










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.44 no.2 Santa Marta July/Dec. 2015
COMUNIDAD DE PECES ARRECIFALES EN RELACIÓN CON LA INVASIÓN DEL PEZ LEÓN: EL CASO DEL CARIBE SUR
REEF FISH COMMUNITY IN RELATION TO THE LIONFISH INVASION: THE SOUTHERN CARIBBEAN CASE
Víctor Coronado-Carrascal1, Rocío García-Urueña1 y Arturo Acero P.2
1 Universidad del Magdalena, Grupo de Investigación de Ecología y Diversidad de Algas Marinas y Arrecifes Coralinos, Santa Marta, Colombia. vicoronado85@hotmail.com, garciarociop@yahoo.com
2 Universidad Nacional de Colombia sede Caribe, Instituto de Estudios en Ciencias del Mar (Cecimar), Santa Marta, Colombia. aacerop@unal.edu.co
RESUMEN
Se evaluó la abundancia y talla de peces arrecifales en presencia del pez león (Pterois volitans) en las localidades de Morro de Santa Marta, Remanso, Isla Aguja y las bahías de Chengue y Cinto en el Parque Nacional Natural Tayrona (PNNT). Se tomó un grupo de especies de peces de importancia comercial (Lutjanus mahogoni, Cephalopholis cruentata y Haemulon plumierii) y otro de importancia de salud arrecifal (Abudefduf saxatilis, Chaetodon ocellatus, C. striatus, C. sedentarius y C. capistratus) y se evaluó su abundancia mensualmente durante 2012. El conteo se llevó a cabo a través de censos visuales en un transecto de 30 x 10 m. La mayor abundancia del pez león fue para Morro de Santa Marta (2.60 ± 2.12) y la menor para Remanso (0.58 ± 1.16); sin embargo, no hubo diferencia significativa entre localidades (p > 0.05). La mayor abundancia se registró en junio y julio y la menor en marzo, septiembre y octubre; para febrero no se registró pez león. La mayor abundancia de las especies de importancia comercial y de salud arrecifal se encontró en las localidades del PNNT. Los individuos más pequeños del pez león y de H. plumierii se registraron en Morro, Remanso e Isla Aguja. La talla promedio de las especies comerciales fue similar para todas las localidades, mientras que el pez león presentó una talla mayor. Esto puede indicar que las especies estudiadas no son de una talla suficiente para competir y por lo tanto podrían ser desplazadas, así mismo que la baja abundancia de las especies de peces arrecifales se deba a otros factores de estrés. Aun así, el invasor no ha alcanzado altas abundancias como en otras regiones de Caribe y puede que aún no haya un efecto negativo en la comunidad íctica.
PALABRAS CLAVES: Pterois volitans, Pez león, Invasión, Caribe colombiano.
ABSTRACT
The abundance and size of reef fishes in presence of lionfish were evaluated in Morro de Santa Marta, Remanso, and Isla Aguja and Chengue and Cinto Bays in the Parque Nacional Natural Tayrona (PNNT). Some commercially important fish species (Lutjanus mahogoni, Cephalopholis cruentata, and Haemulon plumierii) and some fish species important for reef health (Abudefduf saxatilis, Chaetodon ocellatus, C. striatius, C. sedentarius, and C. capistratus) were evaluated monthly during 2012. Counting was carried upon using visual censuses along 30 x 10 m transects. The highest lionfish abundances were found at Morro (2.60 ± 2.12) and the lowest at Remanso (0.58 ± 1.16), however, no differences were found between localities (p > 0.05). Highest abundance was recorded in June and July and the lowest in March, September, and October; no lionfish was observed in February. The highest abundances of both, commercially important and reef health important species were detected in the PNNT localities. The smallest lionfish and H. plumierii specimens were found at Morro, Remanso, and Isla Aguja. The size of the species was similar for all localities and lionfish sizes where larger. This may indicate that the studied species are not large enough to compete and thus they may be displaced, likewise their low abundances may be due to other stress factors. In any case, the invader has not reach yet the abundances shown in other Caribbean regions and then it may not be still causing a negative effect on the reef community.
KEY WORDS: Pterois volitans, Lionfish, Invasion, Colombian Caribbean.
INTRODUCCIÓN
Los arrecifes coralinos figuran entre las comunidades biológicas más productivas y taxonómicamente más diversas (Polovina, 1984). Esto resulta de la elevada heterogeneidad en cuanto a factores físicos, químicos y biológicos que presentan estos ecosistemas (Glynn, 1976). Sin embargo, su deterioro por diversos factores climáticos y antrópicos han hecho evidente la necesidad de generar información de las posibles causas y variables que afectan estos sistemas naturales (Mónaco et al., 2010).
La introducción de especies exóticas o no nativas, principalmente por comercio de especies ornamentales, ha sido considerada un problema para los arrecifes, tan importante como la eutrofización y la utilización de productos químicos tóxicos (Calado y Chapman, 2006). El pez león (Pterois volitans) es una especie originaria de los océanos Índico y Pacífico que, debido a su forma atractiva y su adaptación al cautiverio, ha sido ampliamente comercializada a nivel mundial para acuarios marinos (Morris et al., 2009). Su introducción al océano Atlántico en la década de los años ochenta del siglo pasado ocurrió en varios eventos por posibles liberaciones accidentales o intencionales al medio marino en la costa este de Estados Unidos (Whitfield et al., 2002). Su presencia en el Caribe ha sido exitosa ya que, además de colonizar rápidamente distintos hábitats, posee una dieta generalista, donde los peces han sido afectados principalmente por cambios en la abundancia, diversidad, riqueza y uniformidad en las poblaciones (Albins y Hixon, 2008, 2011; Brown et al., 2009; Darling et al., 2011; Albins, 2012).
El efecto negativo del pez león puede actuar conjuntamente con otras causas de deterioro como el calentamiento global, la contaminación y la pesca intensiva (Côté y Green, 2012; Morris, 2012). Esta última puede llevar al deterioro del hábitat por el uso de técnicas de pescas destructivas y la captura de peces no objetivo de gran importancia para la salud arrecifal, como los peces herbívoros que controlan la proliferación algal (Russ y Alcala, 1989). En Colombia los estudios realizados sobre el pez león hasta el momento se han dirigido a registrar su presencia y evaluar sus hábitos alimentarios (González et al., 2009; Martínez-Viloria et al., 2011; Muñoz-Escobar y Gil-Agudelo, 2012). La existencia de áreas de control, acceso y manejo por el sistema de Parques Nacionales Naturales permitió definir, de acuerdo con la presión pesquera, la premisa en la existencia de diferencias en talla y número de posibles controladores naturales del pez león, así como de aquellas indicadoras de salud arrecifal. Por lo cual, el propósito del estudio fue medir la abundancia y talla de ocho especies categorizadas de importancia comercial y ecológica como indicativo de si la presencia del pez león puede afectar su dinámica en la comunidad íctica arrecifal.
ÁREA DE ESTUDIO
Se muestrearon cinco arrecifes de coral de la región de Santa Marta, tres de los cuales son localidades cercanas a zonas urbanas, siendo más vulnerables a las escorrentías y alta presión de pesca: Morro de Santa Marta (11°14’57.94"N, 74°13’49.99"O), Remanso (11°16’26.00"N, 74°12’24.00"O) e Isla Aguja (11°19’8.29"N, 74°12’3.00"O). Los dos restantes, bahías de Chengue (11°19’’40.58"N, 74°8’7.92"O) y Cinto (11°20’19.68"N, 74°3’16.65"O), localidades pertenecientes al Parque Nacional Natural Tayrona (PNNT), están más alejados de las zonas urbanas, son de difícil acceso y en ellas se prohíbe la pesca y el turismo (Figura 1).
MATERIALES Y MÉTODOS
Se seleccionaron tres especies de importancia comercial (Haemulon plumierii, Lutjanus mahogoni y Cephalopholis cruentata) por ser aun relativamente abundantes y considerarlas como aquellas que pudieran ejercer algún control natural en zonas arrecifales y cinco especies de importancia para la salud arrecifal (Chaetodon ocellatus, C. sedentarius, C. striatus, C. capistratus y Abudefduf saxatilis), basados en su frecuencia, hábitos no crípticos y fácil identificación en campo. Se utilizó el método de censo visual por medio de un transecto fijo de banda descrito por Brock (1954), modificado a 30 m de largo y 10 m de ancho (300 m2). Se nadó en forma de zig-zag anotando el número de individuos de las especies seleccionadas y se determinó las tallas (cm LT) con la ayuda de una vara metreada. Para el registro del pez león se utilizó la misma metodología y se hizo necesaria la búsqueda de los individuos en grietas y oquedades donde pudiera refugiarse. Los muestreos se realizaron mensualmente entre diciembre 2011 y diciembre 2012.
La abundancia para cada una de las especies y las localidades de muestreos se analizó a través de análisis de varianza no paramétrica (Kruskal-Willis) y la abundancia de las especies entre localidades en los diferentes meses de muestreo se analizó a través de un Anova multivariado no paramétrico a una vía con permutaciones (NPMANOVA). Se realizó igualmente un análisis discriminante canónico que relaciona los sitios de muestreo y la abundancia de las especies en estudio a través del programa R versión 2.15.1 (R Development Core Team 2012 disponible en: www.r-project.org).
RESULTADOS
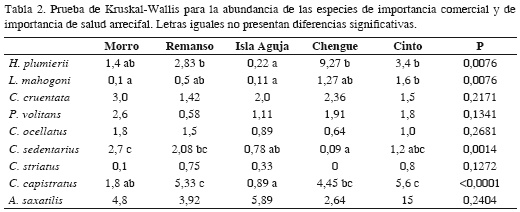
Para el pez león, en el año de muestreo se encontró la mayor abundancia en Morro de Santa Marta (2.60 ± 2.12 ind./300 m2), seguida de bahía de Chengue (1.91 ± 2.26) y bahía de Cinto (1.80 ± 2.04) y la menor en Remanso (0.58 ± 1.16). La prueba de Kruskal-Wallis no mostró diferencias significativas entre localidades (p > 0.05). La talla promedio más alta se encontró para Remanso (18 ± 2.8 cm) y la menor en el Morro de Santa Marta (16.8 ± 5.2 cm) y bahía de Cinto (15.9 ± 3.9). Sin embargo, estas dos últimas localidades presentaron una mayor variación de tamaño de los especímenes, con una talla máxima de 28 y 22 cm, respectivamente, y una mínima de 9 cm (Tabla 1).
En general, la especie de importancia comercial más abundante fue Haemulon plumierii (3.58 ± 7.2 ind/300 m2) con una talla promedio de 16.3 ± 3.3 cm; la bahía de Chengue fue la localidad donde se registró la mayor cantidad promedio de individuos (9.27 ± 12.62) e Isla Aguja la menor (0.22 ± 0.44). Cephalopholis cruentata tuvo una abundancia promedio total de 2.04 ± 1.6 ind/300 m2 con una talla de 15.2 ± 2.6 cm, encontrándose en Morro de Santa Marta como la especie más abundante (3.02 ± 1.76) y menor para Remanso (1.42 ± 1.24). Lutjanus mahogoni fue la especie menos abundante (0.73 ± 1.32), con una talla de 16.1 ± 1.7 cm (Tabla 1). El análisis de Kruskal-Wallis mostró diferencias significativas entre la abundacia de las especies (p < 0.05), excepto entre el pez león y H. plumierii.
Entre las especies de salud arrecifal, Abudefduf saxatilis fue en promedio la más abundante (6.29 ± 8.7 ind/300 m2) con una talla de 13.2 ± 1.7 cm; la bahía de Cinto fue la localidad con mayor abundancia (15.04 ± 12.45) y Chengue la menor (2.64 ± 2.11). Chaetodon capistratus siguió en abundancia (3.7 ± 3.2) con tallas de 11.3 ± 1.9 cm, e igualmente Cinto fue la localidad en donde esta especie fue más abundante (5.61 ± 2.67). Chaetodon sedentarius, C. ocellatus y C. striatus fueron las especies menos abundantes con menos de 2 ind./300m2 y solo en Morro de Santa Marta se encontró un promedio de 2.71 ± 1.70 ind./300 m2 (Tabla 1). El análisis de Kruskal-Wallis mostró diferencias significativas entre la abundancia de las especies (p < 0.05), solo C. ocellatus y C. sedentarius no presentaron diferencias significativas con la abundancia del pez león.
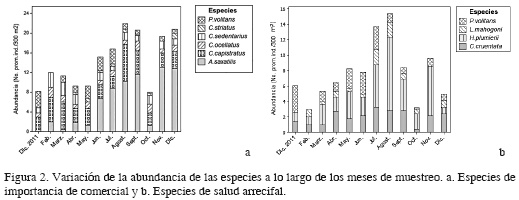
La variación temporal mostró que A. saxatilis fue la especie más abundante, observándose entre 8 y 13 individuos entre junio y diciembre a excepción de octubre donde solo se observó un promedio entre todas las localidades de 1.2 ind./300 m2. Chaetodon capistratus presentó una abundancia similar en todas las localidades durante el año de muestreo con valores promedio entre 2.5 y 7.6 ind./300 m2. La especie menos abundante fue C. striatus, siendo ausente en la mayoría de los meses y solo en diciembre de 2012 se observó 1.2 individuos en promedio para todas las localidades (Figura 2a). Con respecto a las especies de importancia comercial H. plumierii fue la más abundante en agosto (9.6 ind./300 m2) y noviembre (6.4 ind./300 m2). Cephalopholis cruentata presentó mayor abundancia en julio (3.25 ind./300 m2) mientras que L. mahogoni solo presentó 2 ind./300 m2 en julio y agosto (Figura 2b).
Lutjanus mahogoni y H. plumierii mostraron diferencias significativas entre localidades, siendo ambas más abundantes en las bahías de Chengue y Cinto. En cuanto a C. cruentata y el pez león, las mayores abundancias estuvieron dadas en Morro de Santa Marta; sin embargo, la prueba de Kruskal-Wallis entre localidades no mostró diferencias significativas para estas dos especies (Tabla 2). A pesar que el pez león presentó una mayor abundancia que C. striatus y C. ocellatus entre localidades, la prueba de Kruskal-Wallis no mostró diferencias significativas en la abundancia de estas especies (p > 0.05).
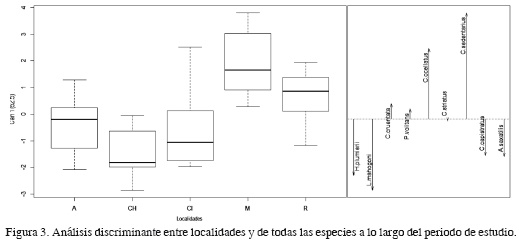
El análisis multivariado a lo largo del período de estudio y entre localidades, teniendo en cuenta tanto las especies de importancia comercial como de salud arrecifal, mostró diferencias significativas (F = 2.07, gl = 9, p = 0.00099). En adición, el análisis discriminante mostró que el primer eje canónico explica 62.5% de la variación, en donde las localidades de Isla Aguja, bahía de Chengue y bahía de Cinto se diferenciaron de Morro de Santa Marta y Remanso en cuanto a la abundancia de los peces; esta diferencia está dada principalmente por las especies H. plumierii, L. mahogoni, C. capistratus y A. saxatilis (Figura 3).
DISCUSIÓN
El bajo número de individuos del pez león en Remanso e Isla Aguja y los altos valores encontrados en Morro de Santa Marta coinciden con lo registrado por Muñoz-Escobar y Gil-Agudelo (2012), quienes encontraron 41 ejemplares de esta especie en la bahía de Gaira y Morro de Santa Marta. González-C et al. (2011) también registraron una mayor captura de pez león en Santa Marta que en el PNNT, sin embargo, ese estudio presentó un mayor esfuerzo de muestreo en Santa Marta entre 2009 y 2010. La baja abundancia en Remanso e Isla Aguja se atribuye a que son sitios de buceo y pesca donde se ejerce una alta presión de control del pez león.
En febrero, cuando la temperatura superficial del agua registra un mínimo de 21.8°C (CIOH, 2012), y en octubre, cuando se presenta una alta turbiedad del agua, se observó el menor número de individuos para todas las especies. Para el caso de P. volitans, éste se hace más letárgico e inactivo con las bajas temperaturas, llevándolo a permanecer mayor tiempo refugiado en grietas (Kimball et al., 2004); sin embargo, para el resto de las especies la baja abundancia registrada puede obedecer a limitaciones de los observadores debido a la temperatura y turbiedad. Con el aumento de la temperatura superficial del agua (Bernal et al., 2006) se registraron en todas las localidades mayores abundancias de todas las especies, a excepción de Remanso, cuya formación arrecifal presenta una baja cobertura coralina (10.4%) (Hernández-Abello, 2013). Según Biggs y Olden (2011), arrecifes con alta cobertura coralina favorecen al pez león ya que la estructura arrecifal ofrece mayores oportunidades de camuflaje y protección que otro tipo de sustratos.
En lo que respecta a las especies de importancia comercial, la abundancia del pez león fue más alta que la de Lutjanus mahogoni, pero significativamente menor que Haemulon plumierii y no presentó diferencias significativas con Cephalopholis cruentata. Lo anterior indica que el pez león ha ocupado distintos hábitats al igual que las especies de importancia comercial, tales como los meros y pargos. Biggs y Olden (2011), al describir las preferencias que el pez león y las especies de meros (Mycteroperca venenosa y Epinephelus striatus) y pargos (Lutjanus jocu y L. analis) presentan con el sustrato, encontraron patrones similares de ocupación en microhábitats como corales duros y arenas, lo cual, según Morris (2012), ocurre porque el pez león compite por recursos y espacios con dichas especies.
En sitios protegidos por el sistema de Parques Nacionales se registró mayor abundancia de H. plumierii y L. mahogoni. Olaya-Restrepo et al. (2008) para estas especies encontraron abundancias relativas similares en zonas del PNNT con una menor abundancia en localidades fuera del PNNT. Cuadrado (2002), alrededor de Isla Aguja, al cuantificar la comunidad de peces asociados encontró bajas abundancias para especies de importancia comercial y Grijalba-Bendeck et al. (2004) para L. mahogoni encontraron en Morro de Gaira una abundancia relativamente alta de 0.47 individuos bajo el método de cilindro imaginario de 7.5 m de radio, siendo superior que H. plumierii (0.02). Lo anterior indica como las especies de importancia comercial tienen una mayor presión de pesca en zonas donde no hay ninguna protección. Por ejemplo, Whitfield et al. (2007), compararon especies como meros entre áreas marinas protegidas desde el norte de la Florida a Carolina del Norte y sitios expuestos a la presión de pesca en Florida, Georgia y Carolina del Sur con la abundancia del pez león y, aunque no mostraron diferencias significativas en la abundancia de Cephalopholis cruentata, Mycteroperca microlepis y Epinephelus adscensionis, si encontraron una abundancia superior en áreas marinas protegidas de las especies M. interstitialis y E. guttatus. Por su parte, el pez león no presentó diferencias entre las áreas protegidas y expuestas, lo cual concuerda con lo observado para este estudio.
Al analizar las tallas de capturas para el Caribe por Duarte et al. (1999) y MADR y CCI (2006) para L. mahogoni y H. plumierii, éstas muestran un promedio de 22.5 y 22 cm LT, respectivamente, tallas superiores al promedio encontrado en este estudio para todas las localidades. Para el pez león, Cárdenas (2011) registra una talla máxima de 32 cm para Santa Marta y Morris y Akin (2009) indican que un individuo del tamaño como el registrado por Cárdenas (2011) puede consumir presas de 14 a 16 cm, lo cual es el ámbito de talla promedio para la mayoría de las especies estudiadas. Por lo cual, la sobreexplotación de peces grandes y posibles competidores del pez león, puede significar una resistencia natural reducida, desplazamiento o incluso depredación por el invasor.
Sobre las especies catalogadas como de importancia de salud arrecifal, estudios realizados para el Caribe prueban el efecto negativo que tiene el pez león, ya sea por depredación o desplazamiento por competencia. Chaetodon striatus presentó en el estudio una abundancia menor que el pez león; sin embargo, no se puede asegurar que su baja abundancia esté dada como respuesta a la invasión, ya que Acero y Rivera (1992) ya registraban densidades bajas para el PNNT y localidades aledañas. Para peces como C. striatus, C. sedentarius, C. ocellatus y C. capistratus, Albins y Hixon (2008) realizaron en las Bahamas un experimento para determinar el efecto de la presencia del pez león respecto al desplazamiento de las especies; los resultados mostraron una reducción significativa (hasta un 80%) en el reclutamiento de las especies de peces en cinco semanas.
Albins (2012) menciona que los efectos negativos de la invasión conllevan cambios en la diversidad, riqueza y uniformidad de las comunidades de peces nativos. Abudefduf saxatilis, que fue la especie más abundante, ya se ha encontrado por MuñozEscobar y Gil-Agudelo (2012) en el contenido estomacal de un pez león de 18.2 cm en el PNNT al igual que por Valdez-Moreno et al. (2012), quienes a través de técnicas moleculares identificaron a A. saxatilis entre las presas consumidas por el pez león. Esta especie, que según Carter (2002) es una de las más frecuentes en los arrecifes del Caribe, podría ver reducida su abundancia por efecto de la depredación del invasor.
La densidad del pez león puede aumentar significativamente en bajos periodos de tiempo; por ejemplo, Green y Côté (2009) registraron en las Bahamas una abundancia promedio de 393 individuos por hectárea de mayo a julio y Ruttenberg et al. (2012) registraron un incremento de las densidades y la biomasa del pez león seis veces mayor desde 2010 a 2011 en los cayos de la Florida. Por lo tanto, el registro de sus abundancias y el seguimiento y monitoreo ambiental para documentar cambios en las poblaciones de peces arrecifales luego de la invasión son necesarios con el fin de establecer planes de acción para mitigar los posibles impactos en el Caribe colombiano.
AGRADECIMIENTOS
Esta investigación fue financiada por Colciencias (código: 1361-52128271), ejecutada por la Universidad Nacional de Colombia, y por Fonciencias por medio de la Universidad del Magdalena.
BIBLIOGRAFÍA
Acero P., A. y M. Rivera. 1992. Peces de las familias Chaetodontidae y Pomacanthidae en la región de Santa Marta (Colombia): densidad y relación con la calidad del arrecife. Carib. J. Sci., 28(3): 184 190. [ Links ]
Albins, M. 2012. Effects of invasive Pacific red lionfish Pterois volitans versus a native predator on Bahamian coral-reef fish communities. Biol. Inv., 15: 29-43. [ Links ]
Albins, M. y M. Hixon. 2008. Invasive Indo-Pacific lionfish Pterois volitans reduce recruitment of Atlantic coral-reef fishes. Mar. Ecol. Prog. Ser., 367: 233-238. [ Links ]
Albins, M. y M. Hixon. 2011. Worst case scenario: potential long-term effects of invasive predatory lionfish (Pterois volitans) on Atlantic and Caribbean coral-reef communities. Environ. Biol. Fish., DOI 10.1007/s10641-011-9795-1. [ Links ]
Bernal, G., G. Poveda, P. Roldán y C. Andrade. 2006. Patrones de variabilidad de las temperaturas superficiales del mar en la costa Caribe colombiana. Rev. Acad. Colomb. Cienc., 30(115): 195-208. [ Links ]
Biggs, C. y J. Olden. 2011. Multi-scale habitat occupancy of invasive lionfish (Pterois volitans) in coral reef environments of Roatán, Honduras. Aq. Inv., 6: 347-353. [ Links ]
Brock, V. 1954. A preliminary report on a method of estimating reef fishes. J. Wildl. Manag., 18(3): 297-308. [ Links ]
Brown, M., J. Cowhey, S. Dawson, M. Farrington, M. Nash, J. Shi y S. Young. 2009. Lionfish (Pterois volitans) affecting biodiversity of patch reef species. Marine Mar. Ecol. Prog. Ser., 367: 233-238. [ Links ]
Calado, R. y P. Chapman. 2006. Aquarium species: Deadly invaders. Mar. Pollut. Bull., 52: 599-601. [ Links ]
Cárdenas, E. 2011. Aspectos tróficos y reproductivos del pez león Pterois volitans (Linnaeus, 1758) en el área de Santa Marta, Caribe colombiano. Trabajo grado, Univ. Bogotá Jorge Tadeo Lozano, Santa Marta. 69 p. [ Links ]
Carter, J. 2002. Pomacentridae: Damselfishes. The living marine resources of the Western Central Atlantic. FAO Species Identification Guide for Fishery Purposes and American Society of Ichthyologists and Herpetolologists.Vol.2, Roma. 1550 p. [ Links ]
CIOH. 2012. Pronóstico climático del Caribe colombiano de octubre a noviembre http://www.cioh.org.co/meteorologia/PronosticoClimatologico.php.11/03/2013. [ Links ]
Côté, I. y S. Green. 2012. Potential effects of climate change on a marine invasion: The importance of current context. Curr. Zool., 58: 1-8. [ Links ]
Cuadrado, J. 2002. Caracterización de la estructura de las comunidades ícticas en las zonas arrecifales de Isla Aguja, Parque Nacional Natural Tayrona, Caribe colombiano. Trabajo de grado, Pontificia Univ. Javeriana, Bogotá. 125 p. [ Links ]
Darling, E., S. Green, J. Leary y I. Côté. 2011. Indo-Pacific lionfish are larger and more abundant on invaded reefs: a comparison of Kenyan and Bahamian lionfish populations. Biol. Invasions, 13: 2045-2051. [ Links ]
Duarte, L., C. García, N. Sandoval, D. von Schiller, G. Melo y P. Navajas. 1999. Length-weight relationships of demersal fishes from the Gulf of Salamanca, Colombia. Naga, 22(1): 34-36. [ Links ]
Glynn, P. 1976. Some physical and biological determinants of coral community structure in the eastern Pacific. Ecol. Monogr., 46: 431-456. [ Links ]
González, J., M. Grijalba-Bendeck, A. Acero P. y R. Betancur-R. 2009. The invasive red lionfish, Pterois volitans (Linnaeus 1758), in the southwestern Caribbean Sea. Aq. Inv., 4(3): 507-510. [ Links ]
González-C, J.D., A. Acero P., A. Serrat-LL. y R. Betancur-R. 2011. Caracterización taxonómica de la población del pez león Pterois volitans (Linnaeus 1758) (Scorpaenidae) residente en el Caribe colombiano: merística y morfometría. Biota Colomb., 12(2): 15-22. [ Links ]
Green, S. y I. Côté. 2009. Record densities of Indo-Pacific lionfish on Bahamian coral reefs. Coral Reefs, 28: 107. [ Links ]
Grijalba-Bendeck, M., E. Castañeda-Moya y A. Acero P. 2004. Estructura de un ensamblaje íctico asociados a fondos duros en el Caribe colombiano empleando la técnica del censo visual estacionario (CVE). Actual. Biol., 26(81): 197-211. [ Links ]
Hernández-Abello, J. 2013. Estructura de talla y selectividad al sustrato del pez león (Pterois volitans) en Santa Marta, Caribe colombiano. Trabajo de grado, Univ. Magdalena, Santa Marta. 57 p. [ Links ]
Kimball, M., J. Miller, P. Whitfield y J. Hare. 2004. Thermal tolerance and potential distribution of invasive lionfish (Pterois volitans/miles complex) on the east coast of the United States. Mar. Ecol. Progr. Ser., 283: 269-278. [ Links ]
MADR y CCI. 2006. Pesca y acuicultura Colombia 2006. Informe técnico regional litoral Caribe y Pacífico, Minist. Agric. Des. Rur. Inst. Colomb. Des. Rur., Bogotá. 138 p. [ Links ]
Martínez-Viloria, H., A. Rosado y A. Acero P. 2011. Presencia del pez león, Pterois volitans (Actinopterygii: Scorpaenidae), en el departamento de La Guajira, mar Caribe de Colombia. Bol. Invest. Mar. Cost., 40(2): 445-447. [ Links ]
Mónaco, C., S. Narciso, F. Alfonso, E. Jiménez y F. Bustillo. 2010. Evaluación de las comunidades de corales y peces de algunos arrecifes de la Isla la Tortuga y cayos adyacentes, Venezuela. Bol. Centro Invest. Biol., 44(3): 355-378. [ Links ]
Morris, J. 2012. The lionfish invasion: Past, present and future. 1-2. En: Morris, J.A. (Ed.). Invasive lionfish: A guide to control and management. Gulf and Caribbean Fisheries Institute Special Publication Series Number 1, Marathon, EE.UU. 113 p. [ Links ]
Morris, J. y J. Akins. 2009. Feeding ecology of the invasive lionfish (Pterois volitans) in the Bahamas archipelago. Environ. Biol. Fish., 86: 389-398. [ Links ]
Morris, J., J. Akins, A. Barses, D. Cerino, D. Freshwater, S. Green, R. Muñoz, C. Paris y P. Whitfield. 2009. Biology and ecology of the invasive lionfishes, Pterois miles and Pterois volitans. Proc. Gulf Carib. Fish. Inst., 29: 409-414. [ Links ]
Muñoz-Escobar, L. y D. Gil-Agudelo. 2012. Composición dietaría del pez león, Pterois volitans (Pisces: Scorpaenidae) en Santa Marta y el Parque Nacional Natural Tayrona. Bol. Invest. Mar. Cost., 41(2): 471-477. [ Links ]
Olaya-Restrepo, J., C. Reyes-Nivia y A. Rodríguez-Ramírez. 2008. Ensamblaje de peces arrecifales en área de Santa Marta y el Parque Nacional Natural Tayrona. Bol. Invest. Mar. Cost., 37(1): 111-127. [ Links ]
Polovina, J. 1984. Model of a coral reef ecosystem. Coral Reefs, 3(1): 1-11. [ Links ]
R Development Core Team. 2012. R: A language and environment for statistical computing. R. foundation for statistical computing, Vienna. 1706 p. ISBN 3-900051-07-0, URL: http://www.R-project.org. [ Links ]
Russ, G. y A. Alcala. 1989. Effects of intense fishing pressure on an assemblage of coral reef fishes. Mar. Ecol. Progr. Ser., 56: 13-27. [ Links ]
Ruttenberg, B., P. Schofield, J. Akin, A. Acosta, M. Feeley, J. Blondeau, S. Smith y J. Ault. 2012. Rapid invasion of Indo-Pacific lionfishes (Pterois volitans and Pterois miles) in the Florida keys, USA: evidence from multiple pre- and post invasion data sets. Bull. Mar. Sci., 88(4): 1051-1059. [ Links ]
Valdez-Moreno, M., C. Quintal-Lizama, R. Gómez-Lozano y M. García-Rivas. 2012. Monitoring an alien invasion: DNA barcoding and the identification of lionfish and their prey on coral reefs of the Mexican Caribbean. Plos One, 7(6): e36636. [ Links ]
Whitfield, P., T. Gardner, S. Vives, M. Gilligan, W. Courtenay, G. Ray y J. Hare. 2002. Biological invasion of the Indo-Pacific lionfish Pterois volitans along the Atlantic coast of North America. Mar. Ecol. Prog. Ser. 235: 289-297. [ Links ]
Whitfield, P., J. Hare, A. David, S. Harter, R. Muñoz y C. Addison. 2007. Abundance estimates of the Indo-Pacific lionfish Pterois volitans/miles complex in the Western North Atlantic. Biol. Inv., 9: 53-64. [ Links ]
RECIBIDO: 22/05/2014 ACEPTADO: 27/01/2015
