











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
INTRODUCTION
Coastal ecosystems are highly complex and undertake countless processes, besides they are involved in an enormous amount of ecosystem services (Roberts et al., 2003; Harborne et al., 2008; Barbier et al., 2011). For example: they fulfil a crucial role in the recycling of nutrients, play an important role in maintaining water quality, protect shoreline from natural erosive processes, contribute to the global carbon stock via carbon storage, function as spawning and nursery grounds for several fish species, be key habitats in migratory routes, and so on. Moreover, benthic organisms, which inhabit coastal habitats, function as important food sources for higher organisms such as fish. Therefore, these habitats and the connectivity among them are crucial for the successful life cycle of species and to guarantee the maintenance of genetic diversity of gene pool protection (Moberg and Folke, 1999; Liquete et al., 2013; Lee et al., 2014).
Recently, map coastal habitat spatial distribution has been recognized for its enormous importance in management and conservation (Wright and Heyman, 2008). The process of producing seafloor habitat maps cuts across the disciplines of marine biology, ecology, geology, hydrography, oceanography and geophysics, and involves the combining of disparate data sets from these disciplines to produce simplified spatial representations of the seafloor relating to the distribution of biological characteristics (Shumchenia and King, 2010).
Although benthic biology is influenced by both seafloor characteristics (geological and morphological) and the overlying water column influence benthic biology, in order to simplify it can be used substratum type as the determinant parameter (Lund and Wilbur, 2007). Substrate type is one of the most stable and commonly used parameters to classify the seafloor of coastal habitats, and its effect on the spatial distribution of benthic fauna is widely recognized (Cousson and Bourget, 2005; Becking et al., 2006; Howell, 2010; Beaman et al., 2016). In tropical regions, the extent and nature of biogenic communities, such as coral reefs, sea grass beds and mangrove forests, can be mapped as biogeomorphical units (Roff and Zacharias, 2011). Thus, mapping is a vital first step in unravelling ecological complexities and providing improved spatial information for management of marine systems (Brown et al., 2011).
Colombian North Pacific coast is a biologically valuable ecosystem, and the local communities rely heavily on its natural resources (Giraldo and Valencia, 2008). Given its importance, a proactive approach to marine and coastal conservation is vital. Due to this, the aim of this study was to develop a practical habitat (intertidal and subtidal) classification and create a fine-scale map of coastal habitats of two ecologically important sites of the Colombian North Pacific coast: Bahía Aguacate and Bahía Cupica (Figure 1), which was not previously available for the region.
STUDY AREA
Both locality have rather unique combination of marine conditions because: (i) shallow-water habitats are limited due to the narrow continental shelf and steep slope, (ii) it is a remote area there is limited accessibility to the shore, since there is no land access infrastructure to most areas (iii) the rain forest terminates at the water's edge (iv) and as it's sparsely populated the anthropogenic pressure is considerably low. Bahia Aguacate (6° 51' 49" N, 77° 40' 05" W) extend over approximately 17 km of shoreline, standing out Cabo Marzo (6° 50' 08" N, 77° 40' 42" W), where the geomorphology creates a special site named El Acuario (6° 49' 56" N, 77° 40' 45" W) surrounded by islets and crags, which provide exceptionally unique marine conditions. Bahia Cupica (6° 41' 11" N, 77° 27' 42" W) extend over approximately 36 km. The area is characterized by the presence of the Cupica mangrove and an outstanding cove named Piñas (6° 39' 16" N, 77° 30' 43" W) in Punta Cruces (Figure 1).
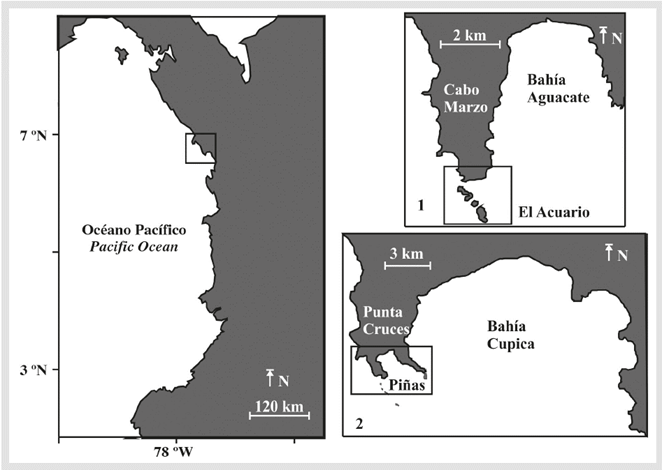
Figure 1 Map of Colombian Pacific coast showing the general study site indicated by a black square. The specific study sites (1) Cabo Marzo and Bahia Aguacate (2) Punta Cruces and Bahia Cupica are shown separately with Piñas and El Acuario indicated by black squares.
During 2008, an ecological characterization of key ecosystems was carried out in both bays, which included coral communities (Zapata et al., 2008a), coral and rocky icthyofauna (Zapata et al, 2008b; Castellanos et al., 2008), mangroves and intertidal rocky shore fauna (Cantera et al., 2008; Londoño-Cruz et al., 2008) and pelagic oceanographic conditions (Giraldo et al., 2008). However, habitat mapping effort was not carried out. In order to create a map, first we must establish habitat classification.
MATERIALS Y METHODS
The goal of habitat classification is to provide a language through which data and information regarding habitats can be communicated and managed. Greater understanding of how habitats relate to each other and the environment around them is essential for ecosystem based management (McDougall et al., 2007). There are a number of elements and issues to take into consideration when classifying marine habitats. It is critical to take into account that habitat classification is scale dependent; however, the scheme should be adaptable to various scales as needed for management.
For this research habitat was defined as both the physical and environmental conditions that support a particular biological community together with community itself (Lund and Wilbur, 2008) and the term is used synonymously with "biotope". This definition makes the assumption that in order to spatially represent biological patterns, we must impose distinct boundaries between adjacent habitat types. Therefore, a standardized hierarchical classification system suitable for the study area was developed. This approach also allows other categories to be added in the future. The classification was made taking into account previous literature concerning habitat types and biodiversity elements of the Colombian coasts classified by Segura-Quintero et al. (2012).
The intertidal habitat classification was mapped according to the categories summarized in Table 1. The mapping rules for level 2, coast type rules applied in this study were: (i) a sandy beach was identified where the entire intertidal was comprised of sand, (ii) a rocky shore was identified where the entire intertidal was comprised of rock, (iii) mixed shores comprised of a combination of sand and rock; (iv) islets are recognized as a feature in their own right and thus are not categorized into their habitat types; (v) a river mouth shore was identified along the river mouth region; (vi) Mangroves are mapped as biogeomorphical units.
Table 1 The hierarchical classification scheme used to code intertidal environments along the study site shoreline.
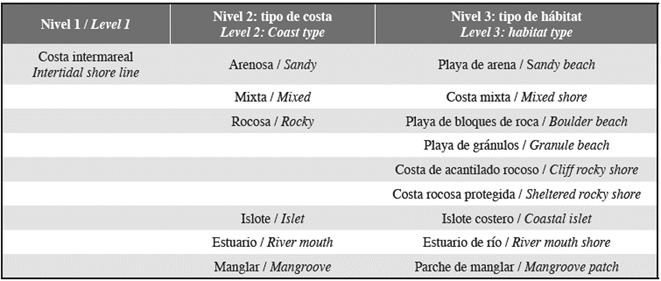
Rocky coast type was further subdivided by habitat types, by the following mapping rules. Cliff rocky shores were recognized when the rock is almost perpendicular to the water. When the rock creates some kind of shelter at the intertidal region, we named it sheltered rocky shore. The region's strong tidal variation, coupled with sheltered rocky shores enable the formation of puddles, which generally have species specially adapted to this habitat type. Boulder beach was identified where beach deposit consists largely of boulders (256-4096 mm) and Granule beach was recognized where beach deposit consists largely of granule (2-4 mm).
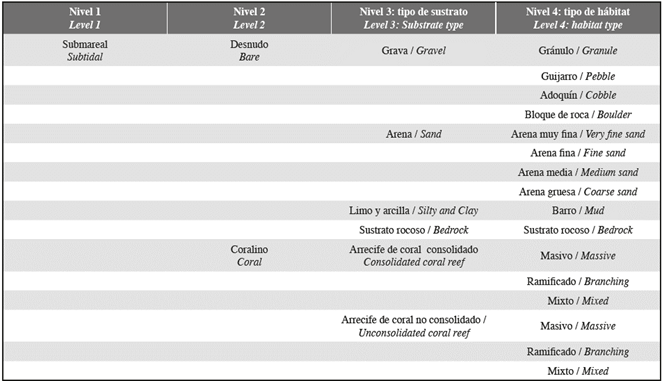
Subtidal habitats were identified along the coast on the continental shelve down to 50 m. The subtidal region was mapped according to Table 2. For subtidal classification, we use the substrate type to define level 2: sediment grain size and biogenic structures (coral). Two main categories are recognized (bare and coral). Bare is define as having a seabed with 0 to less than 10% colonization by corals. Level 3 divisions were based on the seabed's substrate type. The substrate may be comprised of bedrock or unconsolidated sediments-gravel, sand, silt and clay. When the seabed was comprised of unconsolidated sediments, the habitat type was determined by the major grain size: boulder (256-4096 mm), cobble (64-256 mm), pebble (4-64 mm), granule (2-4 mm), coarse sand (0.50-2.00), medium sand (0.25 0.50), fine sand (0.125-0.25 mm), very fine sand (0.0625 0.125 mm), and mud (<0.0625).
Coral Reef is comprised of a hardened substrate formed by reef building corals. Often bedrock is the base for these reefs but the presence of coral or remnant coral on the bedrock surface above 10% is reason to categorize the dominant habitat as coral reef. Substrate consisting of 10% or greater reef hard bottom was considered coral and subdivided by substrate type for level 3 into consolidated coral reef or unconsolidated coral reef. A further level 4 was determined based on habitat type, assisted by coastal and marine ecological classification standards. Level 4 was subdivided based on the dominant specie massive, branching or mixed (Table 2). We identify as massive coral habitat type areas dominated by massive corals that are characteristically ball- or boulder-shaped and relatively slow growing. This type includes reefs dominated by submassive corals, which are similar to massive corals. Branching habitat type was identified at areas where was dominant coral that grow in a tree-like shape and have numerous branches, some with secondary branches. Areas where massive and branching coral abundance were similar were recognized as mixed coral habitat.
Table 2 Hierarchical classification scheme used to code intertidal environments along the study site shoreline.
Fieldwork was challenging due to logistic and resources limitations imposed by the study site. Shoreline subtidal classification was made along transects parallel to the coast geographically located using Global Positioning System (GPS). Habitat types were classified in situ and geo-referenced. In order to make near shore subtidal classification of the shallower areas, both study sites were subjected to underwater field observation. Subtidal survey methodologies used were dependant on water depth. Underwater snorkel, photo and video survey were the methods used for the first meters, close to the shore, where discontinuous boundaries exist between seafloor features. Further offshore, where there are gradational shifts in seafloor characteristics, we use grab as the sampling technique. Subtidal classification of the deeper areas was accomplished by analyzing grain size of grab samples.
Sediment grab samples was collected from discreet point locations of a grid (0.005°). A representative subsample of dry sediment was toked and ran it though a set of US Standard Sieve Mesh and the Phi Scale to classify the sediment by size classes. A histogram and accumulative curve were constructed for all the samples. The median and the mean grain size were computed using R software. For the map, we used the statistical median of the samples, which represents the value of the middle item in a set of data arranged in rank order. Graphic median was calculated with the breakpoint of 50%, and Graphic mean was calculated by the mean grain size of the cumulative 16, 50 and 84 percent.
Coastline and some intertidal features were delineated from Google Earth aerial imagery available for the study zone. ArcGis 10 (Geographic Information System from ESRI) and its extensions were used to map data from an Excel database. The WGS84 (Universal Tranverse Mercator) projection was used for all shape files. Shoreline classification was mapped from GPS points converted into lines. To produce continuous coverage data layers of the segmented grain size classed from the analysis, we used the median grain size data. The points of the grid were transformed to a raster shape.
RESULTS
The intertidal habitat types found at both study sites are very heterogeneous (Fig. 2, Fig. 3). The largest stand of mangrove is found in Bahia Cupica, along Cupica River (Fig. 4). Cupica Mangrove extends behind a sandy beach. The presence of bit quantity of islets and crags along the shore, it is also characteristic at both study sites (Fig. 4), and different types of beaches (Fig 4). Subtidal substrate type of shallow water area of both sites, show a gradual decrease of grain size with depth (Fig. 5). This trait dominates silt and clay areas situated at the inner part of the bays. El Acuario at Cabo Marzo and Piñas at Punta Cruzes exhibit more seabed habitat heterogeneity. Due to the fact that at previous studies they only found significant coral reef growth at Piñas and El Acuario, more detailed maps were made for these locations. Figure 6 shows coral formations spatial distribution. Coral were found forming on mainly unconsolidated coral reef with small colony density (Fig. 7). At El Acuario, we found mainly branching coral types, but also mixed communities of branching and massive coral. Piñas area presented more abundant massive type of coral.
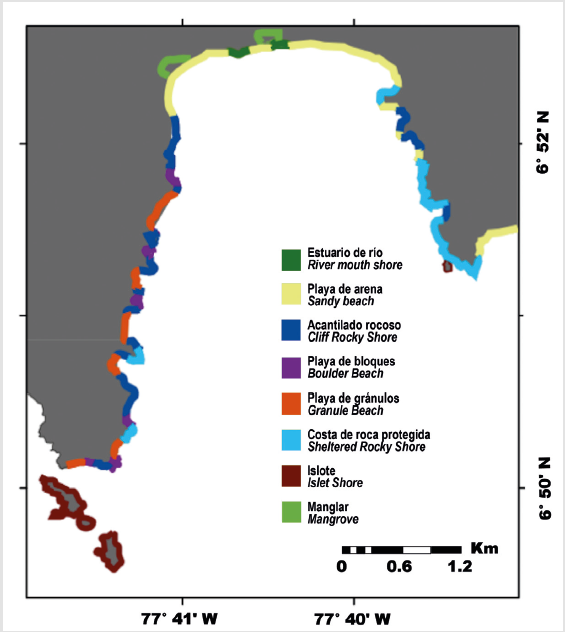
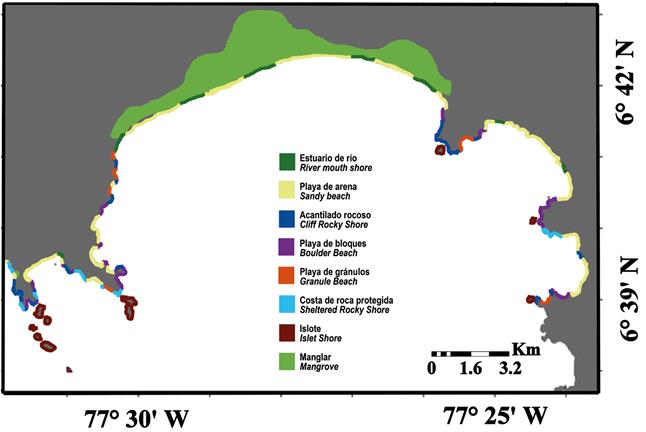

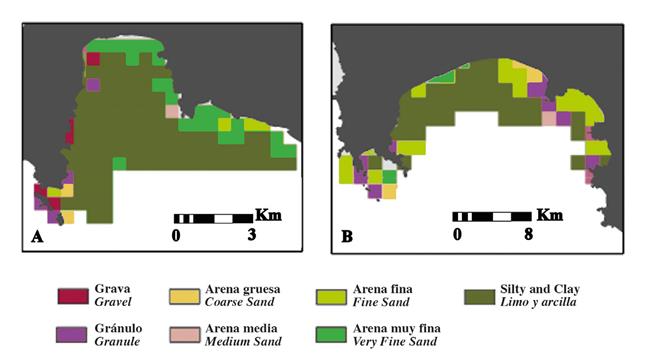
Figure 5 Habitat types of sediment grain size (A) Cabo Marzo and Bahia Aguacate; (B) Punta Cruzes and Bahia Cupica.
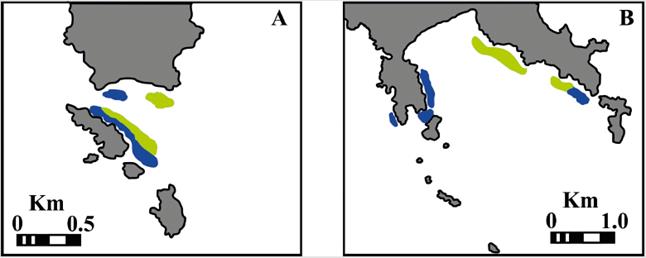
Figure 6 Coral spatial distribution at (A) El Acuario (B) Piñas. Blue: branching coral reef. Green: massive coral reef.

Figure 7 Images of subtidal habitat unconsolidated coral reef type (branching and massive) in El Acuario - Cabo Marzo and Piñas - Punta Cruces.
El Acuario and Pina were identified as two main areas highly heterogenic based on grain size, which simultaneously are the areas where the coral formations where found, stressing the importance of these areas for the coastal ecosystem of the region. Another characteristic emphasized from the subtidal maps is the simultaneous fast decrease of grain size with the steep slope, probably related with ocean currents patterns of the region. The inshore-offshore pattern clearly reflected the hydrodynamic conditions of the area, from highly hydrodynamic shallow depth to the calmer deeper areas. Based on the map we can identify two sedimentary areas: the inshore are characterized by coarser sands and the offshore area that includes finer sediments. At the inner part of both bays (Bahia Aguacate, Bahia Cupica) there was an area of predominant silt/clay sediment. This finer sediment is mainly due to the proximity to the river mouths that supplies this region with large amounts of fine particles.
DISCUSSION
The majority of today's technologies and methods used for ocean mapping involve indirect or remote sampling (satellite data, multibeam and single-beam sonar, sides can sonar, LiDAR) (Mumby et al., 1999; Mishra et al, 2006; Brown et al., 2011; Micallef et al, 2012). These methods are extremely expensive and require advanced technology, which is not always available. Moreover, they have limitations due to environmental factors such as water turbidity (Van Rein HB et al, 2009). In spite of the fact that the selection of in situ sampling techniques, for instance snorkel/SCUBA dive observations or grab samples, require extensive sampling effort in order to create an adequate fidelity map, we chose this approach because they are accurate and easily implemented at remote locations.
The habitat classification used in this research was based in biogeomorphic units and substrate type because these are conservative parameters which influence both the benthic community (e.g. macrofauna, meiofauna) and the pelagic community living above (e.g. fish assemblages) (Bartley et al., 2001; Huang et al., 2011). Moreover, biogemorphic units -mangroves, at intertidal level; and coral formations, at subtidal level-represent key marine coastal habitats hosting biodiversity offering a wide range of ecosystems services (Friess and Webb, 2014; Yee et al, 2015)
At the intertidal level results of this study illustrate a diverse and heterogeneous shoreline, which may be attributed to the unique characteristics of the local geology, climate and oceanography. The geology is the first order determinant of the nature and distribution of coastline ecosystems and habitats (e.g. sheltered rocky shores) at broad and fine scales, respectively (Harris et al, 2013). Regional climate and oceanography are also major influences. Local tropical climate is characterized by abundant rainfall along the coast. Freshwater-input-associated systems are highly influenced by rainfall patterns and rivers flood sporadically (Poveda, 2004; Rangel and Arellano, 2004). This has implications for the distribution of coastal habitats, like mangroves, that are associated with fresh water systems. In addition, abundant rainfall also increases weathering and erosion, resulting in an increase of terrigenous sediment to the coast and increasing area of sandy beaches (McLachlan and Brown, 2006; Short, 2010).
Other climatic factors that influence shoreline and subtidal features include temperature and wind, which affect the prevalence and morphology of different coastal habitats (Harris et al, 2013). The annual variation of the intertropical convergence zone (ITCZ) has a direct effect on the climate in the region influencing all local oceanographic processes (Giraldo and Ramírez, 2010; Poveda et al, 2011; Hoyos et al, 2013; Villegas et al, 2016), therefore, the biology of coastal habitats. In summary, the nature and heterogeneity of coastal habitats and features along the north Colombian Pacific coast is largely the result of interacting patterns in the underlying geology, climate (rainfall, wind regime, temperature and humidity) and oceanography (sea surface temperature, currents, and wave climate). The shoreline studied show a highly variable topography that leads to a complex diversity of coastal habitats, from beaches to cliffs. The maps show bigger presence of cliff rocky shore at shoreline oriented to west, boulder beaches oriented to the east and most sandy beaches oriented to the south. In addition, the region exhibits high abundance of islets and crags; these features might modify the nearshore hydrography patterns of the region, promoting the concentration of early stages of the hydrobiological resources of the region.
By mapping the spatial distribution of coastal habitats, we expect to facilitate quantification of biologically relevant patterns on the shoreline and seafloor. This will help to address important ecological questions pertaining to animal-environment interactions at further studies, and facilitate the local community's ability to manage coastal ecosystems effectively. Proper management needs also to integrate sociological, cultural and economic aspects of the region. Therefore, there is need to increase sustainable and eco-friendly economical activities as a way of money income. The present study contributed towards filing a knowledge gap, by offering a map with the spatial distribution of key habitat types.

