










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
versión impresa ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.44 no.2 Santa Marta jul./dic. 2015
TEREBÉLIDOS (ANNELIDA: POLYCHAETA: TEREBELLIDAE) DE ISLA FUERTE, CARIBE COLOMBIANO
TEREBELLIDS (TEREBELLIDAE: POLYCHAETA: ANNELIDA) FROM ISLA FUERTE, COLOMBIAN CARIBBEAN.
Isabel C. Molina-Acevedo y Mario H. Londoño-Mesa
Instituto de Biología, Grupo LimnoBasE y Biotamar, Facultad de Ciencias Exactas y Naturales, Universidad de Antioquia, Medellín, Colombia. isacrismoliace@gmail.com, imolina@ecosur.edu.mx, hernan.londono@udea.edu.co.
RESUMEN
La familia Terebellidae ha sido poco estudiada en Colombia; la información disponible ha sido reciente y solo en forma de listas faunísticas, las cuales suman 17 especies en el nororiente del Caribe colombiano. Los estudios taxonómicos de terebélidos que incluyan descripciones detalladas son prácticamente nulos. Esta investigación tuvo como objetivo evaluar taxonómicamente la diversidad de los terebélidos en Isla Fuerte (Caribe colombiano), así como aumentar el conocimiento de las especies hacia el suroccidente de la región. Se analizaron los organismos recolectados en aguas someras de siete playas, asociados a roca coralina y algas calcáreas. En total, seis géneros y diez especies de terebélidos fueron identificados. Eupolymnia rullieri, Lanicola carus, Pista palmata y Terebella verrilli ya habían sido registradas previamente para el Caribe colombiano. Polycirrus sp.1, Polycirrus sp.2 y Pista sp. son especies que aún no se describen debido a la falta de más material. Loimia minuta, Pista cetrata y Polycirrus angeli se encontraron como nuevos registros para la región. Por lo tanto, el número de especies de la familia para Caribe colombiano se aumenta a 20 y se amplía el ámbito de distribución de las últimas tres especies para el país. Se ofrece información relevante en cuanto al patrón de tinción con verde de metilo y a la variación morfológica de los ejemplares encontrados para cada especie.
PALABRAS CLAVES: Caribe colombiano, Poliquetos, Taxonomía, Caracteres morfológicos, Descripciones.
ABSTRACT
The family terebellidae has been poorly studied in colombia. The only available information on the family presented is in term of lists, which comprise 17 species occurring in the northeastern Colombian Caribbean. Taxonomic studies including detailed descriptions are none. This research focused on taxonomic evaluation of terebellids diversity from Isla Fuerte, Colombian Caribbean, as well as on the increasing the species knowledge for the southwestern region. Specimens from coralline rock and calcareum algae were collected and analyzed from seven beaches around the island. Six genera and ten species were found: Eupolymnia rullieri, Lanicola carus, Pista palmata, and Terebella verrilli have been already identified in the Colombian Caribbean; Polycirrus sp.1, Polycirrus sp.2 and Pista sp. remain undescribed due the lack of enough material justifing new species category. Finally, Loimia minuta, Pista cetrata and Polycirrus angeli are new records for Colombia. After this research, the number of recorded terebellid species increased to 20, and the species distributions were enlarged. Staining patterns and morphologic variation for each species are provided.
KEY WORDS: Colombian Caribbean, Polychaetes, Taxonomy, Morphological characters, Descriptions.
INTRODUCCIÓN
Los terebélidos son denominados gusanos espagueti por su apariencia cuando tienen sus tentáculos retraídos. El tamaño de los gusanos puede variar desde menos de 1 cm (e.g. Polycirrus spp.) hasta 15 cm (e.g. Loimia spp.). Se distribuyen desde aguas por debajo de la línea mareal hasta grandes profundidades. Los terebélidos son gusanos tubícolas considerados un componente importante del bentos, puesto que son responsables de la remoción de altas cantidades de sedimento del suelo marino. Construyen sus tubos utilizando diferentes partículas dispuestas en el fondo, tales como granos de arena, restos de algas calcáreas, fragmentos de corales, conchas y fragmentos de pastos marinos, entre otros. Los gusanos espagueti se encuentran generalmente debajo de piedras, entre macroalgas o fondos arenosos. Presentan hábitos sedimentívoros, consumiendo microalgas, bacterias y otros organismos microscópicos. Para alimentarse, extienden sus tentáculos ciliados hasta cinco veces el tamaño corporal para transportar las partículas de sedimento hacia la boca.
De acuerdo con Londoño-Mesa (2009b), en el Caribe hay 24 géneros y 44 especies de terebélidos. No obstante, el estudio de la familia ha sido fragmentado, dependiendo principalmente de la disponibilidad de material recolectado y depositado en diversas colecciones. Así, la abundancia del material estudiado proviene de aguas poco profundas, principalmente de México y Venezuela, siendo rezagados los estudios taxonómicos detallados en los demás países. En Colombia, el estudio de los gusanos espagueti ha sido poco. Báez y Ardila (2003) hicieron una lista de especies de poliquetos registradas en el Caribe colombiano durante los últimos 30 años, siendo Terebellidae una de las familias más ricas en géneros (8) y la sexta más rica en especies (11). Más tarde, Londoño-Mesa (2011) realizó una revisión y discusión taxonómica del mismo material de terebélidos y demostró que en realidad este material está representado por 10 géneros y 17 especies en el Caribe colombiano.
El propósito de esta investigación fue hacer una revisión y descripción taxonómica de las especies de la familia Terebellidae encontradas en Isla Fuerte, Caribe colombiano. Se tomaron muestras de diferentes sustratos en aguas someras de la isla, para estimar la diversidad taxonómica.
ÁREA DE ESTUDIO
Isla Fuerte (Figura 1) está localizada a 11 km del continente (9°23’N 76°11’W para Puerto Limón) sobre la plataforma continental al suroeste del Caribe colombiano, formando parte de la cadena de islas de origen coralino pertenecientes a Cartagena (Bolívar). La isla tiene 3.25 km2 de superficie emergida y una elevación máxima de 12 m. En la zona marina, los fondos duros son arrecifes coralinos con un área de 6.24 km2, mientras que los fondos blandos son praderas de pastos marinos con 33 km2 y pocos parches de bosques de manglar al norte y al sur. La isla presenta una ligera inclinación hacia el este, en su eje norte-sur, y una porción emergida en el sector NE (Estela, 2006).
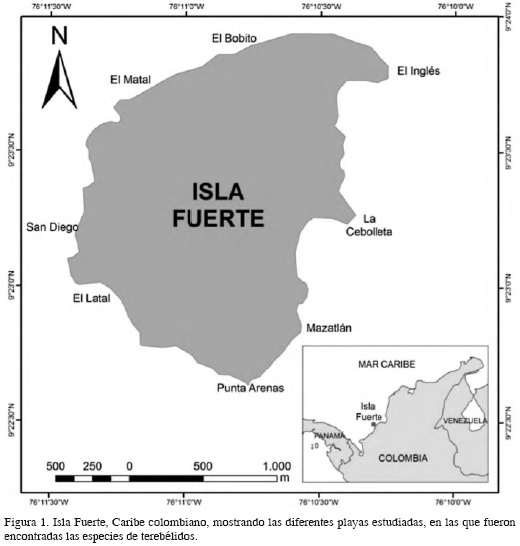
MATERIALES Y MÉTODOS
Se estudiaron los terebélidos de siete playas de Isla Fuerte (Tabla 1) durante 2009-2011. Las muestras fueron recolectadas durante las salidas de campo del curso de Zoología de Invertebrados de la Universidad de Antioquia y se depositaron en la Colección Estuarina y Marina de la misma universidad (CEMUA). Los ejemplares se fijaron en formol al 4 % diluido con agua marina, y se preservaron en alcohol etílico al 70 %. Los organismos provienen de dos tipos de sustratos: blandos (algas y sedimento), a una profundidad máxima de 1.5 m, y duros (roca coralina), extraídos entre 0 y 7 m de profundidad. Los sustratos blandos fueron procesados usando tamices de 0.5 y 0.25 mm de poro de red. Los sustratos duros fueron fragmentados usando cincel y martillo.

La identificación taxonómica se realizó mediante la observación bajo el microscopio compuesto y de disección. El material se rotuló adecuadamente y se depositó en la colección CEMUA, utilizando POLY para indicar la clase Polychaeta, y TERE para la familia Terebellidae. El número de especímenes para cada lote de individuos revisados se indicó con el número de éstos entre paréntesis. Se describieron los caracteres de importancia para cada especie, como los labios superior e inferior, membrana tentacular, ocelos, alas laterales, branquias, escudos ventrales, papilas nefridiales y notosetas. La descripción de los uncinos se realizó con base en Londoño-Mesa (2009b).
Se realizó una tinción usando como colorante de contraste verde de metilo en una solución saturada con alcohol etílico a 70%; esto permitió identificar los patrones de zonas glandulares que ayudaron a separar especies, así como a dar mayor contraste a estructuras con importancia taxonómica. Cada lote se identificó de acuerdo con las claves de Londoño-Mesa (2009a, 2009b). La descripción de los uncinos se realizó siguiendo la terminología sugerida por Holthe (1986).
RESULTADOS
Análisis taxonómico
Familia Terebellidae Johnston, 1846
Subfamilia Polycirrinae Malmgren, 1866
Polycirrus Grube, 1850
Polycirrus Grube, 1850: 597. Hutchings & Glasby 1986: 330. Hartmann-Schröder 1996: 524.
Especie tipo. Polycirrus medusa Grube, 1850, por monotipia.
Diagnosis. Membrana tentacular con tres lóbulos; dos tipos de tentáculos, ambos con extremos hinchados; ocelos ausentes; escudos ventrales divididos en pares por un surco medio-ventral; notópodos desde los segmentos 2 o 3; tórax con un número variable de segmentos con notosetas; setas pinadas, hirsutas, con limbos estrechos y lisos, o capilares lisos, pero sólo con uno o dos tipos de setas por especie; neurópodos pueden comenzar entre los segmentos 5 y 21 (Londoño-Mesa, 2009b).
Polycirrus angeli Londoño-Mesa, 2009 Figura 2 A-G
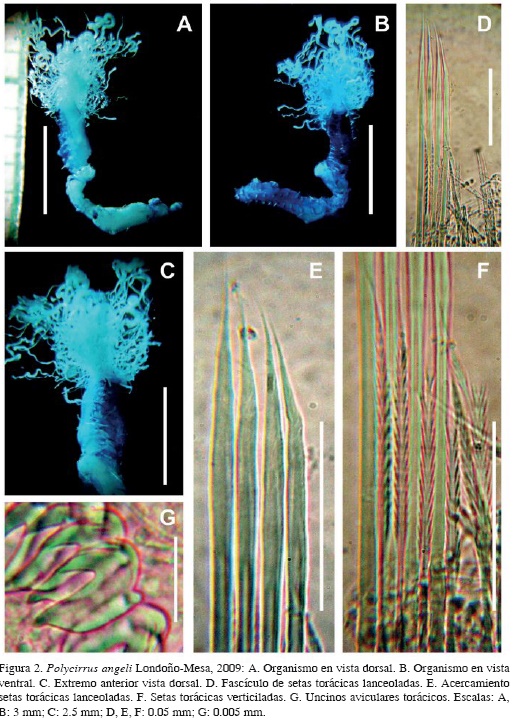
Polycirrus angeli Londoño-Mesa, 2009b: 21-22; Fig. 5D-G.
Material examinado. CEMUA POLY-TERE 0001 (1), Playa El Matal, 28.II.2011.
Descripción. Organismo incompleto, región distal torácica y anterior abdominal maltratadas (Figura 2A, B y C); 27 segmentos. Tórax 3.5 mm de largo y 0.8 mm de ancho. Dorso hinchado y largo, continuando con la membrana tentacular; con líneas intersegmentales claramente definidas. Numerosos tentáculos de dos tipos: cortos, gruesos distalmente y largos, delgados. Labio superior fusionado a la membrana tentacular. Labio inferior inflado, desplazando los primeros dos escudos ventrales. Membrana tentacular con un lóbulo, tan largo como cinco segmentos torácicos, plegado y dirigido anteriormente. Ocelos ausentes. Diez segmentos con escudos ventrales, iniciando desde el segmento 1; escudos medialmente divididos en dos por un surco ventral profundo; primer par de escudos delgado, triangular, dirigido lateralmente; segundo par más grueso, rectangular; tercer par más hinchado; últimos pares decrecientes, aunque poco distinguibles por el estado del organismo. Papilas nefridiales no visibles. Notópodos desde segmento dos hasta el final del fragmento. Notosetas de dos tipos (Figura 2D); lanceoladas, largas, limbos muy estrechos (Figura 2E); verticiladas, cortas (Figura 2F). Neurópodos desde el segmento siete, uncinos aviculares torácicos (Figura 2G) MF: 2-3:1-2; PP desarrollado, redondeado; PF ausente; Oc levemente cóncavo y corto; Cp con dientes de la segunda fila superando la mitad del MF; MF corto y grueso; Sr liso; SrP y SrA ausentes; AP proyectado anteriormente, redondeado; Bs cóncava anteriormente, y recta posteriormente. Pigidio no observado.
Patrón de tinción. Las líneas intersegmentales dorsales tiñen tenuemente; los escudos ventrales tiñen intensamente.
Discusión. Polycirrus angeli, descrita para Veracruz, golfo de México, difiere de las especies registradas por Kritzler (1984) para Florida y golfo de México en cuanto al tipo, cantidad y el segmento en donde incian las notosetas, y en el segmento en donde aparecen los uncinos. Dichas especies de Kritzler son las siguientes: P. eximius dubis Day, 1973, Polycirrus plumosus (Wollebaek, 1912), Polycirrus cf. albicans, Polycirrus cf. denticulatus, Polycirrus cf. haematodes, Polycirrus sp. A y Polycirrus sp. B. En P. angeli, los notópodos comienzan en el segmento dos, mientras que en algunas de las otras especies los notópodos comienzan en el segmento tres. Aunque este espécimen está incompleto, el número de segmentos con notosetas en la descripción original es mayor que la mayoría de esas especies; sólo P. eximius dubis tiene 41 setígeros. La especie en cuestión se diferencia de Polycirrus cf. haematodes, Polycirrus sp. B, P. eximius dubis y Polycirrus cf. albicans, por la presencia de dos tipos de setas: setas limbadas largas, y setas verticiladas cortas. Sólo Polycirrus plumosus, Polycirrus sp. A y Polycirrus cf. denticulatus poseen dos tipos de setas. Finalmente, en cuanto al número de segmento en el cual aparecen los uncinos, ninguna de las especies comparadas presenta uncinos desde el segmento siete; sólo P. eximius dubis presenta uncinos desde el segmento nueve. Por lo tanto, las diferencias presentadas en este carácter sirven para establecer que se trata de una especie diferente a las demás presentadas en el análisis.
Localidad tipo. Veracruz, golfo de México.
Distribución. Posiblemente en todo el golfo de México. Panamá, costa Caribe y sur del Caribe colombiano.

Material examinado. CEMUA POLY-TERE 0002 (1), Playa El Latal, 05.IX.2010.
Descripción. Organismo incompleto (Figura 3A y B), fragmento anterior con ocho segmentos, 2 mm de largo y 0.6 mm de ancho. Dorso hinchado y largo hasta el segmento seis. Numerosos tentáculos de dos tipos; cortos, gruesos distalmente y largos, delgados. Labio superior fusionado con la membrana tentacular (Figura 3C y D). Labio inferior hinchado y bien desarrollado. Membrana tentacular longitudinal larga, con pliegues en vista ventral; en forma de "U" invertida, en vista dorsal. Ocelos ausentes. Nueve segmentos con escudos ventrales, iniciando en el segmento uno; escudos medialmente divididos en dos, por un surco ventral profundo; últimos pares decrecientes, aunque poco distinguibles por el estado del organismo. Papilas nefridiales no visibles. Siete pares de notópodos largos, desde el segmento dos. Notosetas de dos tipos: lanceoladas largas, con limbos muy estrechos y verticiladas cortas (Figura 3E y F). Neurópodos desde segmento ocho, uncinos aviculares torácicos (Figura 3G) MF: 1-3:1-3; PP extendido y redondeado; PF ausente; Oc cóncavo y corto; Cp con diente central sobresaliendo más de la mitad del MF; MF grueso y corto; Sr recto; SrP y SrA ausentes; AP desarrollado y redondeado; AF delgado y dirigido hacia la base; Bs cóncava anteriormente y recta posteriormente. Pigidio no observado.
Patrón de tinción. Los escudos ventrales tiñen intensamente. Labio inferior tiñe claro en comparación con los primeros cuatro escudos. Dorso, membrana tentacular y labio superior sin tinción.
Discusión. Polycirrus sp. 1 comparte con P. angeli el segmento donde comienzan los notópodos y dos tipos de setas; sin embargo, se diferencian por el segmento de inicio de los uncinos, ya que en Polycirrus. sp. 1 aparecen en el 8 y en P. angeli surgen a partir del segmento 7. Igualmente, se diferencian en la forma de la membrana tentacular; donde la especie aquí mencionada presenta una membrana en vista dorsal hinchada y en forma de "U" invertida, mientras que P. angeli tiene una membrana tentacular larga y proyectada anteriormente, sin lóbulos laterales en vista dorsal. También difieren en la fórmula uncinal, donde Polycirrus sp. 1 posee un MF: 1-3:1-3, mientras que para P. angeli es MF: 2-3:1-2. Finalmente, la especie en cuestión tiene la base de los uncinos gruesa y presenta un proceso posterior con filamento.
Distribución. Playa El Latal, Isla Fuerte.
Polycirrus sp. 2
Material examinado. CEMUA POLY-TERE 0003 (1) Playa El Latal, 05.IX.2010.
Descripción. Organismo incompleto (Figura 3H), 10 segmentos. Tórax 3.1 mm de largo y 1 mm de ancho. Dorso con estrías circulares, levemente más hinchado en los primeros seis segmentos. Tentáculos no observados. Labio superior fusionado a la membrana tentacular (Figura 3I), dispuesto en tres lóbulos, uno proyectado anteriormente y otros dos hacia los lados del labio inferior. Labio inferior hinchado, formando dos lóbulos. Membrana tentacular con borde anterior grueso e hinchado dorsalmente (Figura 3J), que sobresale del primer segmento; ventralmente ancha, con las cicatrices de los tentáculos. Ocelos ausentes. Diez segmentos con escudos ventrales, a partir del segmento dos; escudos medialmente divididos en dos, por un surco ventral profundo; escudos de los segmentos tres al siete más hinchados; últimos pares decrecientes en tamaño, aunque poco distinguibles por el mal estado del organismo. Papilas nefridiales no visibles. Nueve pares de notópodos desde segmento dos, largos e hinchados en la base, con cirro posterior. Notosetas de dos tipos (Figura 3 K y L): largas, bilimbadas, serradas en el borde externo y cortas verticiladas, más numerosas, tan largas como la mitad de las setas largas. Neurópodos desde segmento 10, uncinos aviculares (Figura 3M) torácicos com MF: 1-2:1:1; PP redondeado y saliente; PF ausente; Oc levemente cóncavo y corto; Cp con dientes de la segunda fila alcanzando la mitad de la longitud del MF; MF largo, delgado y puntiagudo; USr y LSr levemente diferenciados por un SrP apenas visible; SrA ausente; AP redondeado y amplio, apenas sobresaliente; AF ausente; Bs cóncava. Pigidio no observado.
Patrón de tinción. Base anterior de los notópodos tiñen, se observa una textura granulosa debido a la presencia del tejido notopodial glandular. Escudos ventrales tiñen intensamente entre los segmentos dos y seis; el labio inferior tiñe más claro. Dorso, membrana tentacular y labio superior sin tinción.
Discusión. Polycirrus sp. 2 es similar a P. angeli y a Polycirrus sp. 1 en el segmento de inicio de los notópodos y por presentar dos tipos de setas. Sin embargo, las tres se diferencian por el segmento en el cual comienzan los uncinos; mientras que en Polycirrus sp. 2 los uncinos comienzan en el segmento 10, en P. angeli comienzan en el siete y en Polycirrus sp. 1 en el ocho. Igualmente, las tres se diferencian en la forma de la membrana tentacular, donde Polycirrus sp. 2 presenta una membrana en vista dorsal hinchada y levemente curva; P. angeli tiene la membrana larga y proyectada anteriormente, sin regiones hinchadas en vista dorsal y Polycirrus sp. 1 la tiene en forma de "U" invertida. La fórmula de los uncinos para Polycirrus sp. 1 es MF: 1-3:1-3, mientras que para Polycirrus sp. 2 es MF: 1-2:1:1, y para P. angeli es MF: 2-3:1-2. La base de los uncinos de las dos especies sin describir es similar, aunque Polycirrus sp. 2, al igual que P. angeli, no presenta proceso posterior con filamento.
Distribución. Playa El Latal, Isla Fuerte.
Subfamilia Terebellinae Grube, 1850 Eupolymnia Verrill, 1900
Eupolymnia Verrill, 1900: 660 (syn.). Hessle 1917: 174, Text. Fig. 41 (Polymnia). Hartmann-Schröder 1996: 509. Capa y Hutchings 2006:4. Londoño-Mesa 2009b: 26-27.
Especie tipo. Amphitrite nesidensis delle Chiaje, 1828, por designación subsecuente.
Diagnosis. Branquias en los segmentos dos al cuatro; ocelos algunas veces presentes; alas laterales en segmentos dos y tres o dos al cuatro; escudos ventrales desde el segmento dos; papilas nefridiales desde el segmento dos o tres, continuas o discontinuas, hasta los segmentos tres al nueve; 17 pares de notópodos desde el segmento cuatro, notosetas lisas, limbadas; neurópodos desde el segmento cinco, uncinos aviculares en una sola fila desde el segmento cinco hasta el 9-10; en doble fila, arreglados frente a frente, solo hasta el segmento 20; luego, en una fila (Londoño-Mesa, 2009b).
Eupolymnia rullieri Londoño-Mesa, 2009 Figura 4A-H
Eupolymnia rullieri Londoño-Mesa, 2011: 20-21.
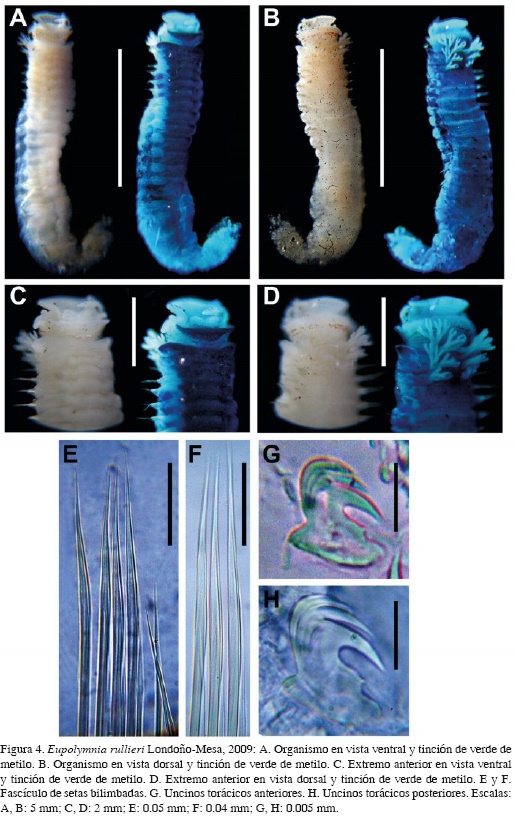
Material examinado. CEMUA POLY-TERE 0004 (1) Playa Mazatlán, 27.II.2011.
Descripción. Organismo incompleto (Figura 4A y B), en regular estado de preservación. Tórax con 6.0 mm de largo y 1.1 mm de ancho. Labio superior tan amplio como el inferior, redondeado, liso, con borde hinchado (Figura 4C). Labio inferior hinchado lateralmente. Membrana tentacular lisa y circular en su base; con una fila de ocelos distribuidos al azar, ubicados en el medio del dorso con una discontinuidad (Figura 4D). Branquias arborescentes, con pedúnculos cortos y gruesos. Alas laterales cortas, hinchadas en los segmentos tres y cuatro. Dieciséis escudos ventrales, crecientes en longitud pero decrecientes en ancho. Papilas nefridiales no visibles. Notópodos cilíndricos, notosetas bilimbadas asimétricas y estriadas hasta la su parte más distal (Figura 4E y F). Neurópodos torácicos planos, anchos y gruesos; uncinos aviculares torácicos (Figura 4G y H) con MF: 2:3:1; PP levemente pronunciado; PF ausente; Oc cóncavo; Cp con cuatro líneas de dientes; USr recto; SrP ausente; LSr corto y conexo, terminando en AP redondeado; AF ausente; Bs alargada y convexa anteriormente. Uncinos abdominales similares. Pigidio no observado.
Patrón de tinción. Labio inferior, escudos ventrales y alas laterales del segmento tres tiñen profundamente. Los primeros seis segmentos tiñen dorsalmente. El resto del dorso sin tinción.
Discusión. La principal característica de esta especie es la presencia de ocelos alrededor de la membrana tentacular, con una interrupción dorsal, formando dos grupos de ocelos laterales. Eupolymnia nebulosa (Montagu, 1818) fue ampliamente registrada para el Caribe, pero dichos registros han sido dudosos debido a que fue originalmente descrita para Inglaterra. Londoño-Mesa (2009b) diferenció E. rullieri y E. nebulosa comparando material de la localidad tipo, y clarificó la identificación de E. magnifica (Webster, 1884), considerando que E. crassicornis no posee información suficiente para ser válida. Este estudio amplía la distribución de E. rullieri para las costas continentales del Caribe colombiano. De acuerdo con Londoño-Mesa (2011), sólo se tiene un registro para isla Providencia, identificado como Loimia medusa (Savigny in Lamarck, 1818).
Localidad tipo. Majahual, sur del Caribe mexicano.
Distribución. Golfo de México y Gran Caribe. Desde intermareal hasta 183 m de profundidad, en sustrato rocoso.
Lanicola Hartmann-Schröder, 1986
Lanicola Hartmann-Schröder, 1986: 58. Hutchings y Glasby 1988: 24. Capa y Hutchings 2006: 14. Londoño-Mesa 2009b: 34. Nogueira et al. 2010: 15.
Especie tipo. Lanicola lobata Hartmann-Schröder, 1986, por designación original.
Diagnosis. Membrana tentacular con o sin ocelos. Un par de alas laterales amplias y desarrolladas entre el segmento dos (latero-ventral) y tres (solo lateral). Dos pares de branquias ramificadas dicotómicas decrecientes en tamaño, en segmentos dos y tres. Escudos ventrales desde segmento dos. Papilas nefridiales en segmentos dos al cuatro o seis y siete. Veinte segmentos torácicos; 17 pares de notópodos desde segmento cuatro; notosetas lisas simétricamente bilimbadas. Neurópodos desde segmento cinco; torácicos con torus anchos y numerables uncinos; abdominales con torus cortos y neurópodos largos. Uncinos aviculares, en fila simple en segmentos 5-10, y en fila doble en segmentos 11-20, arreglados capitum frente a capitum; uncinos abdominales en fila simple hasta el extremo final. Pigidio liso (Londoño-Mesa, 2009b).
Lanicola carus (Young y Kritzler, 1987) Figura 5A-J
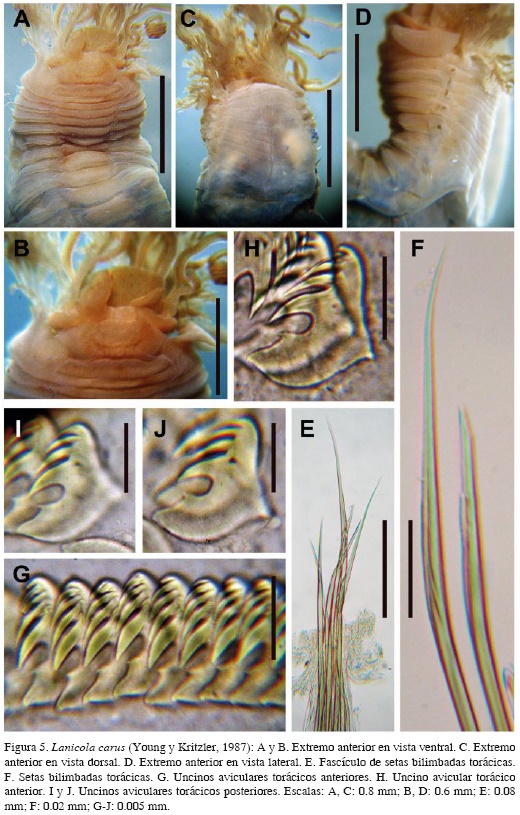
Paraeupolymnia carus Young y Kritzler, 1987: 687-689, Fig. 1. Londoño-Mesa 2006: 24-30, Fig. 1 A-K.
Pista palmata. Rodríguez-Gómez 1988: 417-418. Dueñas 1999: 14.
Lanicola carus. Capa y Hutchings, 2006: 14. Londoño-Mesa 2009b: 34-35; Londoño-Mesa 2011: 21-22. Nogueira et al. 2010: 144.
Material examinado. CEMUA POLY-TERE 0005 (1) Playa El Matal, 08.VIII.2009. POLY-TERE 0006 (1) Sin datos, 08.VIII.2009. POLY-TERE 0007 (1) Playa San Diego, 27.II.2011.
Descripción. Organismo incompleto (POLY-TERE 0005). Tórax con 9.0 mm de largo y 2.0 mm de ancho. Labio superior más amplio que el inferior, redondeado, liso, con borde hinchado. Labio inferior abultado lateralmente (Figura 5A y B). Membrana tentacular lisa y circular en su base, con cuatro filas de ocelos fusionadas dorsalmente formando un cúmulo de ocelos de color café oscuro. Branquias ramificadas con pedúnculos cortos y gruesos (Figura 5C). Alas laterales bien desarrolladas, amplias, de borde delgado y liso, ubicadas lateralmente (Figura 5D). Dieciséis escudos ventrales, crecientes en longitud pero decrecientes en ancho. Papilas nefridiales dorsales y posteriores a los notópodos, en segmentos cuatro, seis y siete. Notópodos cilíndricos, notosetas bilimbadas asimétricas y estriadas hasta la punta (Figura 5E y F). Neurópodos torácicos planos y gruesos, con uncinos aviculares torácicos (Figura 5G-J) MF: 2:3; PP levemente pronunciado; PF ausentes; Oc cóncavo; Cp con dos líneas de dientes; USr cóncavo; SrP redondeado; LSr corto, terminando en AP redondeado; AF ausente; Bs alargada y convexa. Uncinos abdominales similares. Pigidio no observado.
Patrón de tinción. Escudos ventrales, primer segmento y una banda debajo del borde del labio superior tiñeron profundamente.
Variaciones. Individuos adicionales (POLY-TERE 0006, POLY-TERE 0007) completos, con 61-72 segmentos y entre 6-39 mm de largo. El tórax varió entre 2.2-10 mm de largo y entre 0.7-3.0 mm de ancho. Presentaron entre 12-16 escudos ventrales. Papilas nefridiales en los segmentos seis y siete en el individuo más grande (POLY-TERE 0006); (POLY-TERE 0007), al parecer, sin estado reproductivo, ya que no presentó papilas.
Discusión. Esta especie fue registrada por Londoño-Mesa (2011) para isla Tintipán, por lo que se amplia la distribución hacia el suroeste del Caribe colombiano. Lanicola es un género que puede llegar a confundirse con otros como Terebella y Pista. Sin embargo, Terebella no presenta alas laterales y las notosetas son serradas a diferencia de Lanicola que tiene un par de alas laterales y setas con punta lisa. Por otro lado, Pista presenta dos o más pares de alas laterales, el cuerpo muestra una consistencia más rígida, aún en fijación, y tiene uncinos con proceso posterior (PP) largo, al menos, en el primer segmento torácico. También presenta branquias más ramificadas que Lanicola. En Pista es común encontrar este caracter con tamaños diferentes, debido a que se desprenden con gran facilidad y consecutivamente son regeneradas por el organismo. Finalmente, L. carus se diferencia de otra especie del género registrada en el Gran Caribe, L. garciagomezi (Londoño-Mesa, 2006), por la cantidad de tubérculos que presenta en la región anterior dorsal, así como por la presencia de ocelos, características ausentes en L. garciagomezi.
Localidad tipo. Twin Kays, Belice, Caribe. En aguas someras.
Distribución. Londoño-Mesa (2009b) considera esta especie común para el Gran Caribe, en Florida, golfo de México, Caribe mexicano, Belice, Caribe panameño y colombiano, Venezuela y Antillas Menores (Bonaire, Curaçao). Igualmente, se registró en Balboa (golfo de Panamá, Pacífico). Se encuentra generalmente en aguas someras, entre 0-5 m de profundidad, aunque hay registros a 237 m. En el Caribe colombiano, esta especie se presenta en el Parque Nacional Natural Tayrona, Santa Marta, isla Tintipán e Isla Fuerte, como localidad más al sur.
Loimia Malmgren, 1866
Loimia Malmgren, 1866: 380. Hessle 1917: 170. Hutchings y Glasby 1995: 149-150. Londoño-Mesa 2009b: 35-36. Nogueira et al. 2010: 17, 28, 30.
Especie tipo. Terebella medusa Savigny, 1818, por monotipia.
Diagnosis. Membrana tentacular con o sin ocelos. Alas laterales en segmentos uno y tres o dos y tres. Tres pares de branquias ramificadas dicotómicas, en segmentos 2-4. Escudos ventrales desde el segmento dos o tres. Pares de papilas nefridiales en segmentos tres y cuatro y seis, siete y ocho. Veinte segmentos torácicos; 17 pares de notópodos desde el segmento cuatro; notosetas lisas asimétricamente bilimbadas. Neurópodos desde el segmento cinco; torácicos con torus anchos y numerables uncinos; abdominales con torus cortos y neurópodos largos. Uncinos aviculares o pectinados, con una serie vertical de dientes, en fila simple en segmentos cinco al 10; en fila doble en segmentos 11-20, arreglados occipitium contra occipitium; uncinos abdominales en fila simple hasta el extremo final. Pigidio con cirros en algunas especies (Londoño-Mesa, 2009b).
Loimia minuta Treadwell, 1929
Loimia minuta Treadwell, 1929: 10-11, Figs. 28-30. Londoño-Mesa y Carrera-Parra 2005: 21-22, Fig. 5 H-N. Londoño-Mesa 2009b: 39, Fig. 10 A-J.
Material examinado. CEMUA POLY-TERE 0008 (1), sin datos, 08.VIII.2009.
Descripción. Individuo incompleto y en mal estado. Tórax con 5.0 mm de largo y 1.5 mm de ancho. Labio superior amplio, ovalado, con borde liso, delgado, proyectado al frente. Labio inferior pequeño e hinchado. Membrana tentacular aplanada dorsoventralmente, corta y estriada longitudinalmente. Ocelos ausentes. Branquias arborescentes, decrecientes en tamaño, con pedúnculo grueso y corto. Primer par de alas laterales en segmento uno, amplias, proyectadas anteroventralmente, rodeando la membrana tentacular; segundo par de alas en segmento tres, cortas, redondeadas, ubicadas lateralmente, con base corta. Doce escudos ventrales desde el segmento dos; primeros cuatro alargados y rectangulares, los siguientes decreciendo en longitud y crecientes en ancho. Ocho segmentos gruesos, más anchos que largos, luego de los escudos, llegando hasta el abdomen. Papilas nefridiales no visibles. Notópodos cilíndricos, notosetas de dos tamaños, con bases estriadas. Neurópodos torácicos planos y gruesos, con uncinos de los segmentos cinco al 10 aviculares, MF: 1:1:1:1; PP y PF ausentes; Oc alargado, cóncavo; Cp con un sólo diente en cada una de las cuatro filas; el primer diente mide ¾ del MF; USr cóncavo; LSr ovalado y pronunciado en la parte superior; SrP pronunciado; AP desarrollado, ovalado; AF delgado; Bs alargada, ligeramente cóncava. Uncinos de los segmento 11-20 aviculares, MF: 1:1:1:1; Cp con cuatro dientes accesorios. Uncinos abdominales más pequeños, MF: 1:1:1:1:1; Cp con cinco dientes accesorios. Pigidio no observado.
Patrón de tinción. Escudos ventrales tiñen profundamente. Primer par de alas tiñen en la base más no en el borde. Segundo par de alas, segundo segmento y base de los primeros cinco notópodos tiñeron profundamente.
Discusión. Esta especie fue registrada para el Gran Caribe por Londoño-Mesa (2009b); sin embargo, es el primer registro para el Caribe colombiano. Loimia minuta se diferencia de otras especies de género en el Caribe por tener uncinos torácicos con cuatro dientes y uncinos abdonimales con cinco dientes. Loimia bermudensis Verrill, 1900, posee uncinos torácicos y abdominales con cinco dientes, y Loimia salazari Londoño-Mesa y Carrera-Parra, 2005 tiene uncinos torácicos y abdominales con tres y cuatro dientes, respectivamente. Londoño-Mesa (2009b) registró un especimen con 11 mm de largo, posee un número mayor de dientes tanto en el toráx como en el abdomen, comparado con este material, el cual solo mide 5 mm de largo. Esta diferencia puede deberse al tamaño de los ejemplares estudiados; los individuos de menor talla pueden llegar a tener un menor número de dientes. No obstante, hace falta corroborar este hecho con la revisión de un mayor número de ejemplares del Caribe colombiano.
Localidad Tipo. Dry Tortugas, Florida. En aguas intermareales.
Distribución. Florida, golfo de México, Caribe mexicano y colombiano.
Pista Malmgren, 1866
Pista Malmgren, 1866: 382. Hessle 1917: 153, Text. Fig. 34. Saphronova 1991: 243-244. Hartmann-Schröder 1996: 519. Hilbig 2000: 267. Londoño-Mesa 2009b: 45-47; Londoño-Mesa 2012: 40. Nogueira et al. 2010: 17, 23, 29, Fig. 12 D-H.
Especie tipo. Amphitrite cristata Müller, 1776, por designación original.
Diagnosis. Membrana tentacular con o sin ocelos. Tres a cuatro pares de alas laterales en segmentos uno al cuatro. Uno o dos pares de branquias ramificadas espiraladas, en segmento dos, o en segmentos dos y tres, fácilmente desprendibles (dehiscentes), a veces faltando. Escudos ventrales desde segmento dos o tres. Papilas nefridiales en segmentos seis y siete. Veinte segmentos torácicos; 17 pares de notópodos desde segmento cuatro; notosetas lisas asimétricamente bilimbadas. Neurópodos desde segmento cinco; torácicos con torus anchos y numerables uncinos; abdominales con torus cortos y neurópodos largos. Uncinos aviculares, en fila simple en segmentos cinco al 10, y en fila doble en segmentos 11-20, arreglados capitum frente a capitum; uncinos de, al menos, un neurópodo torácico anterior con PP largo. Pigidio liso o con cirros (Londoño-Mesa, 2009b).
Pista cetrata (Ehlers, 1887)
Figura 6 A-J
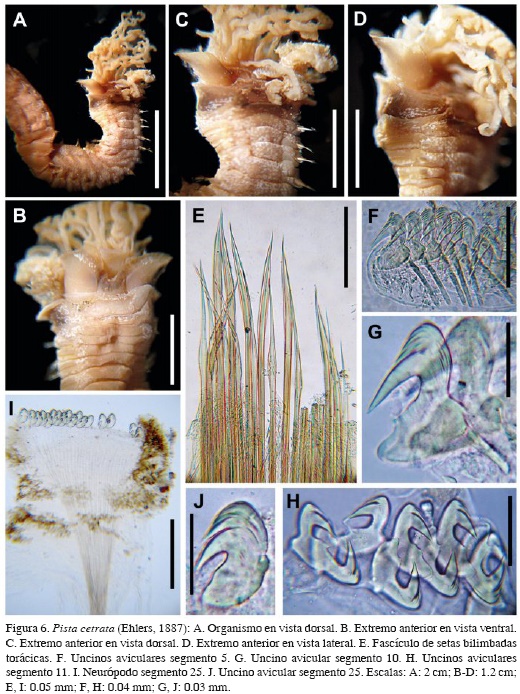
Terebella cetrata Ehlers, 1887: 248-253, Lám. 52, Fig. 8-13. Hessle 1917: 163. Hartman 1938: 18.
Nicolea cetrata. Hartman 1959: 512. Perkins y Savage 1975: 54.
Holthe 1986: 138. Salazar-Vallejo 1996: 32.
Pista cetrata. Londoño-Mesa, 2009b: 47-50, Fig. 13 A-N.
Material examinado. CEMUA POLY-TERE 0009 (1) Playa La Cebolleta, 04.IX.2010.
Descripción. Organismo incompleto (Figura 6A), 20 mm de largo y 2.5 mm de ancho, 37 segmentos. Tórax 12 mm de largo. Dorso con puntos blancos sin un patrón de distribución. Labio superior delgado, membranoso, plegado lateralmente en forma de "M" (Figura 6B), con lóbulos laterales redondeados, más largo que el ala lateral del segmento uno. Labio inferior corto, escasamente visible. Membrana tentacular reducida, cubierta por las alas laterales del segmento uno. Ocelos ausentes. Cuatro pares de alas laterales (Figura 6C y D); primer par en segmento uno, bien desarrolladas, proyectadas anteriormente, redondeadas lateralmente, borde liso y más claro, ventralmente unidas por un tejido más corto y más oscuro, en forma de "U" que cubre al labio inferior. Segundo par en segmento dos, más corto y sólo visible ventro-lateralmente: ventralmente unidas al primer escudo ventral. Tercer par en segmento tres, desarrolladas latero-dorsalmente, más oscuras, decreciendo en tamaño hacia el dorso, con un bolsillo interno oscuro, característico de la especie. Dos pares de branquias, en segmentos dos y tres, decrecientes en tamaño, con pedúnculos gruesos y largos, ramificadas sin un patrón definido. Papilas nefridiales levemente visibles, en segmentos seis y siete, rodeadas por un tejido notopodial glandular pálido. Dieciséis escudos ventrales desde el segmento dos; primer escudo más largo y ancho, el doble del segundo; los siguientes cuatro más cortos; los medios y posteriores con forma rectangular. Notosetas de dos tamaños (Figura 6E), bien desarrolladas: largas gruesas, asimétricamente bilimbadas, con limbo finalizando cerca al extremo distal; y cortas delgadas y simétricamente bilimbadas medialmente. Uncinos aviculares, torácicos de los segmentos 5-10 (Figura 6F y G) MF: 3:8:3; PP desarrollado; PF delgado y largo; Oc levemente cóncavo y largo; Cp con dientes de la segunda fila alcanzando la base del MF; MF grande, largo y puntiagudo; USr cóncavo; SrP en ángulo recto; SrA ausente; LSr corto y cóncavo; AP redondeado; AF corto y dirigido hacia la Bs; Bs levemente recta. Uncinos de los segmento 11-20 (Figura 6H) MF: 3:1; PP desarrollado; PF delgado y corto; Oc levemente cóncavo, largo y delgado; Cp con dientes de la segunda fila alcanzando la base del MF; MF grueso, largo y puntiagudo; USr cóncavo; SrP corto, en ángulo recto; SrA ausente; LSr largo y levemente cóncavo; AP redondeado; AF corto y dirigido hacia la Bs; Bs cóncava. Uncinos abdominales más pequeños (Figura 6I y J) MF: 2:3; PP corto; PF ausente; Oc cóncavo y muy corto; Cp con dientes de la segunda fila alcanzando más de la mitad del MF; MF grueso y corto; USr escasamente cóncavo; SrP escasamente visible; SrA ausente; LSr largo y aplanado; AP redondeado; AF ausente; Bs extremadamente cóncava. Pigidio no observado.
Discusión. Este es el primer registro de P. cetrata en el Caribe colombiano. Las claves taxonómicas antes de Londoño-Mesa (2009b) incluían esta especie como Nicolea cetrata. No obstante, la especie fue redescrita y corregida como combinación nueva, debido a que las características morfológicas demostraban que pertenece a Pista. Por otro lado, esta especie es similar a P. palmata (Verrill, 1873), pues ambas presentan dos lóbulos en el borde dorsal del segmento cuatro. Sin embargo, los lóbulos en P. palmata son más largos y cercanos, mientras que aquellos en P. cetrata son más pequeños y separados entre ellos. Posiblemente, la especie haya sido recolectada en Colombia, pero aún permanezca sin identificar en materiales de colecciones nacionales. Londoño-Mesa (2011) no registró la especie en material identificado previamente en el Caribe colombiano.
Localidad tipo. Key West, Florida, Estados Unidos.
Distribución. Florida, Aruba, Curaçao, Tobago e Isla Fuerte (Caribe colombiano). Intermareal hasta 205 m de profundidad.
Pista palmata (Verrill, 1873)
Scionopsis palmata Verrill, 1873: 320.
Pista palmata. Hartman 1951: 112. Renaud 1956: 34. Day 1973: 119120. Kritzler 1984: 52-51, Figs. 52.47, 52.48 A-F. Báez y Ardila 2003: 106. Londoño-Mesa y Carrera-Parra 2005: 28-29, Fig. 7 E-J. Londoño-Mesa 2009b: 50-52, Fig. 14 A-L; 2011: 24-25.
Eupolymnia crassicornis. Dueñas 1999: 14.
Loimia medusa. Dueñas 1999: 14.
Terebella rubra. Dueñas 1999: 14.
Material examinado. CEMUA POLY-TERE 00010 (5) Playa El Matal, 07.VIII.2009. POLY-TERE 0011 (1) Sin datos, 09.VIII.2009. POLY-TERE 0012 (16) Playa El Latal, 05.VIII.2010.
Descripción. Organismo completo (POLY-TERE 0010), 35 mm de largo y 2 mm de ancho, 88 segmentos. Tórax 9 mm de largo. Labio superior ancho, más corto en la base y más ancho en el borde, sobresale por encima de las alas del primer segmento, borde liso con ondulaciones. Labio inferior corto, con borde hinchado, de forma rectangular, cubierto por las alas del primer segmento. Membrana tentaculargh reducida, con borde hinchado. Tres pares de alas laterales; primer par en segmento uno, bien desarrolladas, amplias, redondeadas, borde liso, unidas dorsalmente; segundo par en segmento tres, más corto que el primer par, con borde ligeramente hinchado en los laterales; tercer par en segmento cuatro, poco desarrolladas, cortas y laterales, delgadas y unidas al par de lóbulos dorsales del segmento cuatro. Branquias en segmentos dos y tres, desiguales en tamaño, con pedúnculos gruesos y largos. Papilas nefridiales ausentes. Trece escudos ventrales; primer escudo ocupando la mitad inferior del segmento, los dos siguientes ocupan el segmento completo; el resto decreciendo en tamaño, más anchos que largos, con forma rectangular. Notosetas bien desarrolladas, bilimbadas simétricas, con limbo finalizando cerca al extremo distal. Uncinos aviculares, torácicos MF: 3:5:α, PP desarrollado, PF alargado, Oc cóncavo corto, Cp con dientes de la segunda fila alcanzando la mitad del MF, MF grande y grueso, USr cóncavo, SrP termina leventemente en punta, SrA ausente, LSr cóncavo, AP estriado con punta redonda, AF ausente, Bs levemente recta; uncinos de los segmento 11 al 20 sin PP. Pigidio con papilas.
Patrón de tinción. El borde de la membrana tentacular, la parte dorsal del primer par de alas, (menos su borde) y el segundo par de alas laterales tiñen profundamente, al igual que, los 13 escudos ventrales, la base de los notópodos y dorsalmente los segmentos uno, dos y tres.
Variaciones. Individuos completos e incompletos (POLY-TERE 00010, POLY-TERE 0011, POLY-TERE 0012), con 56-94 segmentos, 20-49 mm de largo y 1-2 mm de ancho. Tórax con 1-15 mm de largo y 1-2 mm de ancho. Los escudos ventrales pueden variar desde 12-14. Algunos presentan dos pares de papilas nefridiales en los segmentos seis y siete. Todos con uncinos con PP desarrollado en los primeros segmentos y MF: 2:2-3:1-∞.
Discusión. Pista palmata se diferencia claramente de las otras especies del género presentes en la región por poseer todos los uncinos con procesos posteriores largos; tener tres pares de alas laterales y un par de lóbulos digitiformes separados en el borde dorsal del segmento cuatro. Pista cetrata tiene cuatro pares de alas y un par de lóbulos más pequeños y separados. Las branquias de P. palmata se presentan en forma de hoja de palma, mientras que las de Pista sp., descrita más adelante, son arborescentes dicotómicas. Este registro amplía la distribución de la especie hacia el sur del Caribe colombiano.
Localidad tipo. Nueva Inglaterra, Gran Egg Harbor hasta New Haven y Vineyard Sound, Estados Unidos.
Distribución. Connecticut, Rhode Island, Massachusetts, Florida, golfo de México, Caribe mexicano, panameño y colombiano. Desde la zona intermareal hasta 243 m de profundidad.
Pista sp.
Material examinado. CEMUA POLY-TERE 0013 (1) Sin datos, 08.VIII.2009.
Descripción. Organismo incompleto, 32 segmentos. Tórax 3 mm de largo y 1.8 mm de ancho. Labio superior pronunciado, redondeado, liso y con borde delgado. Labio inferior grueso con borde hinchado. Membrana tentacular lisa, circular, con borde grueso. Ocelos ausentes. Dos pares de alas laterales; primer par amplio, borde grueso en segmento uno, las alas termina en la base del labio inferior. Segundo par entre los segmentos dos y tres, más pequeñas, ovaladas y bien desarrolladas. Lóbulo dorsal recto. Dos pares de branquias decrecientes en tamaño en los segmentos dos y tres, con pedúnculo largo y ligeramente delgado. Papilas nefridiales no visibles. Doce escudos ventrales desde segmento dos, decrecientes en tamaño; últimos cuatro centrados en el segmento. Notosetas de dos tamaños, lanceoladas, bilimbadas asimétricamente. Uncinos aviculares, torácicos de los segmentos cinco al 10 MF: 3:4-5, con PP largo desde los segmentos cinco al nueve, sin PF, Oc ligeramente cóncavo, casi recto; Cp con tres filas de dientes, la segunda línea llega a ¼ del MF; USr ovalado, muy pronunciado; SrP más cercano al AP que al USr; AP marcado; ausencia de AF; Bs cóncavo y estriado. Uncinos de los segmento 11-20 MF: 3:2; PP desarrollado; PF delgado y corto; Oc levemente cóncavo, largo y delgado; Cp con dientes de la segunda fila alcanzando la base del MF; MF grueso, largo y puntiagudo; USr cóncavo; SrP corto, en ángulo recto; SrA ausente; LSr largo y levemente cóncavo; AP redondeado; AF corto y dirigido hacia la Bs; Bs cóncava. Uncinos abdominales más pequeños, MF: 2:2-3; PP corto; PF ausente; Oc cóncavo y muy corto; Cp con dientes de la segunda fila alcanzando más de la mitad del MF; MF grueso y corto; USr escasamente cóncavo; SrP escasamente visible; SrA ausente; LSr largo y aplanado; AP redondeado; AF ausente; Bs extremadamente cóncava. Pigidio no observado.
Patrón de tinción. Los escudos ventrales, los dos primeros segmentos, las alas laterales y el borde dorsal de los notópodos tiñen intensamente.
Discusión. Pista sp. se diferencia de otras especies del género registradas para el Gran Caribe por Londoño-Mesa (2009b) ya que posee solo dos pares de alas laterales, a diferencia de P. palmata que tiene tres y P. cetrata que tiene cuatro pares. Las branquias de Pista sp. son similares a las de P. cetrata, decreciendo en tamaño y con un pedúnculo largo y delgado, pero son diferentes a las de P. palmata pues ésta presenta tamaños diferentes entre las branquias del mismo segmento y el pedúnculo es corto y grueso. Igualmente, Pista sp. posee un lóbulo dorsal recto, a diferencia de P. palmata que posee dos lóbulos dorsales largos y P. cetrata que tiene dos lóbulos cortos separados por un borde cóncavo amplio.
Distribución. Isla Fuerte, Caribe colombiano.
Terebella Linnaeus, 1767
Terebella Linnaeus, 1767: 1092. Hessle 1917: 187-188, Figura 45. Day 1967: 747. Holthe 1986: 93. Capa y Hutchings 2006: 20. Londoño-Mesa 2009b: 63.
Especie tipo. Terebella lapidaria Linnaeus, 1767, por monotipia.
Diagnosis. Membrana tentacular con o sin ocelos. Dos o tres pares de branquias en los segmentos dos y tres o dos a cuatro. Escudos ventrales desde segmento dos. Papilas nefridiales en segmentos tres y cuatro y en números variables desde el segmento cinco o seis. Veinte segmentos torácicos; diecisiete o más pares de notópodos desde el segmento cuatro; notosetas bilimbadas en la base y aserradas distalmente. Neurópodos desde segmento cinco; torácicos con torus anchos y numerables uncinos; abdominales con torus cortos y neurópodos largos. Uncinos aviculares, en fila simple en segmentos cinco al 10, y en fila doble en segmentos 11-20, arreglados capitum frente a capitum hasta el segmento 20. Pigidio liso o con cirros.
Terebella verrilli Holthe, 1986
Terebella rubra. Hartman 1945: 44; 1951: 112. Day 1973: 119.
Kritzler 1984: 52-39, 52-40, Fig. 52-35, 36 A-E. Dueñas 1999: 14. Terebella verrilli Holthe, 1986: 151. Londoño-Mesa y Carrera-Parra 2005: 33-34, Fig. 8 F-K. Londoño-Mesa 2009b: 64-65; Londoño-Mesa 2011: 27.
Material examinado. CEMUA POLY-TERE 0014 (5) Playa San Diego, 27.II.2011. POLY-TERE 0015 (1) Playa El Matal, IX.2011.
Descripción. Organismo incompleto (POLY-TERE 0014), 30 segmentos. Tórax 4.5 mm de largo y 1.0 mm de ancho. Labio superior delgado, ovalado, con borde delgado. Labio inferior grueso y ovalado. Membrana tentacular lisa, corta, con borde hinchado; base con fila de ocelos que dorsalmente hacen una línea y ventralmente forman un cúmulo. Tres pares de branquias en segmentos dos al cuatro, arborescentes, no dicotómicas, decrecientes en tamaño y con pedúnculos cortos. Sin alas laterales. Catorce escudos ventrales decrecientes en tamaño. Un par de papilas nefridiales, dorsales a las branquias del segundo segmento. Más de 20 notópodos cilíndricos desde el segmento cinco. Notosetas de dos tamaños; setas largas serradas distalmente hasta la punta; setas cortas con dientes con la serración más largos. Uncinos aviculares, torácicos MF: 3:2:1-2; PP y PF ausentes; Oc cóncavo y corto; Cp con dientes de la segunda fila alcanzando la base del MF; MF grande, largo y puntiagudo; Sr recto; USr, SrP, SrA y LSr ausentes; AP redondeado; AF ausente; Bs convexa. Uncinos abdominales con MF: 3:2, más pequeños. Pigidio no observado.
Patrón de tinción. Los escudos ventrales, los segmentos dos y tres dorsalmente, algunas papilas del dorso, los tentáculos y el tejido notopodial glandular, tiñen intensamente.
Variaciones. Individuos incompletos (POLY-TERE 0014, POLY-TERE 0015), tórax mide entre 1.2-3.0 mm de largo y 1 mm de ancho. Poseen entre 12-14 escudos ventrales. Papilas nefridiales se ubican entre las branquias del segmento dos y tres y seis y siete o no logran observarse.
Discusión. Terebella verrilli se diferencia de la otra especie registrada para el Caribe, T. turgidula Ehlers, 1887, básicamente por la presencia de ocelos, carácter faltante en la segunda especie. Por otro lado, uno de los ejemplares observados en esta investigación (POLY-TERE 0014), demuestra que el segundo par de branquias se presentan en la intersección de los segmentos dos y tres, por lo que aparenta tener dos pares de branquias en el segmento dos y entre ellas las papilas nefridiales. Al comparar esta condición con lo descrito por Londoño-Mesa (2009b), se muestra una diferencia que podría ser aclarada con futuros estudios histológicos para hallar la verdadera inserción de las branquias.
Localidad tipo. Esta especie fue descrita para Vineyard Sound, Massachusetts.
Distribución. Massachusetts, Florida, golfo de México, Caribe mexicano, panameño y colombiano y Antillas Menores. Este estudio extiende la distribución de la especie hasta el litoral medio de la costa del Caribe colombiano.
AGRADECIMIENTOS
Los autores agradecen al Comité para el Desarrollo de la Investigación, CODI, Universidad de Antioquia, por proveer los recursos económicos, mediante el Acta número TE20100 de apoyo a trabajos de grado. Igualmente, se agradece a Leris Zúñiga, Corregidora de Isla Fuerte, por el apoyo durante la estancia en la Isla. Agradecimientos especiales a Leiver Galvis "Niño" por su apoyo en la logística para alcanzar todos los sitios de muestreo, así como a los estudiantes del curso de Zoología de Invertebrados, pregrado en Biología, Universidad de Antioquia, semestres 2009, 2010 y 2011 por su apoyo en la separación de muestras durante las recolectas en la isla. Finalmente, a los evaluadores y al Comité Editorial por sus valiosos aportes.
BIBLIOGRAFÍA
Augener, H. 1925. Über westindische und einige andere Polychaeten-typen von Grube (Oersted), Krøyer, Morch und Schmarda. Publ. Univ. Zool. Mus., København, 39: 1-47. [ Links ]
Báez, D. y N. Ardila. 2003. Poliquetos (Annelida: Polychaeta) del mar Caribe colombiano. Biota Col., 4(1): 89-109. [ Links ]
Capa, M. y P. Hutchings. 2006. Terebellidae (Polychaeta) from the Pacific coast of Panama (Coiba National Park) including descriptions of four new species and synonomy of the genus Paraeupolymnia with Lanicola. Zootaxa, 1375: 1-29. [ Links ]
Day, J.H. 1967. A monograph on the Polychaeta of Southern Africa. Brit. Mus. Nat. Hist. Publ., 656: 1-878. [ Links ]
Day, J.H. 1973. New Polychaeta from Beaufort, with a key to all species recorded from North Carolina. NOAA Tech. Rep. NMFS Circ., 375: 1-153. [ Links ]
Dueñas, P.R. 1999. Algunos poliquetos (Annelida) del Caribe colombiano. Milenio, 1(2): 9-18. [ Links ]
Ehlers, E. 1887. Reports on the results of dredging under the supervision of Alexander Agassiz, in the Gulf of Mexico (1877-78), and in the Caribbean Sea (1878-79), in the United States Coast Survey Steamer "Blake", Lieutenant-Commander C.D. Sigsbee, U.S.N., Commanding. Preliminary report on the worms. Bull. Mus. Comp. Zool., 15: 237-253. [ Links ]
Estela, F.A. 2006. Aves de Isla Fuerte y Tortuguilla, dos islas de la plataforma continental de Caribe colombiano. Bol. Inst. Inv. Mar. Cost., 35: 267-272. [ Links ]
Grube, A.E. 1850. Die Familien der Anneliden. Arch. Naturgesch., 16: 249-364. [ Links ]
Hartman, O. 1938. Annotated list of the types of polychaetous annelids in the Museum of Comparative Zoology. Bull. Mus. Comp. Zool., 85(1): 1-31. [ Links ]
Hartman, O. 1939. Polychaetous annelids collected by the Presidential Cruise of 1938. Smith. Miscel. Coll., 98: 1-22. [ Links ]
Hartman, O. 1945. The marine annelids of North Carolina. Duke Univ. Mar. Stat. Bull., 2: 1-54. [ Links ]
Hartman, O. 1951. The littoral marine annelids of the Gulf of Mexico. Publ. Inst. Mar. Sci. Port Aransas, Texas, 2(1): 7-124. [ Links ]
Hartman, O. 1959. Catalogue of the Polychaetous Annelids of the World. Part 2. Allan Hancock Found. Pub. Occas. Pap., 23(2): 355-628. [ Links ]
Hartmann-Schröder, G. 1986. Zur Kenntnis des Eulitorals der Australischen Kuüsten unter besonderer Beruücksichtigung der Polychaeten und Ostracoden, 12. Die Polychaeten der antiborealen Suüdkuüste Australiens (zwischen Wallaroo im Westen und Port Mac Donnell im Osten). Mitt. Hamb. Zool. Mus. Inst., 83: 31-70. [ Links ]
Hartmann-Schröder, G. 1996. Annelida, Borstenwuürmer, Polychaeta. Tierwelt Dtl, Jena, 58: 1-645. [ Links ]
Hessle, C. 1917. Zur Kenntnis der Terebellomorphen Polychaeten. Zool. Bidrag. Uppsala, 5: 39-258. [ Links ]
Hilbig, B. 2000. Family Terebellidae Grube, 1851. 231-293. En: Blake, J.A., B. Hilbig and P.V. Scott (Eds.). Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and the Western Santa Barbara Channel. The Annelida 4, Polychaeta: Flabelligeridae to Sternaspidae. Santa Barbara Museum of Natural History, Santa Barbara, EE.UU. 377 p. [ Links ]
Holthe, T. 1986. Evolution, systematics, and distribution of the Polychaeta Terebellomorpha, with a catalogue of the taxa and a bibliography. Gunneria, 55: 1-236. [ Links ]
Hutchings, P.A. y C.J. Glasby. 1988. The Amphitritinae (Polychaeta: Terebellidae) from Australia. Rec. Aust. Mus., 40: 1-60. [ Links ]
Hutchings, P.A. y C.J. Glasby. 1995. Description of the widely reported terebellid polychaetes Loimia medusa (Savigny) and Amphitrite rubra (Risso). Mitt. Hamb. Zool. Mus. Inst., 92: 149-155. [ Links ]
Kinberg, J.G.H. 1867. Annulata nova. Öfversigt af Kongl. Vetenskaps-Akademiens Förhandlingar, 23: 337-357. [ Links ]
Kritzler, H. 1984. Family Terebellidae Grube 1950. 53-124. En: Uebelacker, J.M. y P.G. Johnson (Eds.). Chapter 52. Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Barry A. Vittor and Associates Inc., Mobile, Estados Unidos. 228 p. [ Links ]
Linnaeus, C. 1767. Systema naturae sive regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Laurentii Salvii, Holmiae. 12(1): 533-1327. [ Links ]
Londoño-Mesa, M.H. 2006. Revision of Paraeupolymnia, and redescription of Nicolea uspiana comb. nov. (Terebellidae: Polychaeta). Zootaxa, 1117: 21-35. [ Links ]
Londoño-Mesa, M.H. 2009a. Terebellidae Grube, 1850. 641-655. En: de León, J.A., J.R. Bastida, L.F. Carrera, M.E. García, A. Peña, S.I. Salazar-Vallejo y V. Solis (Eds.). Poliquetos (Annelida: Polychaeta) de México y América Tropical. Universidad Autónoma de Nuevo León, Monterrey, México. 737 p. [ Links ]
Londoño-Mesa, M.H. 2009b. Terebellidae (Polychaeta: Terebellida) from the Grand Caribbean región. Zootaxa, 2320: 1-93. [ Links ]
Londoño-Mesa, M.H. 2011. Terebélidos (Terebellidae: Polychaeta: Annelida) del Caribe colombiano. Biota Col., 12(1): 17-34.
Londoño-Mesa, M.H. 2012. New species of Pista Malmgren, 1866 (Polychaeta: Terebellidae) from the Caribbean. Zootaxa, 3317: 39-48. [ Links ]
Londoño-Mesa, M.H. y L.F. Carrera-Parra. 2005. Terebellidae (Polychaeta) from the Mexican Caribbean with description of four new species. Zootaxa, 1057: 1-44. [ Links ]
Malmgren, A.J. 1866. Nordiska Hafs-Annulater. Öfversigt af Kongl. Vetensk. Akad. Handl. Stockholm, 5: 355-410. [ Links ]
Muüller, O.F. 1776. Zoologica Danicae. Prodromus seu Animalium Daniae et Norvegiae indigenarum characters, nomine et synonyma imprimis popularium. Zool. Mus. Copenhagen. 274 p. [ Links ]
Nogueira, J.M.M., P. Hutchings y M.V. Fukuda. 2010. Morphology of terebelliform polychaetes (Annelida: Polychaeta: Terebelliformia), with a focus on Terebellidae. Zootaxa, 2460: 1-185. [ Links ]
Nogueira, J.M.M., L. Harris, P. Hutchings y M.V. Fukuda. 2011. Four terebellines (Polychaeta, Terebellidae) with problematic taxonomic histories. Zootaxa, 2995: 1-26. [ Links ]
Perkins, T.H. y T. Savage, 1975. A bibliography and checklist of polychaetous annelids of Florida, the Gulf of Mexico, and the Caribbean region. Florida Mar. Res. Pub., 14: 1-62. [ Links ]
Renaud, J.C. 1956. A report on some polychaetous annelids from the Miami-Bimini area. Amer. Mus. Novitates, 1812: 1-39. [ Links ]
Rodríguez-Gómez, H. 1988. Contribución al conocimiento de los anélidos (Annelida: Polychaeta) de aguas someras en la bahía de Nenguange Parque Nacional Natural Tayrona, Colombia. Trianea, 2: 403-443. [ Links ]
Salazar-Vallejo, S.I. 1996. Lista de especies y bibliografía de poliquetos (Polychaeta) del Gran Caribe. An. Inst. Biol. Univ. Nal. Auton. Mex., Ser. Zool., 67(1): 11-50. [ Links ]
Saphronova, M.A. 1991. Redescription of some species of Scionella Moore 1903, with a review of the genus and comments on some species of Pista Malmgren 1866 (Polychaeta: Terebellidae). Ophelia Supplements, 5: 239-247. [ Links ]
Savigny, J.S. 1818. Section on Annelida. En Lamarck, J.B., Histoire naturelle des animaux sans vertèbres, vol. 5. Déterville & Verdière, París, 618 p. [ Links ]
Schmarda, L.K. 1861. Neue wirbellose Thiere beobachtet und gesammelt auf einer Reise um die Erdr 1853 bis 1857. 1. Turbellarien, Rotatorien und Anneliden, pt. 2. Wilhelm Engelmann, Leipzig, 164 p. [ Links ]
Treadwell, A.L. 1929. New species of polychaetous annelids in the collections of the American Museum of Natural History, from Porto Rico, Florida, Lower California, and British Somaliland. Amer. Mus. Novitates, 392: 1-13. [ Links ]
Verrill, A.E. 1873. Report upon the invertebrate animals of Vineyard Sound and the adjacent waters, with and account of the physical characters of the region. Bull. US. Fish. Comm., Rep. 1871-1872: 295-778. [ Links ]
Verrill, A.E. 1900. Additions to the Turbellaria, Nemertina, and Annelida of the Bermudas, with a revision of the New England genera and species. Trans. Conn. Acad. Arts Sci., 10, 595-671. [ Links ]
Young, M.W. y H. Kritzler. 1987. Paraeupolymnia, a new genus of terebellid (Polychaeta: Terebellidae) from Belize. Publ. Inst. Mar. Sci. Port Aransas., 100: 687-690. [ Links ]
RECIBIDO: 06/12/12 ACEPTADO: 19/11/2014
