










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.45 no.1 Santa Marta Jan./June 2016
FECUNDIDAD DEL JUREL ALETA AMARILLA, CARANX HIPPOS (LINNAEUS) EN EL CARIBE COLOMBIANO
FECUNDITY OF YELLOW FIN MACKEREL, CARANX HIPPOS (LINNAEUS) IN THE COLOMBIAN CARIBBEAN
Sandra Alfaro-Martínez1, Diana Bustos-Montes2*, Sarith Salas-Castro3, Javier Gómez-León3 y Mario Rueda3
1 Universidad del Magdalena, Facultad de Ciencias Básicas, Biología, Santa Marta, Colombia. salmaalfaro111@gmail.com.
2 Instituto de Estudios en Ciencias del Mar (Cecimar), Universidad Nacional de Colombia sede Caribe, Playa Salguero, Santa Marta, Colombia. Calle 25 # 2-55. Playa Salguero, Santa Marta, Colombia. Tel: +57- 5 - 432 86 00 ext. 180. dmbustosmo@unal.edu.co (*autor de correspondencia).
3 Instituto de Investigaciones Marinas y Costeras (Invemar). Santa Marta, Colombia. sarith.salas@invemar.org.co, javier.gomez@invemar.org.co, mario.rueda@invemar.org.co.
RESUMEN
Caranx hippos es uno de los recursos ícticos marinos más importantes en Colombia, especialmente en la pesca artesanal. Con el propósito de profundizar en su conocimiento biológico, se estimó la fecundidad parcial y el diámetro de los ovocitos. Las muestras provenían de faenas artesanales comerciales. A cada individuo se le midió longitud total, el peso total y el peso gonadal. La fecundidad se estimó combinando el método gravimétrico con análisis de imágenes, usando el software de libre acceso Image J®. A partir de la medición de 149523 ovocitos se obtuvo un diámetro promedio de 0.32 ± 1.24E-4 mm, y valores de fecundidad variables con un ámbito entre 291439 y 4380480 ovocitos/gónada. No se evidenció una relación directa entre la cantidad y el diámetro de los huevos con la talla y el peso del pez, lo que sugiere que no siempre las hembras más grandes son las más fecundas. Se describe el desarrollo microestructural de los oocitos maduros, encontrando diámetros promedio en alvéolo cortical y vitelogénesis de 98.6 + 33.1 µm y 207.5+ 59.6 µm, respectivamente. Dada la importancia en el país de C. hippos, este trabajo constituyó un importante aporte al conocimiento de la especie, que puede ser usado en modelos pesqueros con aplicabilidad directa en medidas de manejo.
PALABRAS CLAVE: Caranx hippos, Histología, Método gravimétrico, Diámetro ovocitario, Costa Caribe.
ABSTRACT
Caranx hippos is one of the most important fish resources in Colombia, particularly in artisanal fisheries. In order to deepen C. hippos biological knowledge, fecundity (number of eggs in development) and oocyte diameter was estimated. Fish samples were collected from commercial artisanal fishing trips, each fish was measured (total length, Lt) and weighted (total weight, Pt); gonads were removed, weighed and analyzed for determining fecundity using a gravimetric method combined with image analysis performed using the open access software Image J®. From the measurement of 149552 oocytes, an average diameter of 0.32 ± 1.24E-4 mm, was obtained, while fecundity varied between 291439 and 4380480 oocytes per gonad. No direct relationship was evidenced between egg number and diameter, and fish size and weight. Mature oocyte microstructural development is described, average diameters in cortical alveoli and vitellogenesis were 98.6 + 33.1 µm y 207.5+ 59.6 µm, respectively. This result suggests that larger females are not always the more fertile. Given the importance of C. hippos in Colombia, this work is a valuable contribution to the knowledge of the species, which can be used in fishery models with direct applicability to management procedures.
KEY WORDS: Caranx hippos, Histology, Gravimetric method, Oocyte diameter, Caribbean coast.
INTRODUCCIÓN
El jurel aleta amarilla, Caranx hippos (Linnaeus), es una especie que puede alcanzar 1 m de longitud total y más de 25 kg de peso total (Cervigón, 1993). Es uno de los recursos pesqueros de gran importancia para la economía y sostenimiento de la pesca artesanal del Caribe colombiano (Caiafa, 2013). En esta costa, presentó altos volúmenes de desembarco en los últimos años (9.2% del total de los desembarques en 2014; AUNAP-UNIMAGDALENA, 2015) y se considera que durante la última década ha sido sometido a una explotación acelerada y destructiva (Caiafa et al., 2011). Por otro lado, el conocimiento de la biología de las especies y específicamente la comprensión de los procesos de reproducción puede ser una herramienta para el manejo y conservación de los recursos, ya que permite el entendimiento de mecanismos que influyen en su comportamiento, subsistencia y migraciones (Alarcón et al., 2004).
En Colombia, son pocos los estudios en biología reproductiva o afines dirigidos a C. hippos. Pinilla (1986) menciona características biológicas de la especie, dentro de las que se destacan las variaciones del estado gonadal y distribuciones de tallas y sexo. Posada y Sandoval (2007) estudiaron durante un año aspectos reproductivos del jurel aleta amarilla en Bocas de Ceniza (Atlántico), estimando fecundidades absoluta y relativa para la especie entre 3304700 y 4975000 huevos por gónada y 456 y 904 huevos por gramo de peso, respectivamente. Concluyen que C. hippos cumple un ciclo reproductivo anual con dos períodos de desove en enero-febrero y junio-julio y que es un desovador asincrónico continuo multiporcional, que libera un alto número de ovocitos en cada desove como estrategia reproductiva para la rápida reposición de individuos a la población. Otros estudios, como los de Palacio (2009), Restrepo (2010), Guerrero (2011) y Posada-Peláez et al. (2012), mencionan que es uno de los recursos ícticos más importantes en la región Caribe y describen la progresión de los estados de madurez para inferir la actividad reproductiva. Por lo anterior y considerando la importancia económica de jurel aleta amarilla, este estudio tiene como objetivo determinar la fecundidad parcial de C. hippos en el Caribe colombiano, describir microestructuralmente la fase de madurez avanzada de desarrollo ovocitario, además de establecer la posible relación entre el tamaño y número de los ovocitos y el tamaño y peso de las hembras maduras.
ÁREA DE ESTUDIO
La costa Caribe continental colombiana ubicada en la parte extrema del noroccidente de Suramérica, limita al este con Venezuela en Castilletes en la desembocadura del río Sillamaná (11° 50' N; 71° 18'W) y al oeste con Panamá en Cabo Tiburón (08° 42' N; 77° 19'W) y tiene una longitud de 1642 km (MADR y CCI, 2006). Aunque existen varios sitios de desembarco de pesca artesanal en el Caribe continental colombiano, para la obtención de muestras de este trabajo fueron escogidos Las Flores (departamento de Atlántico) y Ciénaga (departamento de Magdalena), por ser lugares donde tradicionalmente se han registrado altos volúmenes de desembarco de C. hippos de grandes tallas (Posada y Sandoval, 2007; Caiafa et al., 2011; Posada et al., 2012; Caiafa, 2013).
MATERIALES Y MÉTODOS
Fase de campo
Las muestras provinieron de faenas comerciales de pesca artesanal que tuvieron lugar entre septiembre de 2013 y junio de 2014. Se obtuvieron en total 46 hembras, a las que se les tomaron medidas de longitud y peso total. Se realizó una disección, se obtuvo el peso gonadal y se determinó el grado de madurez a nivel macroscópico utilizando la escala de cinco estadios propuesta por Holden y Raitt (1974) que se resume así: I: inmaduro; II: virgen madurando y recuperando; III: Madurando; IV: Maduro y V: Desovado.
Análisis de laboratorio
El sexo y la madurez sexual fueron corroborados microscópicamente a través de técnicas histológicas con los caracteres propuestos por Posada y Sandoval (2007), Guerrero (2011) y Posada et al. (2012). Se realizaron mediciones de los ovocitos en estado alvéolo cortical y vitelogenados, mediante el programa de libre acceso Govocitos®, además de obtener un registro fotográfico y realizar la descripción histológica de cada etapa de desarrollo encontrada. Para esto fueron analizadas 92 fotografías, correspondientes a cuatro campos escogidos al azar del corte histológico transversal de 23 ovarios.
Para la estimación de la fecundidad parcial se seleccionaron 30 ovarios maduros (estados III y IV), realizando el conteo y medición de los ovocitos a través de la combinación del método gravimétrico tradicional (Holden y Raitt, 1974) con el análisis de imágenes (Klibansky y Juanes, 2008; Camacho, 2011; Bustos-Montes et al., 2015) usando el software de libre acceso Image J®. De acuerdo con un análisis previo se decidió que el intervalo de tamaño de los ovocitos que serían medidos y contabilizados era el comprendido entre 0.23 y 0.39 mm, ámbito en el cual se encontraban en avanzado estado de madurez.
Análisis de información
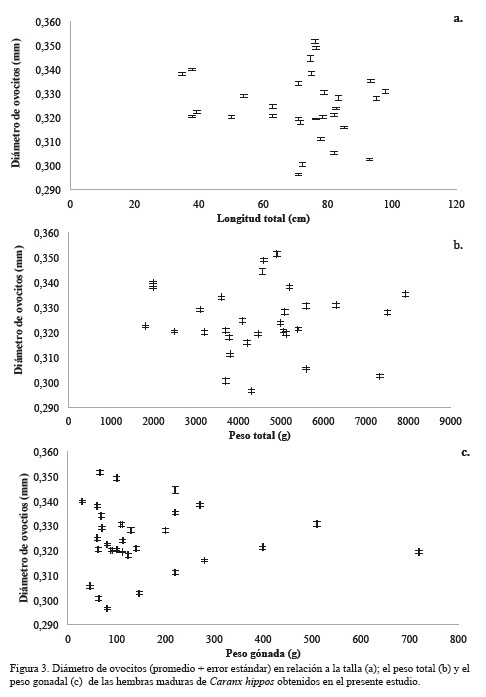
La fecundidad parcial (número de ovocitos en estado avanzado de madurez), se determinó con la ecuación propuesta por Lucano-Ramírez et al. (2005):
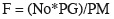
Donde No es el número de ovocitos contabilizados en la muestra, PG el peso de la gónada y PM el peso de la muestra gonadal (que en todos los casos fue de 0.50 g). Para la estimación del diámetro ovocitario, se midió la longitud de los ejes mayor y menor de cada uno de los ovocitos y se aplicó la siguiente ecuación (Camacho, 2011):
Diámetro ovocito (mm)= [(longitud eje mayor (mm)* longitud eje menor (mm)]0,5
En todos los casos se usaron como medidas de tendencia central y variabilidad el promedio y el error estándar, respectivamente. Para definir la función que mejor explicaba la relación entre las variables morfométricas (longitud total, peso total y peso gonadal) con la fecundidad parcial y el diámetro ovocitario se probaron modelos de regresión lineal, potencial y exponencial, que fueron comparados entre sí a través del coeficiente de determinación R2, el estadístico F y su nivel de significancia (Guisande-González et al., 2006).
RESULTADOS
Los peces (n = 46) midieron entre 35 y 110 cm con promedio de 74.66 ± 2.48 cm, presentándose la mayoría entre 70 y 85 cm (58.70% del total). El peso promedio total de los peces era de 4.91 ± 0.24 kg, siendo el pez menos pesado de 1.80 kg y el mayor de 9.00 kg. Se obtuvieron 46 gónadas, tres madurando (estado III; 7%), 31 maduras (estado IV; 67%) y 12 desovadas (estado V; 26%). Solo dos gónadas provenían de Ciénaga, por lo que para la estimación de fecundidad se usaron solo las 30 gónadas maduras (estado IV) de Las Flores. El peso de las gónadas de dicha submuestra varió entre 30 y 720 g (promedio de 162.95 ± 27.57 g).
En la fase de alvéolo cortical, los ovocitos midieron en promedio 98.6 ± 33.1 µm (n = 245). Se caracterizaron por presentar hacia la periferia de la célula alvéolos corticales, que tiene la apariencia de estructuras huecas e indican el inicio del proceso de maduración del pez; a medida que avanza el desarrollo se distribuyen homogéneamente en el citoplasma hasta ubicarse en la periferia de la envoltura nuclear. El citoplasma contiene material basófilo de apariencia granular. En el núcleo eosinófilo los nucléolos se ubican en el borde (Figura 1a).
En la fase de vitelogénesis, se observan glóbulos de vitelo ubicados en la periferia migrando hacia el núcleo y cercanos a éste se ubican algunos alvéolos corticales. Posteriormente, los gránulos de vitelo se fusionan unos con otros y esta coalición da lugar a una masa lipídica que se distribuye por todo el citoplasma ocasionando el aumento considerable en diámetro del ovocito (207.5 + 59.6 µm; n = 745). El núcleo con contenido eosinófilo comienza a migrar progresivamente hacia el polo animal de la célula y los nucléolos disminuyen su tamaño. En algunos ovocitos se pueden observar gotas lipídicas de gran tamaño que se fusionan ocupando gran parte del citoplasma (Figura 3b).
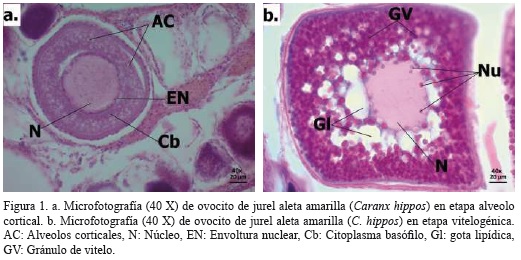
El diámetro promedio de los 149523 ovocitos estudiados gravimétricamente fue de 0.32 ± 1.24E-4 mm, siendo el más pequeño de 0.22 mm y el más grande de 0.40 mm. El valor promedio para la fecundidad parcial fue de 1446141 ± 212068 ovocitos por gónada, la mayor cantidad (4380480 ovocitos por gónada) se contabilizó en una hembra de 71 cm de Lt, Pt de 5.12 kg y peso gonadal de 0.72 kg, en tanto que la menor cantidad (291439 ovocitos por gónada) en una hembra de 78.5 cm de Lt, Pt de 5.06 kg y peso gonadal de 0.06 kg.
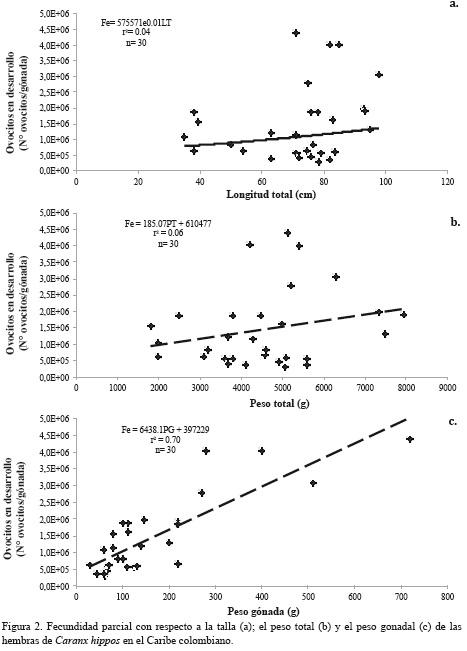
La relación entre la fecundidad parcial y la longitud se ajustó a un modelo exponencial (Figura 2a; r2 = 0.04, F1,28 = 4.6, p = 0.04), mientras que la relación con el peso total (Figura 2b; r2 = 0.06, F1,28 = 4.8, p = 0.04) siguió un modelo lineal. En ambos casos el ajuste al modelo no es tan bueno, como lo muestran los estadísticos calculados. Por otro lado, se encontró relación significativa de ajuste lineal entre la fecundidad y el peso gonadal (Figura 2c; r2 = 0.7, F1,28 = 35.0, p < 0.001); sin embargo, no se encontró tendencia significativa entre el diámetro promedio de los ovocitos y las variables morfométricas (Figura 3).
DISCUSIÓN Y CONCLUSIONES
Como pudo observarse en el presente estudio, la variación de la fecundidad entre individuos de la misma especie y de la misma población es muy amplia y en condiciones normales puede diferir hasta en un factor de diez entre un individuo y otro (Csirke, 1980). Varios fenómenos pueden afectar la fecundidad de un grupo de peces e incluso de toda la población, tales como la disponibilidad de alimento, las condiciones del medio ambiente y el nivel de saturación de la población, entre otros (Csirke, 1980; Murua et al., 2003). Los cambios en la fecundidad están estrechamente ligados con la edad o el tamaño de los individuos (Cerna y Oyarzún, 1998; Marteinsdottir y Begg, 2002; Urriola et al., 2004; Murua et al., 2006; Cooper et al., 2013), característica que no se pudo corroborar en el presente estudio, ya que no siempre las hembras más grandes y pesadas produjeron una mayor cantidad de huevos o los huevos más grandes.
En cuanto a la descripción histológica de los tipos de ovocitos (alvéolos corticales, vitelogénicos), coincide con las características mencionadas por Posada y Sandoval (2007), Guerrero (2011) y Posada-Peláez et al. (2012). La organización histológica del ovario permitió confirmar que el desarrollo ovocitario es de tipo asincrónico multiproporcional, y sugiere que la fecundidad es indeterminada y se presentan múltiples desoves, como afirman Posada y Sandoval (2007) y Bustos-Montes et al. (2015), lo que implica una mayor variación intraespecífica en la capacidad reproductiva.
El diámetro promedio de los ovocitos en estado de desarrollo alvéolo cortical y en vitelogénesis encontrado en el presente estudio es mucho más cercano al registrado por Posada y Sandoval (2007) que al estimado por Posada-Peláez et al. (2012). Estas diferencias pueden ser explicadas por las variaciones temporales y espaciales. Los muestreos de las tres investigaciones se realizaron en años diferentes. Además, el área de muestreo de Posada-Peláez et al. (2012) fue el departamento del Magdalena, principalmente en el golfo de Salamanca, mientras que las muestras del presente estudio, junto con las de Posada y Sandoval (2007), provenían de áreas de pesca aledañas a Bocas de Ceniza, desembocadura del río Magdalena en el mar Caribe. Por otro lado, es importante que la fecundidad sea estimada analizando gónadas cercanas al desove y en aras de comparar valores interanuales, los muestreos deberían ser en la misma época del año, intentando contrastar las mismas etapas de desarrollo de la maduración (Ganias et al., 2014). Lo anterior no fue posible, ya que las investigaciones no estuvieron relacionadas y en las publicaciones anteriores no se detalla el mes en el que fueron capturados los individuos que fueron usados para las mediciones.
La variabilidad en la producción (cantidad) y calidad (tamaño) ovocitaria en individuos de la misma especie ha sido investigada, concluyendo que la condición maternal es determinante en el éxito reproductivo y esta a su vez puede verse afectada por factores ambientales y antrópicos (Saborido-Rey, 2004; Alonso-Fernández y Saborido-Rey, 2011; Cooper et al., 2013; Ganias et al., 2014, 2015). Altos niveles de explotación pesquera pueden modificar la estructura de tallas de la población haciendo que el potencial reproductivo del stock disminuya desproporcionadamente (Berkeley et al., 2004; Wright y Trippel, 2009) y esos cambios impactan considerablemente las estimaciones del estado anual de la fecundidad (Cooper et al., 2013). La fuerte presión por pesca a la que está sometido el jurel en el Caribe colombiano, se refleja en el acelerado aumento y posterior descenso en los desembarcos durante los últimos 10 años (AUNAP-Unimagdalena. 2015) y en que más de 75% de los peces se capturen inmaduros (Altamar et al., 2015). Dicha presión podría estar impactando la condición de los parentales y por tanto la capacidad reproductiva de la especie, lo que explicaría la reducción en la estimación de fecundidad individual del presente estudio con la registrada por Posada y Sandoval (2007), quienes calcularon valores entre 3304700 y 4975000 ovocitos por gónada.
Desde el punto de vista genético, Caiafa (2013) afirma que C. hippos en el Caribe colombiano se compone de una sola población panmíctica, es decir no estructurada y en la que existe apareamiento aleatorio, donde coexisten dos linajes que no muestran una diferenciación ligada con zonas geográficas. Al confirmar que C. hippos presenta variaciones espacio-temporales marcadas tanto en la producción como en el tamaño de ovocitos, que pueden afectar la biomasa poblacional, se recomienda que, al incluir la estimación periódica de la fecundidad con fines de manejo pesquero, se realice considerando las zonas geográficas en donde habita el recurso.
AGRADECIMIENTOS
Los autores quieren expresar sus agradecimientos a Colciencias y el programa "Valoración y Aprovechamiento de Recursos Marinos - VAR del Instituto de Investigaciones Marinas y Costeras (Invemar) por la financiación de la investigación. De manera muy especial a la comunidad de pescadores artesanales del sitio de desembarco de Las Flores (Barranquilla), al personal encargado de recoger la información en campo y a los auxiliares del laboratorio. Así mismo a los revisores anónimos por sus sugerencias y aportes.
BIBLIOGRAFÍA
Alarcón, C., L. Cubillos y C. Oyarzún. 2004. Influencia del tamaño de la hembra en la duración e intensidad de la actividad reproductiva de Merluccius gayi gayi en la zona centro-sur de Chile. Invest. Mar. Valparaíso, 32(2): 59-69. [ Links ]
Alonso-Fernández, A. y F. Saborido-Rey. 2011. Influencia maternal en la variación del ciclo reproductivo de Trisopterus luscus (Gadidae). Cienc. Mar., 37(4b): 619-632. [ Links ]
Altamar, J., L. Manjarrés-Martínez, L.O. Duarte, F. Cuello y F. Escobar-Toledo. 2015. ¿Qué tamaños deberíamos pescar? Autoridad Nacional de Acuicultura y pesca (AUNAP) -Universidad del Magdalena, Santa Marta, 54 p. [ Links ]
AUNAP y Unimagdalena. 2015. SEPEC-Boletín estadístico enero-diciembre de 2014. Convenio Autoridad Nacional de Acuicultura y Pesca y la Universidad del Magdalena, Santa Marta. 32 p. [ Links ]
Berkeley, S.A., M.A. Hixon, R.J. Larson y M.S. Love. 2004. Fisheries sustainability via protection of age structure and spatial distribution of fish populations. Fisheries, 29: 23-32. [ Links ]
Bustos-Montes, D., M. Rueda, J. Gómez-León, S. Salas, E. Cárdenas, A. Girón, J. Eraso, S. Alfaro-Mz, G. Angulo y V. Weber. 2015. Potencial reproductivo de seis recursos marinos de importancia comercial en la pesca artesanal de Colombia: herramienta para su manejo y conservación. PRY-VAR-002-13-ITF. Santa Marta. 53 p.+ anexos. [ Links ]
Caiafa, H.I. 2013. Evaluación de la variación y estructura genética del Jurel Caranx hippos (Pisces: Carangidae) en el Caribe colombiano. Tesis M.Sc. Biol., Univ. Nacional de Colombia sede Caribe. Santa Marta. 46 p. [ Links ]
Caiafa, H.I., J.C. Narváez y S. Borrero. 2011. Algunos aspectos de la dinámica poblacional del jurel Caranx hippos (Pisces: Carangidae) en Bocas de Ceniza, Caribe colombiano. Rev. MVZ Córdoba, 16(1): 2324-2335. [ Links ]
Camacho, L. 2011. Estimación preliminar de la fecundidad en algunas especies de importancia comercial (familias Carangidae y Scombridae) en el departamento del Magdalena, Caribe colombiano. Trabajo grado Biol. Mar., Univ. Jorge Tadeo Lozano, Santa Marta. 111 p. [ Links ]
Cerna, J.F. y C. Oyarzún. 1998. Talla de primera madurez sexual y fecundidad parcial de la merluza común (Merluccius gayi, Guichenot 1848) del área de la pesquería industrial de la zona de Talcahuano, Chile. Invest. Mar., Valparaíso, 26: 31-40. [ Links ]
Cervigón, F. 1993. Los peces marinos de Venezuela. Vol II. 2a. Ed. Los Roques, Caracas. 497 p. [ Links ]
Cooper W.T., L. Barbieri, M.D. Murphy y S. Lowerre-Barbieri. 2013. Assessing stock reproductive potential in species with indeterminate fecundity: Effects of age truncation and size-dependent reproductive timing. Fish. Res., 138: 31-41. [ Links ]
Csirke, J. 1980. Introducción a la dinámica de poblaciones de peces. FAO Doc. Tec. Pesca., 192: 82 p. [ Links ]
Ganias K., H. Murua, G. Claramunt, R. Dominguez-Petit, P. Gonçalves, F. Juanes, J. Keneddy, N. Klibansky, M. Korta, Y. Kurita, S. Lowerre-Barbieri, G. Macchi, M. Matsuyama, A. Medina, C. Nunes, G. Plaza, R. Rideout, S. Somarakis, A. Thorsen, A. Uriarte y M. Yoneda. 2014. Chapter 4: Egg production. En: Domínguez-Petit, R., H. Murua, F. Saborido-Rey y E. Trippel. 2014. Handbook of applied fisheries reproductive biology for stock assessment and management. Vigo, España. 109 p. [ Links ]
Ganias, K., M. Rakka, E. Mantzouki, T. Vavalidis, M. Tsinganis y C. Nunes. 2015. Maternal versus environmental constraints on the oocyte size of a marine pelagophil fish. Mar. Biol., 162: 1879-1888. [ Links ]
Guerrero, P. 2011. Descripción histológica en las gónadas de algunas especies pelágicas de la familia Scombridae y Carangidae del departamento del Magdalena, Colombia. Trabajo grado Biol. Mar., Univ. Jorge Tadeo Lozano. Bogotá. 41-53 p. [ Links ]
Guisande-González, C., A. Bareiro, I. Maneiro, I. Riveiro, A. R. Vergara y A. Vaamonde. 2006. Tratamiento de datos. Ediciones Días de Santos. España. 356 p. [ Links ]
Holden, M.J. y D.F. Raitt. 1974. Manual of fisheries science.2. Methods of resource investigation and their application. FAO Fish. Tech. Paper, 115(1): 211 p. [ Links ]
Klibansky, N. y F. Juanes. 2008. Procedures for efficiently producing high-quality fecundity data on small Budget. Fish. Res., 89: 84-89. [ Links ]
Lucano-Ramírez, G., S. Ruiz-Ramírez y J.A. Rojo. 2005. Biología reproductiva de Prionotus ruscarius (Pisces: Triglidae) en las costas de Jalisco y Colima, México. Rev. Dig. Univ. 13 p. [ Links ]
MADR y CCI. 2006. Pesca y acuicultura Colombia 2006. Sistema de información sectorial pesquero. Bogotá. 21 p. [ Links ]
Marteinsdottir, G. y G.A. 2002. Essential relationships incorporating the influence of age, size, and condition on variables required for estimation of reproductive potential in Atlantic cod Gadus morhua. Mar. Ecol. Prog. Ser., 235: 235-256. [ Links ]
Murúa, H., G. Kraus, F. Saborido-Rey, PR. Witthames, A. Thorsen y S. Junquera. 2003. Procedures to estimate fecundity of marine fish species relation to their reproductive strategy. J. Northw. Atl. Fish. Sci., 33: 33-54. [ Links ]
Murua, H., P. Lucio, M. Santurtun y L. Motos. 2006. Seasonal variation in egg production and batch fecundity of European hake Merluccius merluccius (L.) in the Bay of Biscay. J. Fish Biol., 69: 1304-1316. [ Links ]
Palacio, B.C. 2009. Algunos aspectos reproductivos de las principales especies ícticas capturadas artesanalmente entre Tasajera y la bahía de Santa Marta Caribe colombiano. Trabajo grado Biol. Mar. Univ. Jorge Tadeo Lozano, Santa Marta. 118 p. [ Links ]
Pinilla, G.A. 1986. Ecología trófica del jurel (Caranx hippos Linnaeus, 1766) la cojinoa (Caranx crysos Mitchill, 1815) y el ojo gordo (Selar crumenophthalmus Bloch, 1793) (pisces: Carangidae) en aguas costeras del departamento del Magdalena, Caribe colombiano. Tesis Biol., Univ. Nacional de Colombia. Bogotá. 90 p. [ Links ]
Posada, A.M. y C. Sandoval. 2007. Algunos aspectos de la biología reproductiva del jurel, Caranx hippos (Linneo, 1766) (Pisces: Carangidae) en Bocas de Ceniza -departamento del Atlántico Caribe colombiano. Tesis Biol., Univ. Atlántico. Barranquilla. 80 p. [ Links ]
Posada-Peláez, C., A. Santafé-Muñoz, M Grijalba-Bendeck, D. Bustos-Montes, E. González-Sarmiento y J. De La Hoz-Maestre. 2012. Aspectos biológicos de Caranx hippos (jurel aleta amarilla) capturado por la pesca artesanal en el departamento del Magdalena, Caribe colombiano: 191-207. En: Grijalba-Bendeck, M, D. Bustos-Montes, C. Posada-Peláez y A. Santafé-Muñoz (Eds.) 2012. La pesca artesanal marítima del departamento del Magdalena: una visión desde cuatro componentes. Fundación Universidad de Bogotá Jorge Tadeo Lozano, Ministerio de la Agricultura y Desarrollo Rural, Proyecto Transición de la Agricultura, Bogotá. 454 p. [ Links ]
Restrepo, D. 2010. Biología reproductiva de las especies ícticas más importantes para las pesquerías artesanales del área Tasajera - Santa Marta, (Magdalena, Colombia) entre febrero y octubre (2009). Tesis Biol. Mar., Univ. Jorge Tadeo Lozano. Santa Marta. 113 p. [ Links ]
Reyes, A. 1999. Hábitos alimentarios y algunos aspectos reproductivos de la cojinoa negra Caranx crysos (Mitchill, 1815) capturada con chinchorro en la bahía de Taganga, Caribe colombiano. Tesis Biol. Mar. Univ. Jorge Tadeo Lozano. Santa Marta. 131 p. [ Links ]
Saborido-Rey, F. 2004. Ecología de la reproducción y potencial reproductivo en las poblaciones de peces marinos. Universidad de Vigo, Vigo, España. 69 p. [ Links ]
Urriola, M., J. Cabrera y M. Protti. 2004. Fecundidad, fertilidad e índice gonadosomático de Poecilia reticulata (Pisces:Poeciliidae) en Heredia, Costa Rica. Rev. Biol. Trop., 52(4): 945-950. [ Links ]
Wright, P.J. y E.A. Trippel. 2009. Fishery induced demographic changes in the timing of spawning: consequences for reproductive success. Fish, 10: 283-304. [ Links ]
RECIBIDO: 01/07/2015 ACEPTADO: 13/05/2016

