










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.45 no.1 Santa Marta Jan./June 2016
INFLUENCIA DEL RÍO ATRATO EN EL GOLFO DE URABÁ DURANTE EL HOLOCENO TARDÍO, MAR CARIBE COLOMBIANO
INFLUENCE OF THE ATRATO RIVER IN THE GULF OF URABÁ DURING THE LATE HOLOCENE, COLOMBIAN CARIBBEAN SEA
Camilo Vélez-Agudelo1 y Néstor Aguirre-Ramírez2
1 Universidad de Antioquia, Grupo de Investigación en Gestión y Modelación Ambiental (GAIA), Calle 62 No 52-59, Sede de Investigaciones Universitarias (SIU), Medellín, Colombia. kmilovelez82@gmail.com
2 Universidad de Antioquia, Grupo Geolimna, Calle 67 No 53-108, Medellín, Colombia. naguirre@udea.edu.co
RESUMEN
A partir del análisis sedimentológico, geoquímico (carbono orgánico total COT, nitrógeno total NT, fósforo total PT, sílice biogénico SiB y carbonato de calcio CaCO3) y de diatomeas en dos núcleos de sedimento se infirieron cambios generales en las condiciones ambientales que dominaron el norte del golfo de Urabá durante el Holoceno tardío. Los modelos de edad (14C AMS) sugieren que el núcleo de la margen noreste (SP 9) corresponde a ∼960 años AP, en tanto que en el núcleo de la margen noroeste (SP 41) el modelo se ajusta a los últimos ∼2845 años AP. Los resultados indican que los sitios de perforación están afectados por procesos hidrodinámicos distintos que, en general, se han mantenido durante el Holoceno tardío. Mientras que en la margen noroeste prevalecen procesos de naturaleza oceánica con aportes menores de sedimentos fluviales, en la margen noreste del golfo existen procesos que favorecen el transporte y la acumulación de sedimentos terrígenos, los cuales limitan el crecimiento de las diatomeas. Entre 2845 y 2489 años AP se asume un mayor nivel del mar y condiciones húmedas en la margen noroeste del golfo. El descenso gradual del nivel del mar desde ∼2445 años AP, favoreció la progradación del delta hacia la margen oriental del golfo, proceso que se habría intensificado en los últimos ∼200 años AP debido, entre otros factores, a una combinación de factores naturales y antrópicos. Aunque las descargas fluviales son canalizadas predominantemente hacia la margen oriental del golfo, la tendencia de aumento del COT y el registro de diatomeas dan cuenta de aportes fluviales en la margen noroeste del golfo desde hace ∼800 años AP, los cuales podrían estar asociados a una mayor actividad de boca Tarenas, el brazo más al norte del río Atrato.
PALABRAS CLAVES: Diatomeas, Carbono orgánico, Nitrógeno total, Sílice biogénico, Holoceno tardío.
ABSTRACT
Sedimentary, geochemical (total organic carbon TOC, total nitrogen TN, total phosphorus TP, biogenic silica BSi and calcium carbonate (CaCO3) and diatoms data of two sediment cores were used to infer general environmental changes in the northern Gulf of Urabá during the late Holocene. The age models(AMS 14C ) suggested that core of the northeastern margin (SP 9) belong to ∼960 years BP, while in the core of the northwestern margin (SP 41) the model was adjusted for the last ∼2845 years BP. The results indicated that drilling sites are affected by different hydrodynamic processes that, in general, have been maintained during the late Holocene. While in the northwestern margin oceanic processes dominate and there is low input of fluvial sediments to this site, in the northeastern margin there are processes that favor the transport and accumulation of terrigenous sediments, which limit the growth of diatoms. Between 2845 and 2489 year BP, a higher sealevel and wet conditions were recorded in the gulf. Since ∼2445 years BP a gradual decrease in sea level allowed delta progradation towards the eastern side of the gulf. Apparently, this process would have intensified its presence in the last ∼200 year BP, due to natural and anthropogenic factors. Even though the river discharges are channeled predominantly towards the east side of the Gulf, the trend of increasing TOC and diatoms data suggested fluvial input to the northwestern margin from ∼800 years BP, which may be associated with a higher activity of Tarenas mouth, the northern branch of the Atrato River.
KEY WORDS: Diatoms, Organic carbon, Total nitrogen, Biogenic silica, Late Holocene.
INTRODUCCIÓN
La Unidad Ambiental Costera del Darién (UAC-Darién) es una región extensa de territorio marino y costero ubicada en el extremo suroeste de la costa Caribe colombiana. En esta unidad ambiental se incluye al golfo de Urabá, el accidente costero (golfo) más grande de Colombia sobre el mar Caribe, una ecorregión que por su ubicación geográfica, características hidroclimatológicas e hidrobiológicas lo posicionan en la agenda estratégica de desarrollo regional y nacional (Invemar, 2005; Invemar et al., 2008).
Ecosistemas frágiles altamente variables y diversos, dinámicas oceanográficas intensas, contextos geológicos activos (diapirismo y volcanes de lodo) y rápido deterioro ambiental de sus aguas y playas son, en buena medida, características distintivas del golfo. Como receptor de aguas y sedimentos de una extensa cuenca, el golfo de Urabá está expuesto continuamente a un progresivo deterioro de la calidad de sus aguas debido, entre otros factores, a actividades como el saneamiento de buques, la deforestación intensiva, el uso de agroquímicos y fertilizantes y al vertimiento de aguas residuales domiciliares e industriales (Chevillot et al., 1993; García-Valencia, 2007; Álvarez-Láinez, 2008; Montoya, 2010). La erosión generalizada de sus costas y de la cuenca de sus tributarios, es otro de los problemas que en la actualidad afecta severamente el golfo (Thomas et al., 2007). Estudios realizados entre Arboletes y Turbo indican que el retroceso de la línea de sus costas es resultado del efecto combinado de factores naturales (ascenso relativo del nivel del mar, fenómenos tectónicos, hidroisostacia) y antrópicos (desviación de cauces fluviales, extracción intensiva de materiales de playa, construcción no planificada de estructuras rígidas) que actúan a distintas escalas de tiempo (Correa y Vernette, 2004).
Aunque en los últimos años se ha avanzado, el conocimiento de estas características en el golfo es incipiente y fragmentario, por tanto, la síntesis e integración de la información sobre sus aspectos oceanográficos, geológicos, geomorfológicos y biológicos son necesarios en la formulación de programas que fortalezcan el manejo integrado de zonas costeras, especialmente en escenarios como los planteados en el cambio climático global y el subsecuente aumento en el nivel del mar. Ante la ausencia de esta línea base en el golfo, los registros paleoambientales generados a partir del estudio de indicadores físicos, geoquímicos y biológicos (diatomeas, foraminíferos, ostrácodos, entre otros) conservados en sedimentos lacustres o marinos, permiten obtener información ambiental de periodos anteriores a la implementación del monitoreo instrumental moderno (Willis y Birks, 2006; Cremer et al., 2007; Cooper, 2010).
En ambientes estuarinos como el golfo de Urabá, elementos biogénicos de origen autóctono o alóctono, experimentan una intensa degradación biológica a medida que decantan en la columna de agua (Seiter et al., 2004; Bianchi y Bauer, 2011). Una pequeña fracción alcanza la fase sedimentaria y en ella son expuestos nuevamente a reacciones diagenéticas que favorecen la acumulación o el intercambio de nutrientes entre los sedimentos y el agua suprayacente. Factores como la productividad primaria, las fuentes del material orgánico, la interacción con partículas minerales, la profundidad de la columna de agua, las tasas de sedimentación, la bioturbación y la concentración de oxígeno disuelto determinan la cantidad, la distribución vertical, y la composición química del material orgánico que se acumula en los sedimentos ( Slomp, 2011; Statham, 2012; Arndt et al., 2013).
Las diatomeas son consideradas excelentes indicadores biológicos que ofrecen información confiable sobre cambios ambientales recientes y holocénicos, en parte por su alta diversidad, amplia distribución en ecosistemas acuáticos, cortos tiempos de generación, sensibilidad diferencial a cambios ambientales específicos y a la preservación de sus frústulos silíceos (Battarbee et al., 2001). A partir de las características autoecológicas de este grupo de microalgas es posible reconstruir fluctuaciones de variables ambientales como temperatura del agua superficial, pH, salinidad, entrada de agua dulce en ambientes estuarinos, variación del nivel del agua, eutrofización y productividad de ecosistemas acuáticos (Mackay et al., 2005; Fritz, 2007; Korhola, 2007).
Aunque son pocas las investigaciones dirigidas a estudiar cambios holocénicos en el golfo de Urabá, algunos trabajos sedimentológicos y de cambios en la malacofauna dan cuenta de variaciones naturales y del efecto de algunas actividades antrópicas en la región (Ospina-Hoyos et al., 2014; Rúa et al., 2014, 2015). El objetivo de este estudio es ampliar el conocimiento de las condiciones ambientales predominantes en el golfo de Urabá durante el Holoceno tardío, a partir del análisis de carbono orgánico total (COT), nitrógeno total (NT), fósforo total (PT), sílice biogénico (SiB), carbonato de calcio (CaCO3) y de la determinación de diatomeas presentes en dos núcleos de sedimento.
ÁREA DE ESTUDIO
El golfo de Urabá es un sistema semicerrado y alargado en dirección norte-sur, localizado en el extremo occidental de la costa Caribe colombiana entre los departamentos de Antioquia y Chocó (García-Valencia, 2007; Invemar et al., 2008) (Figura 1). La precipitación media anual se estima en 2500 mm, con períodos de lluvias entre mayo y noviembre y períodos secos que van desde diciembre hasta mediados de abril; variaciones que obedecen a la oscilación anual de la zona de convergencia intertropical (ZCIT) y a los vientos alisios del noreste (Chevillot et al., 1993).
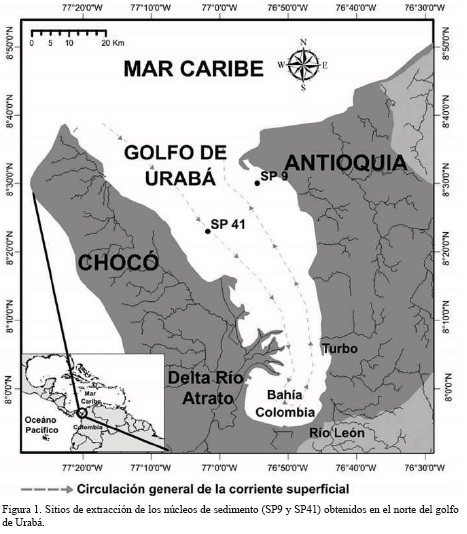
Con apoyo de imágenes satelitales y del conocimiento de los patrones de circulación de la pluma del río Atrato, Molina et al. (1992) y Chevillot et al. (1993) indicaron un fuerte acople del patrón de vientos con la circulación superficial del golfo de Urabá. Ellos identificaron no solo una corriente de entrada por la margen occidental y una corriente de salida por el costado oriental, sino también cambios estacionales en la salinidad superficial del golfo.
En el extremo suroccidental del golfo se localiza el delta del río Atrato, la principal fuente de descarga de aguas y sedimentos hacia el golfo. El caudal medio del río se estima en 2740 m3s-1, en tanto que la descarga de sedimentos es de 11.3 x 106 ton año-1, la segunda en aportes de sedimentos al mar Caribe después del río Magdalena (Restrepo y Kjerfve, 2004). Los ríos León, Caimán Viejo, Caimán Nuevo y Turbo, también desembocan en el golfo y aunque sus descargas y caudales son inferiores a las registradas en el Atrato, localmente pueden afectar la geomorfología e hidrodinámica costera cercana (Bernal et al.,2005; Álvarez-Láinez, 2008).
MATERIALES Y MÉTODOS
En la campaña oceanográfica de diciembre de 2009 a bordo del buque oceanográfico ARC Quindío se extrajeron 16 núcleos de sedimento, de los cuales se seleccionaron uno de la margen noreste (SP 9) (8°30'2,22''N; 76°54'28,14''O) y otro de la margen noroeste (SP 41) (8°22'58,2''N; 77°01'43,14''O) del golfo.
Para la extracción de las secuencias sedimentarias se empleó un nucleador de gravedad con un diámetro de 6.35 cm y capacidad de recuperación máxima de 5 m. En el núcleo SP 9 se obtuvieron 2.05 m de sedimentos y en el SP 41 2.25 m. Ecogramas del fondo y subfondo marino realizadas con una ecosonda Knudsen acoplada a un transductor de 200-12kHz, indican que en ambos sitios de perforación los sedimentos acumulados tienen un espesor mayor a la longitud de los núcleos extraídos. La profundidad de la columna de agua en los sitios de perforación fue de 34 y 46.2 m, respectivamente.
Para evaluar el efecto del río Atrato en los dos sitios de perforación, se tomaron muestras de agua con una botella Schindler cada metro desde la superficie hasta 10 m de profundidad y, posteriormente, cada cinco metros hasta el fondo. Con una sonda multiparamétrica IQ 100 se midieron, en cada profundidad, la temperatura del agua, la salinidad, el pH, la conductividad eléctrica y el oxígeno disuelto.
Los núcleos se almacenaron a una temperatura de 4°C en el laboratorio del grupo GAIA de la Universidad de Antioquia, donde se abrieron longitudinalmente y se hicieron las descripciones estratigráficas correspondientes (color, textura, estructuras sedimentarias, granulometría, contenido de bioclastos y detritos). Para obtener la edad de la secuencia sedimentaria, se tomaron restos de materia orgánica en el tope, centro y base de cada núcleo. Las muestras se enviaron al Centro de Investigaciones Ambientales de la Universidad de Glasgow (Reino Unido) para las dataciones radiocarbono (14C-AMS). Con el programa Oxcal 4.0 se calcularon los modelos de edad y se transformaron las fechas en edades calendarios (AP antes del presente).
Para los análisis geoquímicos y de diatomeas se tomaron submuestras de sedimento a intervalos de 5 cm con un espesor de 2 cm. En un contador Coulter LS130 se determinaron el tamaño y porcentaje de partículas sedimentarias de nueve submuestras. Antes de los análisis químicos, las muestras se secaron durante 24 horas en un liofilizador Labconco-Freezone 6 Plus. En la cuantificación del contenido de SiB, CaCO3 biogénico, PT, COT y NT, se emplearon los métodos sugeridos por Mortlock y Froelich (1989), Quiceno (2008), Rosales (1979), Strickland y Parsons (1972) y Walkley y Black (1934), respectivamente. Los resultados de cada indicador se expresaron en porcentajes (g/100g).
Para la determinación de las diatomeas se tomaron 0.5 g de sedimento húmedo y se trataron con HCl 10%, ácido nítrico 65%, ácido sulfúrico 95% y permanganato de potasio saturado con el fin de oxidar la materia orgánica y los carbonatos (Hasle et al., 1996; Battarbee et al., 2001). Placas permanentes con Zrax se utilizaron para facilitar la identificación y conteo microscópico. Se contaron y determinaron 400 valvas por preparado con un aumento de 1000X, mientras que observaciones en microscopio electrónico de barrido (MEB) apoyaron el análisis taxonómico de los taxones que requerían un mayor nivel de detalle.
Para la determinación taxonómica y la información ecológica de las diatomeas se emplearon los trabajos de Abrantes (1988-1991); Round et al. (1990), Cooper (1995), Hasle et al. (1996), Zong (1997), Witkowski et al. (2000), Sterrenburg (2001), Shiono y Koizumi (2002), McQuoid y Nordberg (2003), Affan et al. (2006), Witon y Witkowski (2006), Cremer et al. (2007) y Gebühr (2011). En la revisión de la nomenclatura y el estado de validez de los taxa se consultaron las bases de datos http://www.marinespecies.org y http://www.algabase.org.
En el análisis descriptivo básico de los datos se utilizó la media como medida de tendencia central, y la desviación estándar y el coeficiente de variación como medidas de dispersión absoluta y relativa, respectivamente. A partir de los indicadores geoquímicos evaluados y de las especies de diatomeas con abundancias relativas superiores a 3%, se realizó un análisis jerárquico de agrupamiento (clúster) basado en el método de vinculación de Ward. Para estos análisis se emplearon los programas Statgraphics Centurion, versión XVI y C2.
RESULTADOS
En general, el oxígeno disuelto, el pH y la temperatura del agua en la estación SP 41 permanecieron constantes en la columna de agua. Se observó un aumento gradual de la salinidad con la profundidad, con un valor promedio de 28.18 (CV = 14.934%) y valores de 22.5 en la superficie y 28.9 en el fondo. En la estación SP 9, los valores de temperatura y pH se mantuvieron constantes. En este punto la salinidad con un valor promedio de 29.0 (CV = 17.716%) aumentó gradualmente desde la superficie (21.4) hasta el fondo (33.5) de la columna de agua.
Cronología y litoestratigrafía
Las dataciones radiocarbono (AMS 14C) sugieren que los 2.25 m de sedimento en el núcleo SP 41 comprenden los últimos ∼2800 años AP, mientras que los 2.05 m del núcleo SP 9 corresponden a los últimos ∼960 años AP. Las secuencias sedimentarias presentaron características sedimentológicas similares: litológicamente los dos núcleos están constituidos por limos y limos arenosos de origen mixto (terrígeno y calcáreo), con fragmentos de conchas calcáreas (bivalvos y gasterópodos) y restos de madera (Figura 2).
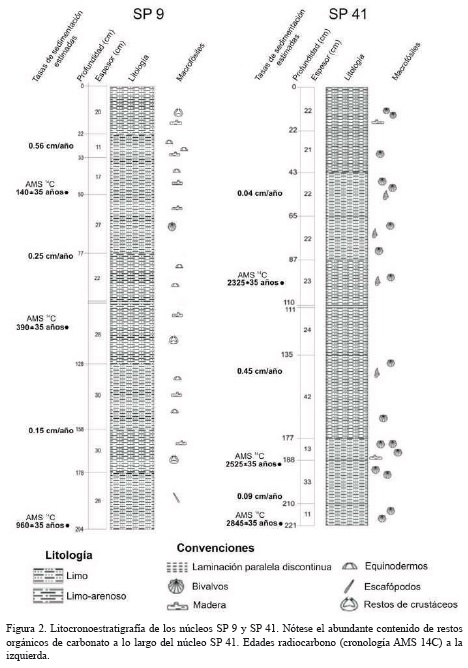
Las tasas de sedimentación estimadas entre la margen noroeste y noreste presentaron tendencias opuestas. Entre los 960 años y los 403 años AP la tasa de sedimentación estimada en el sitio SP 9 fue de 0.15 cm/año, entre los 403 años y los 144 años AP aumentó a 0.25 cm/año, y en los últimos 144 años AP alcanzó un máximo de 0.56 cm/año. Por su parte, los resultados del núcleo SP 41 indican que de 2845 a 2525 años AP la tasa de sedimentación fue de 0.09 cm/año, entre 2525 y 2325 años AP aumentó a 0.45 cm/año y entre 2325-62 años AP disminuyó a una tasa de 0.04 cm/año.
Registro geoquímico
En las figuras 3 y 4 se presentan los perfiles de COT, NT, PT, SiB y CaCO3, así como las relaciones elementales de C:N, N:P y C:P. En el núcleo SP 41 los valores del COT fluctuaron entre 0.97 y 2.19 %, con una ligera disminución desde la superficie del núcleo hasta los 2423 años AP, seguido por un aumento gradual entre 2434-2467 años AP. En el núcleo SP 9 las concentraciones de COT oscilaron entre 1.18 y 2.17 %, con valores mayores en las zonas media e inferior del núcleo. En general, se observó una disminución gradual del NT hacia la base de las secuencias, con una tendencia más marcada en el núcleo SP 41 donde los valores oscilaron entre 0.08 y 0.17 %. En el núcleo SP 9 el porcentaje de NT varió entre 0.11 y 0.16 %.
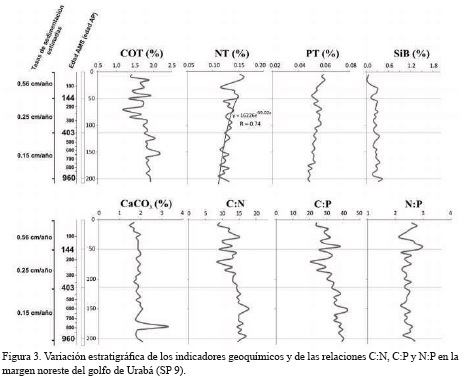
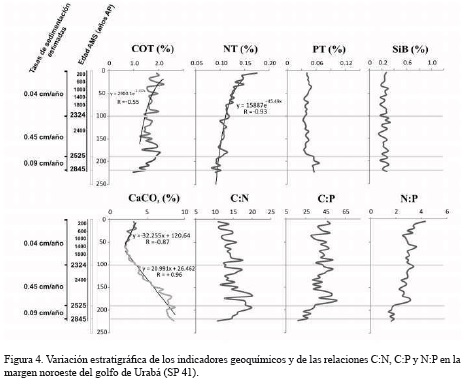
Aunque la concentración media de PT en las dos columnas de sedimento es similar (SP 9 = 0.053±0.003%, SP41 = 0.043±0.007%), se observaron tendencias distintas en ambas secuencias. Mientras en el núcleo SP 9 no se observaron variaciones significativas con la profundidad, en la base del núcleo SP 41, especialmente entre los 2522-2845 años AP, se registraron los mayores niveles. La correlación inversa estadísticamente significativa del NT y PT (p = 0.0156 y 0.0001) con el carbono orgánico, dan cuenta de procesos que favorecerían la acumulación de carbono orgánico en los sedimentos de la margen noreste del golfo de Urabá. Los valores de SiB en los núcleos fueron muy similares y alcanzaron porcentajes medios de 0.241% en el núcleo SP 9 y de 0.245% en el SP 41.
A partir de las variaciones en las concentraciones de CaCO3, COT y PT en el núcleo SP 41, se pueden establecer tres zonas. La primera zona se caracteriza por una ligera disminución de CaCO3 desde el tope hasta los 1266 años AP, descenso que puede obedecer a un aumento de la influencia de aguas oceánicas en la margen noroeste del golfo. La segunda zona, entre 1386 y 2845 años AP, presenta un aumento continuo de CaCO3 y podría estar asociado con un debilitamiento progresivo del efecto de aguas marinas, así como por la posible intensificación en la descarga de aguas y sedimentos por el río Atrato y tributarios menores. La tercera zona, entre 2525 y 2845 años AP, se caracteriza por presentar las mayores concentraciones de CaCO3, COT y PT en el núcleo. En contraste con lo observado en el núcleo SP 41, el contenido de CaCO3 en el núcleo SP 9 no presentó variaciones importantes excepto en los intervalos 170-171 y 190-191 cm (∼764-895 años AP), los cuales son considerados como datos atípicos.
Las relaciones molares C:N y C:P del núcleo SP 9 aumentaron con la profundidad, especialmente en la zona media e inferior de la secuencia entre 222 y 633 años AP. Las relaciones molares C:N y C:P en el núcleo SP 41 mostraron tendencias contrarias y no tan claras como en el núcleo anterior. Mientras la relación C:N aumentó ligeramente, la relación C:P disminuyó con la profundidad. En este núcleo la relación C:N fluctuó entre 10.8 y 20, y la relación C:P osciló entre 17.3 y 55.6.
Registro de diatomeas
En general, las dos secuencias sedimentarias se caracterizaron por presentar baja preservación de diatomeas y sólo se logró cuantificar un número representativo de valvas en los intervalos 5-7, 10-12, 15-17, 20-22, 25-27, 30-32 y 35-37 cm del núcleo SP 41, intervalos que corresponden a los últimos ∼800 años AP. En los 41 intervalos analizados del núcleo SP 9 no se encontraron diatomeas. Para descartar la posible disolución de las valvas por el tratamiento ácido de las muestras, se observaron al microscopio sedimentos frescos y en ninguno de los casos se observó la presencia de las mismas.
Del núcleo SP 41 se identificaron en total 148 taxa de las cuales 43 se determinaron hasta género, 48 hasta nivel de especie y 4 hasta variedad. En orden decreciente, los géneros con mayor número de especies fueron Diploneis (24 especies), Navicula (13 especies) y Amphora (9 especies). Las especies más abundantes fueron Grammatophora marina (11.94%), Tryblionella sp 1 (9.52%), Diploneis nitescens (6.54%), Diploneis weissflogii (5.67%), Thalassionema nitzschioides (4.85%), Cyclotella stylorum (4.80%), Psammodictyon panduriforme (5.02%), Azpeitia nodulifer (3.97%), Lyrella spectabilis (3.78%), Pleurosigma cf. diverse-striatum (3.60%), Paralia sulcata (3.41%) y Diploneis crabro (3.14%) (Figuras 5, 6 y 7).
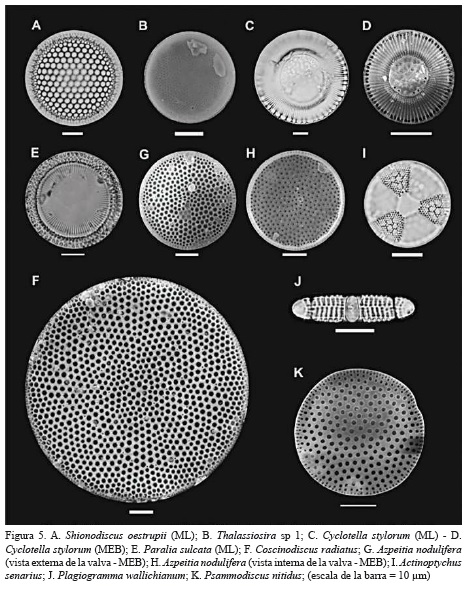

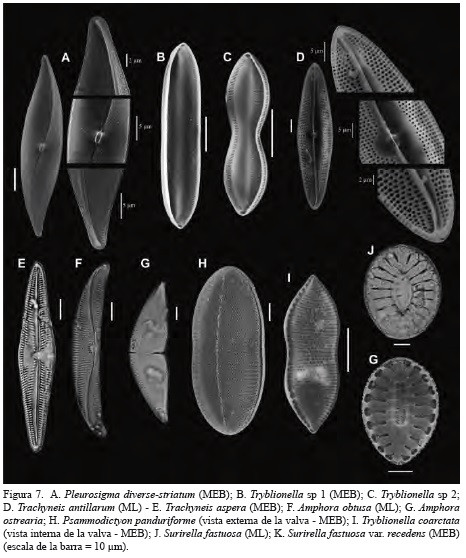
Apartir del análisis de agrupamiento se definieron dos zonas de diatomeas: la zona I (5-20 cm) se caracteriza por presentar las mayores abundancias de Tryblionella sp 1, C. stylorum, P. panduriforme y P. cf. diverse-striatum, P. sulcata y A. nodulifer. Estas especies son reemplazadas en la zona II (25-35) por especies típicamente marinas como G. marina, T. nitzschioides y L. spectabilis (Figura 8).

DISCUSIÓN
Características del agua y de los sedimentos de fondo en los sitios de extracción
El aporte de aguas fluviales, principalmente desde los ríos Atrato y León, y la interacción de sus atributos geomorfológicos, hidrodinámicos e hidroclimatológicos configuran al golfo de Urabá en un ambiente costero altamente dinámico y complejo con características típicamente estuarinas (Bernal et al., 2005; Thomas et al., 2007; Montoya, 2010). Diferencias en los perfiles de salinidad, en las tasas de sedimentación estimada, en las características litoestratigráficas y en el contenido de CaCO3, indican que los sitios de perforación donde se obtuvieron los núcleos de sedimento están afectados por procesos oceánicos y fluviales distintos. Mientras en el costado noreste del golfo se asume una fuerte influencia de la pluma del río Atrato y prevalecen procesos de transporte y acumulación de sedimentos de origen terrígeno, en el costado noroeste la presencia de fragmentos orgánicos calcáreos, un mayor contenido de CaCO3 biogénico y menores tasas de sedimentación, indican que esta parte del golfo es un ambiente donde dominan procesos de naturaleza oceánica.
Los resultados consignados en este trabajo son consistentes con los obtenidos en otros estudios, en donde señalan que el golfo de Urabá presenta marcadas diferencias en la distribución espacial de la concentración de CaCO3, la salinidad superficial del agua y en la disposición del tamaño de grano de los sedimentos de fondo (García-Valencia, 2007; Álvarez-Láinez, 2008; Montoya, 2010). García-Valencia (2007) identifica una lengua de CaCO3 que bordea la costa noroeste del golfo, desde Punta Acandí hasta isla Gloria, con concentraciones superiores a 6% en el extremo norte del golfo, y valores inferiores a 1% hacia el sur, cerca de boca Tarenas. A lo largo de toda la franja costera de este sector, entre Cabo Tiburón y Acandí, se encuentran plataformas de abrasión elevadas de origen arrecifal (Prüssmann, 2012).
En general se asume que el régimen de mareas no ejerce un efecto apreciable en la dinámica hidrológica y sedimentológica del golfo, son la dirección de la corriente superficial y el patrón de vientos los que favorecen una mayor intrusión de aguas oceánicas en la zona noroeste del golfo, y permiten que el aporte de agua y sedimentos desde el río Atrato se canalice por la margen oriental del mismo (Thomas et al., 2007; Montoya, 2010). Tal dinámica en el patrón de dispersión de sedimentos, explica la tendencia inversa en relación con las tasas de sedimentación que se estimaron (Figura 2). El aumento de las tasas de sedimentación en el núcleo de la margen noreste, particularmente en los últimos ∼140 años AP, coincide con las conclusiones de otros trabajos realizados en la región, los cuales plantean una crisis erosiva litoral en las cuencas de drenaje del río Atrato y demás tributarios. Actividades mineras y agrícolas (cultivo de banano), la desviación de cauces fluviales, la extracción intensiva de materiales de playa y la construcción antitécnica de obras de defensa son factores que aceleran los procesos erosivos en el golfo (Correa y Vernette, 2004; Thomas et al., 2007; Rúa et al., 2014).
Uitz et al. (2010) estimaron que las diatomeas son responsables de aproximadamente 80% de la producción primaria en zonas marino-costeras. De esta manera, las concentraciones de SiB determinadas en este trabajo se asumieron como una relación con la abundancia de diatomeas y, por tanto, de la paleoproductividad en el sector norte del golfo de Urabá (Ragueneau et al., 2000; Van der Weijden, 2007). Fragmentos silíceos de radiolarios y silicoflagelados también se pueden encontrar en sedimentos marinos; sin embargo la abundancia de estos organismos en el océano Atlántico es menor comparado con lo que representan las diatomeas (Lazarus, 2007; Boltovskoy et al., 2010;).
El SiB en el tope de las secuencias coincide con registros de áreas adyacentes al delta de los ríos Mississippi y Amazonas, en donde el contenido de SiB no supera 0.5% (DeMaster, 2003; Turner et al., 2004). Asimismo, el contenido de este elemento fue inferior al registrado en sistemas altamente productivos como el Pacífico Ecuatorial (entre 40 a 60% de Bsi), el Atlántico sur (10 a 20%) o el mar de Bering (5 a 10%) (Van der Weijden, 2007). Los registros anteriores sugieren que en el golfo de Urabá, existen procesos que limitan el crecimiento de diatomeas o la acumulación y preservación de SiB en los sedimentos del mismo. Dado que los sedimentos que desembocan en el golfo son transportados y canalizados predominantemente por la margen noreste, es posible que la alta cantidad de sólidos suspendidos y la subsecuente reducción en la disponibilidad de luz solar expliquen la baja abundancia de diatomeas y, en consecuencia, del contenido de sílice biogénico en sedimentos (Nedwell et al., 1999; Paerl y Justić, 2011).
Variabilidad del registro geoquímico y su asociación con algunos cambios generales en las condiciones ambientales del golfo de Urabá durante el Holoceno tardío
Aunque el número de dataciones radiocarbono (AMS 14C) son insuficientes y limitan las inferencias y correlaciones entre los núcleos, es posible describir el comportamiento temporal de algunos factores ambientales que se asume ocurrieron en zonas adyacentes a los sitios de perforación. Las diferencias en las concentraciones de CaCO3, COT y PT entre las zonas inferior y superior del núcleo SP 41, permiten definir escenarios ambientales contrastantes en el sector noroeste del golfo durante los últimos 2845 años. Estas diferencias, que se asocian con variaciones en el nivel del mar y a pulsos de descarga fluvial, demarcan el cambio de un ambiente más antiguo con mayor influencia marina, a un ambiente de mezcla más reciente dominado por aguas estuarinas. Variaciones en la abundancia de la malacofauna de este mismo núcleo, también indican una mayor influencia de aguas oceánicas entre 2845 y 1386 años AP (Ospina-Hoyos et al., 2014). En la base de la secuencia, entre 2845 y 2489 años AP, los valores de CaCO3, COT y PT indican un periodo caracterizado no solo por una relativa estabilidad y predominio de condiciones marinas sino también por el aporte de sedimentos terrestres que provienen desde el río Atrato u otros ríos que como el Acandí, Tolo, Negro y Titumate afectan, en menor medida, la margen noroeste del golfo (Nieto Oliveros, 2004). Es altamente probable que el predominio de dichas condiciones marinas esté relacionado con el aumento en el nivel del mar en el Caribe, en tanto que el aumento en el transporte de sedimentos estaría asociado a un incremento en el régimen de drenaje y, por tanto, a la prevalencia de condiciones húmedas con intensas precipitaciones en la región.
Análisis polínicos, petrográficos y sedimentológicos locales y regionales, establecen un ascenso rápido del nivel del mar entre 6000 y 4000 años AP, y una estabilización de estos niveles alrededor de 3000 y 2500 años AP, periodo en el cual se registran condiciones secas en el norte de Sur América (Ramcharan, 2004; Martínez et al., 2010; Urrego et al., 2013; Velez et al., 2014). Cambios en las geoformas litorales y de los depósitos costeros y deltaicos, dan cuenta de un retroceso de las masas de agua oceánicas en el mar Caribe colombiano desde hace 2600 años AP, periodo donde el nivel del mar se encontraba entre 2 y 3 m por encima del nivel actual (Robertson y Martínez, 1999). A partir del análisis polínico de dos núcleos de sedimento y de las características ecológicas de la vegetación local, Urrego et al. (2006) registraron un periodo húmedo de mayor inundación entre 3900 y 2700 años AP en las planicies fluviales de la cuenca media del río Atrato. Condiciones húmedas en este periodo de tiempo también se han registrado en La Guajira, en zonas adyacentes a Cartagena, en la Ciénaga Grande de Santa Marta y en el Valle del Cauca (Martínez et al., 2010; Urrego et al., 2013; Velez et al., 2013, 2014).
A partir de ∼2445 años AP hasta la actualidad, se observa un descenso gradual del CaCO3 así como valores menores de COT y PT en comparación con la zona inferior de la secuencia. Estas tendencias de disminución son interpretadas como un descenso gradual del nivel del mar y como un debilitamiento en los pulsos de descarga fluvial en el sector norte del golfo de Urabá, debido probablemente a la reconfiguración del delta y al predominio de condiciones secas en la región. Dicho descenso en el nivel del mar a partir de ∼2445 años AP, favoreció el avance del delta y una mayor influencia de la pluma de sedimentos desde el suroeste hacia la margen oriental del golfo, tal como se observa en los resultados del núcleo obtenido en la margen noreste (Nieto Oliveros, 2004).
Los valores observados del COT y C:N permiten diferenciar dos zonas en el núcleo SP 9. La tendencia de aumento de estas variables desde 300 años AP (90 cm) hasta 960 años AP (205 cm), indican procesos que favorecerían la acumulación de materia orgánica de origen terrestre durante este periodo de tiempo. Dicha acumulación de materia orgánica rica en carbono orgánico recalcitrante es consecuencia de una mayor acumulación de arcillas, afines a la materia orgánica, desde el río Atrato (Bianchi, 2007). Las fluctuaciones de COT y de C:N en el tope del núcleo (1.18-1.92%), sugerirían una alternancia de periodos de acumulación de materia orgánica de origen marino y terrestre desde hace ∼200 años AP. Sin embargo, análisis sedimentológicos más detallados realizados en este mismo núcleo, reflejan que esta zona de la secuencia está mayormente compuesta por arenas gruesas y limos finos, situación que explicaría no solo las marcadas variaciones del COT y de C:N en este periodo sino un mayor avance del delta del río Atrato en dirección noreste (Rúa et al., 2015). En estos análisis sedimentológicos la acumulación de arenas finas y limos gruesos en profundidades puntuales de las secuencias, indican eventos con intensas precipitaciones y descargas fluviales en el golfo de Urabá durante el Holoceno tardío (Rúa et al., 2015).
Los cambios en los regímenes de precipitación en el norte de Sur América durante el Holoceno están estrechamente relacionados con el desplazamiento y amplitud latitudinal de la Zona de Convergencia Intertropical (ZCIT) (Haug et al., 2001). Sin embargo, la estacionalidad de las lluvias en la región pacífica en general, y del golfo de Urabá en particular, no puede ser explicada sólo en términos de la variación estacional de la ZCIT, en parte porque fenómenos como el ENSO y la corriente de chorro superficial del oeste ("del Chocó") afectan el ciclo anual e interanual de caudales y precipitación en el occidente colombiano (Poveda y Mesa, 1999; Rueda y Poveda, 2006). Las causas y explicaciones de las variaciones hidro climatológicas y del nivel del mar en el golfo de Urabá durante el Holoceno tardío se encuentran más allá del alcance de este estudio; no obstante, nuestros resultados reflejan como el transporte de sedimentos fluviales responde a complejas interacciones entre procesos atmosféricos, oceánicos y terrestres.
Cambios ambientales de la margen noroeste del golfo de Urabá en los últimos ∼800 años AP a partir del ensamblaje de diatomeas
En general, las diatomeas que lograron ser identificadas hasta género o especie, son consistentes con registros publicados por Nelson Navarro et al. (1989) y por Navarro y Hernández-Becerril (1997) en aguas y sedimentos del mar Caribe. Del listado de diatomeas publicado por Lozano-Duque et al. (2010), se incluyen 23 especies y cuatro variedades como nuevos registros de diatomeas para el mar Caribe colombiano, en tanto que Podosira, Rhopalodia y Rhaponeis son consideradas como nuevos géneros para el área mencionada (Figura 5, 6 y 7).
Las valvas fuertemente silificadas de G. marina, P. sulcata y T. nitzschioides son más resistentes a procesos de disolución y fragmentación y, en consecuencia, son especies susceptibles de preservarse en sedimentos marinos y costeros (Tanimura, 1999; McQuoid y Nordberg, 2003; Ryu et al., 2005, 2008; ). Abrantes (1988-1991) y Resende et al. (2007) sugieren que la abundancia de estas especies son indicadoras de ambientes marino-costeros cercanos a eventos de surgencia o a ambientes que están afectados por la descarga de aguas continentales ricas en nutrientes. En sedimentos marinos, G. marina es considerada como una especie indicadora de ambientes marinos poco profundos altamente salobres, en tanto que las especies del género Tryblionella son típicas del epipelon marino (Round et al., 1990; Piskova et al., 2009).
La información ecológica disponible señala que P. sulcata es una especie ticoplanctónica que habita en zonas litorales y sublitorales altamente productivas (Zong, 1997; Gebühr, 2011). En interpretaciones paleoambientales, cambios en la abundancia de esta especie sugieren cambios en la profundidad de la columna de agua (McQuoid y Hobson, 2001; McQuoid y Nordberg, 2003). Abrantes (1988-1991), Bao et al. (1997), Bárcena y Abrantes (1998), Abrantes y Moita (1999) y Tanimura (1999) coinciden al señalar que la abundancia de T. nitzschioides en sedimentos marino-costeros está relacionada a eventos de surgencias de baja intensidad, a regiones donde la influencia de las surgencias disminuye y, en algunos casos, a áreas costeras afectadas por la descarga de ríos.
La información ecológica anterior sugiere que las altas abundancias de G. marina, T. nitzschioides y P. sulcata son resultado de aportes significativos de nutrientes al área adyacente donde se obtuvo el núcleo SP 41, aportes que estarían asociados con descargas de agua continental desde el río Atrato y cauces menores que desembocan o desembocaron en la costa noroeste del golfo. Por otra lado, la tendencia de disminución de especies mesohalobias como P. sulcata, C. stylorum, P. pandufirome y P. diverse-striatum, y el aumento de especies polihalobias como G. marina, T. nitzschioides y L. hennedyi, sugieren una ligera reducción en la salinidad del agua en la margen noroeste del golfo en los últimos 800 años aproximadamente. Este cambio en la salinidad puede ser consecuencia de un ligero aumento del efecto del río Atrato sobre el sector noroeste del golfo, tal como lo indica la tendencia de aumento del COT hacia el tope de la secuencia. El estudio histórico de mapas cartográficos de la región indica que boca Tarenas, el brazo más hacia el norte del delta, era la principal fuente de descarga hacia el golfo, lo que supondría un mayor efecto del río sobre la margen noroeste del golfo en comparación con lo que se observa en la actualidad. Desde 1930 aproximadamente, boca El Roto se constituyó en el brazo principal del río Atrato (Nieto Oliveros, 2004).
CONCLUSIONES
Al finalizar esta investigación se puede concluir que el sistema de corrientes que rige el golfo y la influencia del río Atrato favorecen procesos de depósito y acumulación de sedimentos finos de origen terrígeno en la margen noreste del golfo. No obstante, la alta cantidad de sólidos suspendidos en este sector del golfo afectan el establecimiento de las diatomeas. En contraste, la abundancia de restos de organismos calcáreos y los altos porcentajes de CaCO3 sugieren que en la margen noroeste del golfo existen condiciones predominantemente marinas.
El registro geoquímico permitió diferenciar un ambiente de mayor influencia marina con aportes importantes de sedimentos terrígenos en la margen noroeste entre 2845 y 2489 años AP. El predominio de estas condiciones estaría relacionado con un mayor nivel del mar para la época, mientras que condiciones húmedas en este mismo periodo favorecerían la capacidad de transporte y descarga de sedimentos al golfo. Desde ∼2445 años AP hasta la actualidad los valores de CaCO3, COT y PT indicaron un descenso gradual del nivel del mar y un menor efecto de los pulsos de descarga fluvial en relación con el periodo anterior. Sin embargo, el ensamblaje de diatomeas indicó una ligera activación del efecto fluvial desde hace 800 años AP. El descenso del nivel del mar favoreció la progradación deltaíca del río Atrato en dirección noreste, proceso que se habría intensificado en los últimos ∼200 años AP debido esencialmente a actividades antrópicas.
AGRADECIMIENTOS
Los autores agradecen al proyecto Universidades EAFIT, de Antioquia y Nacional - Colciencias. Erosión costera en Antioquia: dinámica sedimentaria del golfo de Urabá (erosión-depositación) durante los últimos 10000 años. Cod: 1216-452-21404, Contrato RC No. 525-2008. A Gladys Bernal, Universidad Nacional de Colombia sede Medellín por el permanente acompañamiento en la asesoría técnica y académica requerida durante el desarrollo del proyecto. A la doctora Silvia Estela Salas de la Universidad Nacional de la Plata, al doctor Yimmy Montoya Moreno y a Luis Alfonso Vidal por su ayuda en la determinación taxonómica de las diatomeas.
BIBLIOGRAFÍA
Abrantes, F. 1988. Diatom assemblages as upwelling indicators in surface sediments off Portugal. Mar. Geol., 85(1): 15-39. [ Links ]
Abrantes, F. 1991. Increased upwelling off Portugal during the last glaciation: Diatom evidence. Mar. Micropaleontol., 17(3): 285-310. [ Links ]
Abrantes, F. y M.T. Moita. 1999. Water column and recent sediment data on diatoms and coccolithophorids, off Portugal, confirm sediment record of upwelling events. Oceanologica Acta, 22(1): 67-84. [ Links ]
Affan, A., R. Karawita, Y. Jeon, B. Kim y J. Lee. 2006. Growth characteristics, bio-chemical composition and antioxidant activities of benthic diatom Grammatophora marina from Jeju coast, Korea. Algae., 21(1): 141. [ Links ]
Álvarez-Láinez, A.M. 2008. Estudio del transporte de sedimentos superficiales en el piso marino del golfo de Urabá. Tesis de Maestría, Universidad Nacional de Colombia, Medellín, 100 p. [ Links ]
Arndt, S., B. Jørgensen, D. LaRowe, J. Middelburg, R. Pancost y P. Regnier. 2013. Quantifying the degradation of organic matter in marine sediments: A review and synthesis. Earth Sci. Rev., 123: 53-86. [ Links ]
Bao, R., M. Varela y R. Prego. 1997. Mesoscale distribution patterns of diatoms in surface sediments as tracers of coastal upwelling of the Galician shelf (NW Iberian Peninsula). Mar. Geol., 144(1): 117-130. [ Links ]
Bárcena, M. y F. Abrantes. 1998. Evidence of a high-productivity area off the coast of Málaga from studies of diatoms in surface sediments. Marine Micropaleontol., 35(1): 91-103. [ Links ]
Battarbee, R.W., V.J. Jones, R.J. Flower, N.G. Cameron, H. Bennion, L. Carvalho y S. Juggins. 2001. Diatoms. En: Smol, J.P., H.J. Birks y W.M. Last (Eds.). Tracking environmental change using lake sediments: terrestrial, algal, and siliceous indicators. Springer, Países Bajos. 155-202. [ Links ]
Bernal, G., L.J. Montoya, C. Garizábal y M. Toro. 2005. La complejidad de la dimensión física en la problemática costera del golfo de Urabá, Colombia. Gestión y Ambiente, 8(1): 123-135. [ Links ]
Bianchi, T. S. 2007. Biogeochemistry of estuaries. Oxford University Press, Nueva York. 687 p. [ Links ]
Bianchi, T.S. y J.E. Bauer. 2011. Particulate organic carbon cycling and transformation. 69-117. En: Wolanski, E. y D. McLusky (Eds.). Treatise on estuarine and coastal science. Academic Press, Waltham, EE.UU. 245 p. [ Links ]
Boltovskoy, D., S.A. Kling, K. Takahashi y K. Bjørklund. 2010. World atlas of distribution of recent polycystina (Radiolaria). Palaeontol. Electron., 13(3): 230. [ Links ]
Chevillot, P., A. Molina, L. Giraldo y C. Molina. 1993. Estudio geológico e hidrológico del golfo de Urabá. Bol. Cient. CIOH., 14: 79-89. [ Links ]
Cooper, S.R. 1995. Diatoms in sediment cores from the mesohaline Chesapeake Bay, USA. Diatom Res., 10(1): 39-89. [ Links ]
Cooper, S.R. 2010. Estuarine paleoenvironmental reconstructions using diatoms. 352-373. En: Stoermer, E.F. y J.P. Smol (Eds.). The diatoms: applications for the environmental and earth sciences. Cambridge University Press, NuevaYork. 667 p. [ Links ]
Correa, I.D. y G. Vernette. 2004. Introducción al problema de la erosión litoral en Urabá (sector Arboletes-Turbo) costa Caribe colombiana. Bol. Invest. Mar. Cost., 33(1): 7-28. [ Links ]
Cremer, H., F. Sangiorgi, F. Wagner-Cremer, V. McGee, A.F. Lotter y H. Visscher. 2007. Diatoms (Bacillariophyceae) and dinoflagellate cysts (Dinophyceae) from Rookery bay, Florida, USA. Carib. J. Sci., 43(1): 23. [ Links ]
DeMaster, D. 2003. The diagenesis of biogenic silica: chemical transformations occurring in the water column, seabed, and crust. Treatise. Geochem., 7: 87-98. [ Links ]
Fritz, S.C. 2007. Salinity reconstructions from continental lakes. 514-523. En: Elias, S.A. (Ed.). Encyclopedia of Quaternary Science. Elsevier, Oxford. 3576 p. [ Links ]
García-Valencia, C. 2007. Atlas del golfo de Urabá: una mirada al Caribe de Antioquia y Chocó. Serie Publicaciones Especiales Invemar No 12, Santa Marta. 159 p. [ Links ]
Gebühr, C. 2011. Investigations on the ecology of the marine centric diatom Paralia sulcata at Helgoland Roads, North Sea, Germany. Tesis Ph.D., Jacobs University, Bremen, Alemania, 175 p. [ Links ]
Hasle, G.R., E.E. Syvertsen, K.A. Steidinger, K. Tangen y C.R. Tomas. 1996. Identifying marine diatoms and dinoflagellates. Academic Press Inc, San Diego. 598 p. [ Links ]
Haug, G.H., K.A. Hughen, D.M. Sigman, L.C. Peterson y U. Röhl. 2001. Southward migration of the intertropical convergence zone through the Holocene. Science, 293(5533): 1304-1308. [ Links ]
Invemar. 2005. Building capacity for improve adaptability to sea level rise in two vulnerable points of the Colombian coastal areas (Tumaco-Pacific coast and Cartagena-Caribbean coast) with special emphasis on human populations under poverty conditions. NCAP-Colombia, Santa Marta. 137. [ Links ]
Invemar, Corpourabá, Gobernación de Antioquia y Codechocó. 2008. Cartilla lineamientos y estrategias para el manejo integrado de la Unidad Ambiental Costera del Darién (UAC-Darién). Serie de Documento Generales del Invemar No 2, Santa Marta. 30 p. [ Links ]
Korhola, A. 2007. Data interpretation. En: Elias, S.A. (Ed.). Encyclopedia of Quaternary Science. Elsevier, Oxford. 494-507. [ Links ]
Lazarus, D. 2007. Radiolarians and silicoflagellates. En: Elias, S.A. y C.J. Mock (Eds.). Encyclopedia of Quaternary Science (Second Edition). Elsevier, Ámsterdam. 1682-1692. [ Links ]
Lozano-Duque, Y., L.A. Vidal y G. Navas. 2010. Listado de diatomeas (Bacillariophyta) registradas para el mar Caribe colombiano. Bol. Invest. Mar. Cost., 39(1): 83-116. [ Links ]
Mackay, A.W., V.J. Jones y R.W. Battarbee. 2005. Approaches to Holecene climate reconstruction using diatoms. 297-305. En: Mackay, A.W., V.J. Jones, R.W. Battarbee, J. John Birks y F. Oldfield (Eds.). Global change in the Holocene. Hodder Arnold Publication, Nueva York. 528 p. [ Links ]
Martínez, J.I., Y. Yokoyama, A. Gómez, A. Delgado, H. Matsuzaki y E. Rendón. 2010. Late Holocene marine terraces of the Cartagena region, southern Caribbean: The product of neotectonism or a former high stand in sea-level? J. South Amer. Earth Sci., 29(2): 214-224. [ Links ]
McQuoid, M. y L. Hobson. 2001. A Holocene record of diatom and silicoflagellate microfossils in sediments of Saanich Inlet, ODP Leg 169S. Mar. Geol., 174(1): 111-123. [ Links ]
McQuoid, M. y K. Nordberg. 2003. The diatom Paralia sulcata as an environmental indicator species in coastal sediments. Estuar., Coastal Shelf Sci., 56(2): 339-354. [ Links ]
Molina, A., C. Molina y P. Chevillot. 1992. La percepción remota aplicada para determinar la circulación de las aguas superficiales del golfo de Urabá y las variaciones de su línea de costa. Bol. Cient. CIOH, 11: 43-58. [ Links ]
Montoya, L.J. 2010. Dinámica oceanográfica del golfo de Urabá y su relación con los patrones de dispersión de contaminantes y sedimentos. Tesis de Doctorado, Universidad Nacional de Colombia, Medellín. 254 p. [ Links ]
Mortlock, R.A. y P.N. Froelich. 1989. A simple method for the rapid determination of biogenic opal in pelagic marine sediments. Deep Sea Res. Part A. Oceanogr. Res. Papers, 36(9): 1415-1426. [ Links ]
Navarro, J.N. y D.U. Hernández-Becerril. 1997. Check-list of marine diatoms from the Caribbean Sea. Universidad Autónoma de México, México D.F. 48 p. [ Links ]
Nedwell, D., T. Jickells, M. Trimmer y R. Sanders. 1999. Nutrients in estuaries. Adv. Ecol. Res., 29: 43-92. [ Links ]
Nelson Navarro, J., C. Pérez, N. Arce y B. Arroyo. 1989. Benthic marine diatoms of Caja de Muertos Island, Puerto Rico. Nova Hedwigia, 49(3-4): 333-367. [ Links ]
Nieto Oliveros, M.J. 2004. Estudio morfodinámico del delta del río Atrato, golfo de Urabá, a partir de cartografía histórica y percepción remota. Trabajo de Grado Geología, Universidad Nacional de Colombia, Bogotá. 121 p. [ Links ]
Ospina-Hoyos, J., J. Palacio-Baena y L. Vásquez-Bedoya. 2014. ¿Responden los micromoluscos a los cambios ambientales durante el Holoceno tardío en el sur del mar Caribe colombiano? Universitas Scientiarum, 19(3): 233-246. [ Links ]
Paerl, H.W. y D. Justić. 2011. Primary producers: phytoplankton ecology and trophic dynamics in coastal waters. 23-42. En: Wolanski, E. y D. McLusky (Eds.). Treatise on estuarine and coastal science. Academic Press, Waltham, EE.UU. 304 p. [ Links ]
Piskova, A., T. Grygar, J. Vesela y H. Oberhaensli. 2009. Diatom assemblage variations in the Aral sea core C2/2004 over the past two millennia. Fottea, 9(2): 333-342. [ Links ]
Poveda, G. y O. Mesa. 1999. La corriente de chorro superficial del Oeste ("del Chocó") y otras dos corrientes de chorro en Colombia: climatología y variabilidad durante las fases del ENSO. Rev. Acad. Col. Cienc., 23(89): 517-528. [ Links ]
Prüssmann, J. 2012. Nuevos elementos para el manejo integrado de la región de Urabá, costa Caribe colombiana. Base de datos espacial geomorfológica de la franja litoral de los departamentos Antioquia y Chocó. Tesis de Maestría, Universidad EAFIT, Medellín. 108 p. [ Links ]
Quiceno, M. 2008. Patrones de variabilidad climática y oceánica en la zona costera del río Sinú y su señal en registros naturales lacustres y coralinos. Trabajo de Maestría, Universidad Nacional de Colombia, Medellín. 120 p. [ Links ]
Ragueneau, O., P. Tréguer, A. Leynaert, R. Anderson, M. Brzezinski, D. DeMaster, R. Dugdale, J. Dymond, G. Fischer y R. Francois. 2000. A review of the Si cycle in the modern ocean: recent progress and missing gaps in the application of biogenic opal as a paleoproductivity proxy. Global Planet Change, 26(4): 317-365. [ Links ]
Ramcharan, E.K. 2004. Mid-to-late Holocene sea level influence on coastal wetland development in Trinidad. Quatern. Internat., 120(1): 145-151. [ Links ]
Resende, P., U. M. Azeiteiro, F. Gonçalves y M. J. Pereira. 2007. Distribution and ecological preferences of diatoms and dinoflagellates in the west Iberian coastal zone (North Portugal). Acta Oecolog., 32(2): 224-235. [ Links ]
Restrepo, J. y B. Kjerfve. 2004. The Pacific and Caribbean rivers of Colombia: water discharge, sediment transport and dissolved loads. 169-187. En: Lacerda L., R. Santelli, E. Duursma y J. Abrao (Eds.). Environmental geochemistry in tropical and subtropical environments. Springer, Berlín. 385 p. [ Links ]
Robertson, K. y N. Martínez. 1999. Cambios del nivel del mar durante el Holoceno en el litoral Caribe colombiano. Cuad. Geogr., 8(1): 168-198. [ Links ]
Rosales, L. 1979. Manual de laboratorio de oceanografía química. Anales del Instituto de Ciencias del Mar y Limnología, UNAM, México D.F. 203 p. [ Links ]
Round, F.E., R.M. Crawford y D.G. Mann. 1990. The diatoms: biology and morphology of the genera. Cambridge University Press, Oxford. 747 p. [ Links ]
Rúa, A., G. Liebezeit y J. Palacio-Baena. 2014. Mercury colonial footprint in Darién Gulf sediments, Colombia. Environ. Earth Sci., 71(4): 1781-1789. [ Links ]
Rúa, A., G. Liebezeit, R. Molina y J. Palacio. 2015. Unmixing progradational sediments in a southwestern Caribbean gulf through late Holocene: backwash of low-level atmospheric jets. J. Coast. Res., 32(2): 397-407. [ Links ]
Rueda, O. y G. Poveda. 2006. Variabilidad espacial y temporal del chorro del "Chocó" y su efecto en la hidroclimatología de la región del pacífico colombiano. Meteorol. Col., 10: 132-145. [ Links ]
Ryu, E., S. Yi y S.J. Lee. 2005. Late Pleistocene-Holocene paleoenvironmental changes inferred from the diatom record of the Ulleung Basin, East Sea (Sea of Japan). Mar. Micropaleontol., 55(3-4): 157-182. [ Links ]
Ryu, E., S.J. Lee, D.Y. Yang y J.Y. Kim. 2008. Paleoenvironmental studies of the Korean peninsula inferred from diatom assemblages. Quatern. Internat., 176: 36-45. [ Links ]
Seiter, K., C. Hensen, J. Schröter y M. Zabel. 2004. Organic carbon content in surface sediments-defining regional provinces. Deep Sea Res. Part I: Oceanogr. Res. Pap., 51(12): 2001-2026. [ Links ]
Shiono, M. y I. Koizumi. 2002. Taxonomy of the Azpeitia nodulifera group in late Neogene sediments from the northwest Pacific Ocean. Diatom Res., 17(2): 337-361. [ Links ]
Slomp, C.P. 2011. Phosphorus cycling in the estuarine and coastal zones: Sources, sinks, and transformations. 201-229. En: Wolanski, E. y D. McLusky (Eds.). Treatise on estuarine and coastal science. Academic Press, Waltham, EE.UU. 362 p. [ Links ]
Statham, P.J. 2012. Nutrients in estuaries - An overview and the potential impacts of climate change. Sci. Total Environ., 434(0): 213-227. [ Links ]
Sterrenburg, F.A.S. 2001. Studies on the Genera Pleurosigma and Gyrosigma: The types of Shadbolt and related taxa. Proc. Acad.Nat.Sci. Phila., 151(1): 121-127. [ Links ]
Stidolph, S., F. Sterrenburg, K. Smith y A. Kraberg. 2012. Stuart R. Stidolph diatom atlas. U.S. Geological Survey, Reston, EE.UU. 199 p. [ Links ]
Strickland, J. y T. Parsons. 1972. A practical handbook of seawater analysis. Fisheries Research Board of Canada, Ottawa. 310 p. [ Links ]
Tanimura, Y. 1999. Varieties of a single cosmopolitan diatom species associated with surface water masses in the North Pacific. Mar. Micropaleontol., 37(2): 199-218. [ Links ]
Thomas, Y., M. Cesaraccio, C. García y L. Ménanteau. 2007. Contribución de la hidrografía histórica al estudio de la cinemática de los fondos marinos: evolución del golfo de Urabá, Colombia. Bol. Cient. CIOH, 25: 110-119. [ Links ]
Turner, R.E., C. Milan y N. Rabalais. 2004. A retrospective analysis of trace metals, C, N and diatom remnants in sediments from the Mississippi River delta shelf. Mar. Poll. Bull., 49(7): 548-556. [ Links ]
Uitz, J., H. Claustre, B. Gentili y D. Stramski. 2010. Phytoplankton class specific primary production in the world's oceans: Seasonal and interannual variability from satellite observations. Global Biogeochem. Cycles, 24(3): 1-19. [ Links ]
Urrego, L.E., L.A. Molina, D.H. Urrego y L.F. Ramírez. 2006. Holocene space-time succession of the Middle Atrato wetlands, Chocó biogeographic region, Colombia. Palaeogeog., Palaeoclimatol., Palaeoecol., 234(1): 45-61. [ Links ]
Urrego, L.E., A. Correa-Metrio, C. González, A.R. Castaño y Y. Yokoyama. 2013. Contrasting responses of two Caribbean mangroves to sea-level rise in the Guajira Peninsula (Colombian Caribbean). Palaeogeogr., Palaeoclimatol., Palaeoecol., 370: 92-102. [ Links ]
Van der Weijden, C.H. 2007. Cahiers of geochemistry silica I: Silicon analytical, physical and terrestrial geochemistry. http://www.geo.uu.nl/Research/Geochemistry/Cahiers/Silica%20I%20integral%20text.pdf. 15/01/2013. [ Links ]
Vélez, M.I., J.I. Martínez y F. Suter. 2013. Late Holocene history of the floodplain lakes of the Cauca River, Colombia. J. Paleolimnol., 49(4): 591-604. [ Links ]
Vélez, M.I., J. Escobar, M. Brenner, O. Rangel, A. Betancourt, A.J. Jaramillo, J.H. Curtis y J.L. Moreno. 2014. Middle to late Holocene relative sea level rise, climate variability and environmental change along the Colombian Caribbean coast. The Holocene: 0959683614534740. [ Links ]
Walkley, A. y I.A. Black. 1934. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci., 37(1): 29-38. [ Links ]
Willis, K.J. y H.J.B. Birks. 2006. What is natural? The need for a long-term perspective in biodiversity conservation. Science, 314(5803): 1261-1265. [ Links ]
Witkowski, A., H. Lange-Bertalot y D. Metzeltin. 2000. Diatom flora of marine coasts I. Iconographia diatomologica, 7: 1-925. [ Links ]
Witon, E. y A. Witkowski. 2006. Holocene diatoms (Bacillariophyceae) from Faeroe Islands fjords, Northern Atlantic Ocean. II. Distribution and taxonomy of marine taxa with special reference to benthic forms. Diatom Res., 21(1): 175-215. [ Links ]
Zong, Y. 1997. Implications of Paralia sulcata abundance in Scottish isolation basins. Diatom Res., 12(1): 125-150. [ Links ]
RECIBIDO: 05/01/2015 ACEPTADO: 12/04/2016

