina Tumaco (AMT) localizada en el municipio de Tumaco. Para los experimentos del cultivo, se seleccionó inicialmente una biomasa de 2400 g extraída de poblaciones naturales de C. sertularioides, sin alteraciones en su fronda con pesos entre 3 y 11 g, y se sembraron en 18 surcos de 5 m de largo por 30 cm de ancho, cada 20 cm con profundidad de 60 cm. Semanalmente se realizaron labores de mantenimiento y limpieza de las algas en la piscina. El crecimiento fue monitoreado por seis semanas, luego de las cuales se calculó la tasa de crecimiento relativo (TCR % d-1) expresada como porcentaje de crecimiento diario y se derivó de la fórmula de interés compuesto, propuesta por varios autores (Areces, 1995; Anderson et al., 1997; Marinho-Soriano et al., 2002; Delgadillo y Newmark, 2008).
TCR = [(Pf/Pi)1/t-1] x 100 (Ecuación 1)
donde,
Pf es peso final
Pi es peso inicial
t es intervalo de tiempo transcurrido entre ambas observaciones
A partir de los valores de biomasa mensual del alga en los esteros, se estimó la producción anual de la biomasa (g m-2) de la especie en peso húmedo (Buschmann et al., 1996).
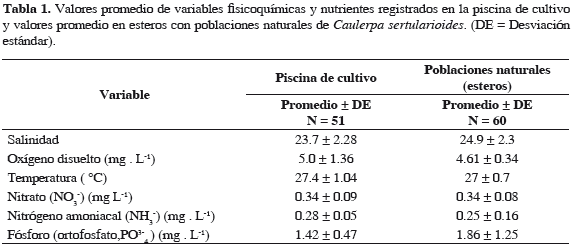
Variables fisicoquímicas como temperatura, salinidad y oxígeno disuelto fueron tomadas dos veces por día con sonda multiparamétrica YSI 85. Semanalmente se tomaron muestras de agua de la piscina de cultivo y quincenal de los esteros donde se encontró población natural, para análisis de nitratos, nitrógeno amoniacal y ortofosfato en espectrofotómetro HACH DR-2700, así como la transparencia del agua con un disco Secchi.
Los análisis estadísticos se realizaron con el programa Minitab 15. Se realizaron análisis de varianza para comparar variación espacial y temporal de biomasa del alga en poblaciones naturales entre los esteros y regresiones lineales para establecer relación entre la variación de biomasa de C. sertularioides y las variables fisicoquímicas.
RESULTADOS
Variables fisicoquímicas
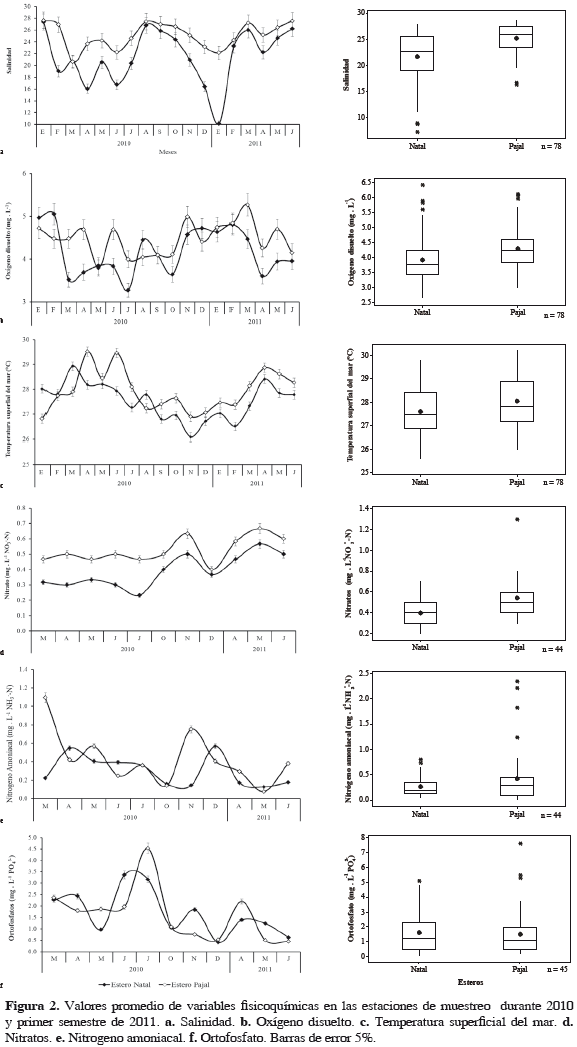
En los esteros Natal y Pajal se registraron valores promedio de salinidad de 21.3 (± 4.0) y 24.9 (± 2.3), respectivamente, con disminuciones para el segundo y tercer trimestre de 2010 en Natal. En los niveles de oxígeno disuelto en el estero Natal para 2010 se registraron valores alrededor de 4.14 mg L-1 (± 0.85) y entre enero y junio de 2011 de 4.23 mg L-1 (± 0.47). Para el estero Pajal se registraron valores promedio de 4.37 mg L-1 (± 0.69) en 2010 y de 4.61 mg L-1 (± 0.34) para el primer semestre de 2011. La temperatura superficial promedio registrada fue de 27° C ± 0.7 para ambos esteros (Figura 2a, 2b y 2c). El nitrato durante el período de estudio presentó un ámbito entre 0.34 (± 0.08) y 0.51 (± 0.05) mg L-1, con incrementos en 2011 hasta 0.6 mg L-1 en ambos esteros, mientras en el nitrógeno amoniacal se observaron disminuciones para cada período con promedios más bajos en 2011 de 0.16 (± 0.03) y 0.25 (± 0.16) mg L-1 respectivamente, sin embargo en noviembre de 2010 se registró un promedio de 0.75 mg L-1 en el estero Pajal y 0.57 mg L-1 en diciembre para el estero Natal (Figura 2d, 2e y 2f).
Dinámica de la biomasa en poblaciones naturales
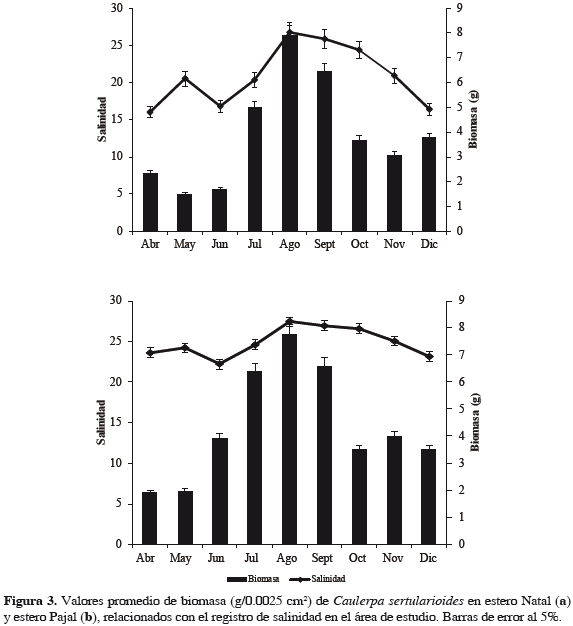
De acuerdo con el análisis de varianza realizado a nivel espacial no se presentaron diferencias significativas de biomasa de C. sertularioides, registrándose promedios de 4.09 (± 3.04) y 4.67 (± 2.81) g cm-2 (p = 0.315) para el estero Natal y Pajal, respectivamente. A nivel temporal, se encontraron diferencias significativas entre los meses de muestreo (p = 0.000039), con los mayores valores registrados entre julio y septiembre. Los valores de salinidad para el estero Natal presentan relación con la variación temporal de biomasa (R2 = 18.6, p = 0.019) registrándose los mayores valores de biomasa entre julio y septiembre de 2010 coincidiendo con los promedios de salinidad superiores a los otros meses, mientras en el estero Pajal no existe tal relación (R2 = 1.5, p = 0.97) (Figura 3). Factores como NO3-, NH3 y PO43- no tienen relación alguna con la variación de biomasa de C. sertularioides (R2 = 0, p = 0.93; R2 = 8.2, p = 0.33; R2 = 0.5, p = 0.55; respectivamente). Un descenso gradual de estos factores se observó durante el segundo y cuarto trimestre de 2010, coincidiendo con la época de lluvias, con un incremento de precipitación superior a 70% con respecto a los registros históricos para la época (Ideam, 2010a, 2010b).
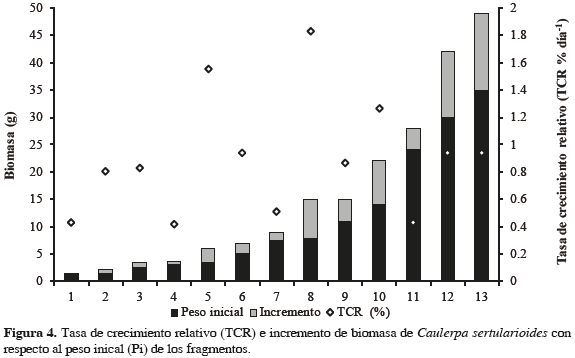
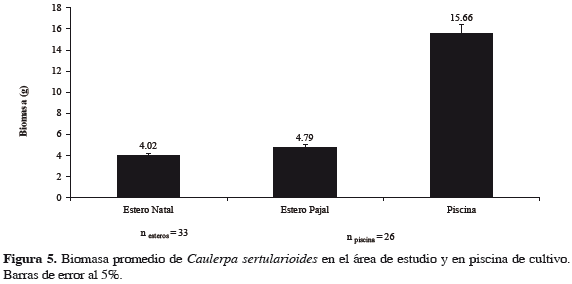
Dinámica de la biomasa en estanques El crecimiento fue monitoreado por seis semanas, luego de las cuales se calculó la tasa de crecimiento relativo (TCR % d-1) para 15 fragmentos, obteniendo valores promedio de 1.69% día-1 (± 0.97), con un valor mínimo de 0.47% día-1 y máximo de 3.20% día-1 (Figura 4). Se midieron variables fisicoquímicas en el estanque registrando valores promedio de 23.7 de salinidad, 5.0 mg de oxígeno disuelto, 0.34 mg L-1 de NO3-, 0.28 mg L-1 de NH3- y 1.42 mg L-1 de PO43-, los cuales son consistentes con los valores registrados en los esteros donde se encuentran poblaciones naturales de esta especie (Tabla 1). Se encontraron diferencias significativas (p = 0.0021) entre la biomasa del alga de los esteros y los valores promedio de la biomasa algal en las condiciones de cultivo, con esta aproximadamente cuatro veces mayor que la biomasa promedio de las poblaciones naturales (Figura 5).
DISCUSIÓN
De acuerdo con los resultados obtenidos se observó un patrón de correlación entre los registros de salinidad y la biomasa de las poblaciones naturales de C. sertularioides, presentándose una disminución notoria de su biomasa en los meses de mayor precipitación (segundo y cuarto trimestres de 2010). En la época de estudio cuando fue particularmente acentuada, siendo superior al 70% del promedio de los registros históricos (Ideam 2010a, 2010b). Debido a esta anomalía en la precipitación, en estos meses se registraron valores de salinidad inferiores al promedio mensual.
Las aguas costeras de la ensenada de Tumaco presentan condiciones estuarinas al tener salinidades promedio de 22.9 ± 8.8 y concentraciones fluctuantes de los nutrientes inorgánicos disueltos (Vivas-Aguas et al., 2010). Entre los factores más relevantes que regulan el crecimiento y distribución de macroalgas en ambientes tropicales son la salinidad y la disponibilidad de nitrógeno (Lapointe et al., 2005; Delgadillo y Newmark, 2008; Burfeind y Udy, 2009; Demes et al., 2010; Lapointe y Bedford, 2010). No se encontró una relación entre la distribución temporal de C. sertularioides y los niveles de nitrato y nitrógeno amoniacal, aunque se observaron incrementos de estos en el último trimestre de 2010, período en el cual hubo una reducción significativa de poblaciones naturales, coincidiendo con el período de mayor precipitación registrado para la época. Las variaciones de salinidad pueden afectar significativamente el crecimiento, distribución y productividad de muchas especies de macroalgas como resultado del estrés osmótico que puede inhibir estos procesos (Kamer y Fong, 2000; Jahnke y White, 2003; Xia et al., 2004); y aunque la mayoría de las algas marinas puede tolerar fluctuaciones de salinidad de corto plazo, grandes variaciones en esta variable pueden afectar significativamente algunos de los procesos bioquímicos involucrados en fotosíntesis y crecimiento.
El cultivo de C. sertularioides en estanques para camarón, presentó un comportamiento estable, teniendo en cuenta que no se observó deterioro de frondas ni pérdidas por epifitismo durante el período de estudio. Sin embargo, la TCR fue baja comparada con registros de otras macroalgas (Tabla 2), probablemente por alta radiación solar en la piscina. Otros estudios, llevados a cabo por Friedlander et al. (2006) con cultivo de rizoides de C. prolifera, logró tasas de crecimiento semanal superiores a 30%, concluyendo que las condiciones óptimas de cultivo están relacionadas con adecuada disponibilidad de sustratos arenosos, concentraciones adecuadas de nutrientes, columna de agua con niveles bajos de turbidez y valores relativamente bajos de radiación solar. Los valores significativamente altos de la biomasa del alga en condiciones de cultivo, con promedios tres veces mayores que la biomasa de poblaciones naturales, están relacionados con las condiciones físico-químicas relativamente estables del estanque, especialmente los valores de salinidad que oscilaron a lo largo del periodo, en un ámbito de 22.6 a 27.1. Con base en estos resultados, se sugiere que el uso potencial de Caulerpa solo sería viable a partir de cultivos en estanques, por cuanto la biomasa de las poblaciones naturales no podría sostener un aprovechamiento comercial de la especie. Proyecciones sobre la biomasa natural de Caulerpa, a partir de los datos obtenidos en este estudio, estimaron producciones entre 5.6 a 12.7 toneladas año-1 para la bahía de Tumaco. Las estadísticas de la explotación de algas rojas en Chile, para el caso de Gracilaria chilensis, mostraron una producción anual de 72 toneladas año-1 a partir de sus poblaciones naturales. En contraste, la producción de biomasa de cultivos intermareales osciló entre 120 y 198 toneladas año-1 (Buschmann et al., 2008), lo que indica que, a pesar de las altas cosechas anuales de las poblaciones naturales de la especie, una estrategia sostenible del uso del recursos algal a partir de cultivos, es la alternativa mas adecuada para el manejo de este recurso.
A partir de experiencias de cultivo de macroalgas en el Caribe colombiano por Peña y Álvarez (2006) y Delgadillo y Newmark (2008) este estudio contribuye como un caso piloto nuevo para el país para el aprovechamiento del recurso algal en la región. Con el fin de incrementar la potencialidad del cultivo de esta especie será necesario desarrollar experiencias adicionales que permitan evaluar la incidencia de los factores ambientales sobre la TCR. Paralelamente, se recomienda estudiar cultivos integrados con otras especies comerciales como el caso del camarón, estableciendo así cultivos multitróficos, que consisten en la diversificación de productos mediante la integración de cultivos de especies de diferentes niveles tróficos en el mismo sistema. Estas experiencias conllevarían a una producción efectiva a favor del sector acuícola en la costa pacífica colombiana.
AGRADECIMIENTOS
Esta investigación se desarrolló en el marco del proyecto "Adaptación, crecimiento y reproducción de especies promisorias para el desarrollo productivo en el Pacífico colombiano: caso algas marinas", financiado por Colciencias (1106- 403-20653) y ejecutado por la Universidad del Valle. Los autores agradecen especialmente a la bióloga Zuleyma Mosquera por su apoyo en los muestreos, a la Sociedad Agromarina Tumaco S. A. por su apoyo en facilitar la infraestructura de la finca San Luis en Tumaco y al personal operativo de la misma, Alexander Rodríguez y Max Meza, por su valioso apoyo en la colectas del material algal y monitoreo del sistema de cultivo.
BIBLIOGRAFÍA
1 Anderson, R. J., G. J. Levitt y A. Share. 1997. Experimental investigations for the mariculture of Gracilaria in Saldanha Bay, South Africa. J. Appl. Phycol., 8: 421-430. [ Links ]
2 Areces, J. A. 1995. Cultivo comercial de carragenófitas del género Kappaphycus Doty. 529-550. En: Alveal, K., M. E. Ferrario, E. C. Oliveira y E. Sar (Eds.). Manual de métodos ficológicos. Univ. Concepción, Concepción, Chile. 863 p. [ Links ]
3 Bellorin, A. M., M. C. Oliveira y E. C. Oliveira. 2004. Gracilaria vermiculophylla: A western Pacific species of Gracilariaceae (Rhodophyta) first recorded from the eastern Pacific. Phycol. Res., 52: 69-79. [ Links ]
4 Bezerra, A. F. y E. Marinho-Soriano. 2010. Cultivation of the red seaweed Gracilaria birdiae (Gracilariaceae, Rhodophyta) in tropical waters of northeast Brazil. Bio. and Bioe., 34: 1813-817. [ Links ]
5 Burfeind, D. D. y J. W. Udy. 2009. The effects of light and nutrient on Caulerpa taxifolia and growth. Aqua. Bot., 90: 105-109. [ Links ]
6 Buschmann, A. H., D. A. López y A. Medina. 1996. A review of the environmental effects and alternative production strategies of marine aquaculture in Chile. Aquacult. Eng., 15 (6): 397-421. [ Links ]
7 Buschmann, A., M. C. Hernández-González y D. Varela. 2008. Seaweed future cultivation in Chile: perspectives and challenges. Int. J. Environ. Poll., 33 (4): 432-456. [ Links ]
8 Celis, C. N. 2008. Evaluación espacio-temporal de la calidad fisicoquímica del agua en las bahías de Buenaventura y Málaga. Dirección General Marítima-Centro Control Contaminación del Pacífico (CCCP). http://www.cccp.org.co/index.php/component/content/article/92-calidad-ambientalmarina/668-evaluacion-espacio-temporal-de-la-calidad-fisicoquimica-del-agua-en-las-bahias-debuenaventura-y-malaga. Consultada 21/05/2014. [ Links ]
9 Choi, H. G., Y. S. Kim, J. H. Kim, S. J. Lee, E. J. Park, J. Ryu y K. W. Nam. 2006. Effects of temperature and salinity on the growth of Gracilaria verrucosa and G. chorda, with the potential for mariculture in Korea. J. App. Phycol., 18: 269-277. [ Links ]
10 Delgadillo, O. y F. Newmark. 2008. Cultivo piloto de macroalgas rojas (Rhodophyta) en Bahía Portete, La Guajira, Colombia. Bol. Invest. Mar. Cost., 37 (2): 7-26. [ Links ]
11 Demes, K. W., M. M. Littler y D. Littler. 2010. Comparative phosphate acquisition in giant-celled rhizophytic algae (Bryopsidales, Chlorophyta): Fleshy vs. calcified forms. Aqua. Bot., 92: 157-160. [ Links ]
12 Draget, K. I., O. Smidsrod y G. Skjak-Braek. 2005. Alginates from algae. 1-30. En: Steinbüchel, A. y S. K. Rhee (Eds.). Polysaccharides and polyamides in the food industry: Properties, production and patents I. Wiley-VCH, Weinheim, Alemania. 171 p. [ Links ]
13 Fama, P., B. Wysor, W. H. Kooistra y G. C. Zuccarello. 2002. Molecular phylogeny of the genus Caulerpa (Caulerpales Chlorophyta) inferred from chloroplast tufA gene. J. Phycol., 38: 1040-1050. [ Links ]
14 Friedlander, M., Y. Kosov, G. Keret y C. Dawes. 2006. Production of rhizoids by Caulerpa prolifera in culture. Aquatic Bot., 85: 263-266. [ Links ]
15 Garay-Tinoco, J. A., D. I. Gómez-López y J. R. Ortiz-Galvis (Eds). 2006. Diagnóstico integral del impacto biofísico y socioeconómico relativo a las fuentes de contaminación terrestre en la bahía de Tumaco, Colombia y lineamientos básicos para un plan de manejo. Proyecto del Programa de las Naciones Unidas para el Medio Ambiente (PNUMA-Programa de Acción Mundial PAM) y Comisión Permanente del Pacífico Sur (CPPS), Instituto de Investigaciones Marinas y Costeras (Invemar), Centro de Control Contaminación del Pacífico (CCCP), Corporación Autónoma Regional de Nariño (Corponariño), Santa Marta. 290 p. [ Links ]
16 Hernández-Carmona, G., D. Robledo y E. Serviere-Zaragoza. 2001. Effect of nutrient availability on Macrocystis pyrifera recruitment survival near its southern limit of Baja California. Bot. Mar., 44 (3): 221-229. [ Links ]
17 Hernández-Carmona, G., Y. E. Rodríguez-Montesinos, D. L. Arvizu-Higuera, R. Reyes-Tisnado, J. I. Murillo-Álvarez y M. Muñoz-Ochoa. 2012. Avances tecnológicos en la producción de alginatos en México. Ingeniería Investigación y Tecnología, 13 (2): 155-168. [ Links ]
18 Ideam. 2010a. Boletín informativo sobre el monitoreo del fenómeno de "La Niña". No. 23, Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia, Bogotá. 7 p. [ Links ]
19 Ideam. 2010b. Boletín informativo sobre el monitoreo del Fenómeno de "La Niña". No. 24, Instituto de Hidrología, Meteorología y Estudios Ambientales de Colombia, Bogotá. 7 p. [ Links ]
20 Jahnke, L. S. y A. L. White. 2003. Long-term hyposaline and hypersaline stresses produce distinct antioxidant responses in the marine alga Dunaliella tertiolecta. J. Plant Physiol., 160: 1193-1202. [ Links ]
21 Kamer, K. y P. Fong. 2000. A fluctuating salinity regime mitigates the negative effects of reduced salinity on the estuarine macroalga Enteromorpha intestinalis (L.) link. J. Exp. Mar. Biol. Ecol., 254: 53-69. [ Links ]
22 Lapointe, B. E. y B. J. Bedford. 2010. Ecology and nutrition of invasive Caulerpa brachypus f. parvifolia blooms on coral reefs off southeast Florida, U. S. A. Harm. Alg., 9: 1-12. [ Links ]
23 Lapointe, B. E., P. J. Barile, M. M. Littler, D. S. Littler, B. J. Bedford y C. Gasque. 2005. Macroalgal blooms on southest Florida coral reefs I. Nutrient stoichiometry of the invasive green alga Codium isthmocladum in the wider Caribbean indicates nutrient enrichment. Harm. Alg., 4: 1092-1105. [ Links ]
24 Mansilla, A., M. Palacios y S. Aguilar. 2004. Efecto de la salinidad en el desarrollo inicial de Sarcothalia crispata (Bory) Leister (Rhodophyta, Gigartinales) bajo condiciones de laboratorio. Anales Instituto Patagonia (Chile), 32: 13-23. [ Links ]
25 Marinho-Soriano, E., C. Morales y W. Moreira. 2002. Cultivation of Gracilaria (Rhodophyta) in shrimp ponds effluents in Brazil. Aquac. Res., 33: 1081-1086. [ Links ]
26 Mary, A., V. Mary, A. Lorella y J. R. Matias. 2009. Rediscovery of naturally occurring seagrape Caulerpa lentillifera from the Gulf of Mannar and its mariculture. Current Science, 97 (10): 1418-1420. [ Links ]
27 McHugh, D. J. 2002. Perspectivas para la producción de algas marinas en los países en desarrollo. FAO. Circular de Pesca No. 968 FIIU/C968. 30 p. http://www.fao.org/DOCREP/004/Y3550S/Y3550S00.HTM. 02/02/2008. [ Links ]
28 McHugh, D. J. 2003. A guide to the seaweed industry. FAO Fisheries Technical Paper, Número 441, Roma. 105 p. [ Links ]
29 Muñoz, J., Y. Freile-Pelegrín y D. Robledo. 2004. Mariculture of Kappaphycus alvarezii (Rhodophyta, Solieriaceae) color strains in tropical waters of Yucatán, México. Aquaculture, 239: 161-177. [ Links ]
30 Newmark, F., C. H. Valverde, J. M. Díaz, G. Parra, S. P. Bonilla y M. Salazar. 2009. Agenda prospectiva de investigación y desarrollo tecnológico para la cadena productiva de camarón de cultivo en Colombia. Ministerio de Agricultura y de Desarrollo Rural, Proyecto Transición de la Agricultura, Bogotá. 224 p. [ Links ]
31 Peña, E. y R. Álvarez. 2006. Experiencias en el cultivo experimental de algas rojas en el Caribe y Pacífico de Colombia. Rev. Luna Azul, 23: 16-20. [ Links ]
32 Peña, E. J., M. L. Palacios y N. Ospina-Álvarez. 2005. Algas como indicadoras de contaminación. Programa editorial Universidad del Valle, Cali. 164 p. [ Links ]
33 Perry, S. 2002. Estudio de la competitividad de la camaricultura colombiana. Ministerios de Agricultura y Desarrollo Rural. Asociación Nacional de Acuicultores de Colombia (Acuanal), Bogotá. 141 p. [ Links ]
34 Peteiro, C. y O. Freire. 2009. Effect of outplanting time on commercial cultivation of kelp Laminaria saccharina at the southern limit in the Atlantic coast, N.W. Spain. Chin. J. Oceanol. Limnol., 27 (1): 54-60. [ Links ]
35 Porchas, M. A., L. Martínez, F. Magallon, J. Naranjo y G. Portillo. 1999. Efecto de la macroalga Caulerpa sertularioides en el desarrollo del camarón Penaeus californiensis (Decapoda: Peniedae). Rev. Biol. Tropi., 47 (3): 437-442. [ Links ]
36 Riaño, R. y J. De la Ossa-Velásquez. 1999. Guía para el manejo, cría y conservación de la ostra de mangle (Crassostrea rhizophorae. Guilding). Convenio Andrés Bello, Bogotá. 64 p. [ Links ]
37 Robledo, D. 1996. Conocimiento de la macroflora marina de interés económico de las costas de Yucatán. Informe final del proyecto B077. Instituto Politécnico Nacional, Centro de Investigación y de Estudios Avanzados Mérida, Departamentos de Recursos del Mar, Laboratorio de Ficología, Informe final, Mérida, México. 42 p. [ Links ]
38 Robledo, D. 2002. Agronomía marina: una alternativa para el desarrollo de las comunidades pesqueras de Dzilam de Bravo, Yucatán. Departamento de Recursos del Mar, Centro de Investigación y de Estudios Avanzados. Mérida, México. 49 p. [ Links ]
39 Scrosati, R. 2001. Population dynamics of Caulerpa sertularioides (Chlorophyta: Bryopsidales) from Baja California, Mexico, during El Nino and La Nina years. J. Mar. Biol. Assoc., 81 (5): 721-726. [ Links ]
40 Troell, M., C. Halling, A. Neori, A. H. Buschmann, T. Chopin, C. Yarish y N. Kautsky. 2003. Integrated mariculture: asking the right questions. Aquaculture, 226: 69-80. [ Links ]
41 Vivas-Aguas, L., M. Tosic, J. Sánchez, S. Narváez, B. Cadavid, P. Bautista, J. Betancourt, J. Parra, L. Echeverry y L. Espinosa. 2010. Diagnóstico y evaluación de la calidad ambiental marina en el Caribe y Pacífico colombiano. Red de vigilancia para la conservación y protección de las aguas marinas y costeras de Colombia (Recam). Informe técnico 2010. Instituto de Investigaciones Marinas y Costeras (Invemar), Santa Marta. 208 p. [ Links ]
42 Wikfors, G. H. y M. Ohno. 2001. Impact of algal research in aquaculture. J. Phycol., (37): 968-974. [ Links ]
43 Xia, J. R., Y. J. Li y D. H. Zou. 2004. Effects of salinity stress on PSII in Ulva lactuca as probed by chlorophyll fluorescence measurements. Aquat. Bot., 80: 129-137. [ Links ]
44 Zertuche-G, J. A. (Ed.). 1993. Situación actual del cultivo de algas agarofitas en América Latina y el caribe. Instituto de Investigaciones Oceanológicas, Universidad Autónoma de Baja California. Ensenada, BC. México. http://www.fao.org/docrep/field/003/AB483S/AB483S01.htm. 01/09/2011. [ Links ]
FECHA DE RECEPCIÓN: 02/12/2011 FECHA DE ACEPTACIÓN: 15/02/2014
