











 text in
text in  Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los arrecifes coralinos son bio-construcciones (i.e. estructuras construidas por organismos) que aportan gran variedad de micro-habitas a diferentes especies de peces e invertebrados marinos (Cumming, 1999; Alvarez-Filip y Gil, 2006), por lo cual son considerados como uno de los ecosistemas con mayor diversidad biológica en todo el mundo (Rotjan y Lewis, 2008), una característica que le confiere a los ecosistemas alta resiliencia (Valiente-Banuet et al., 2015). A pesar de ello, estos ecosistemas son vulnerables a amenazas de tipo biótico, e.g. bioerosión causada por diversas especies de peces, anélidos, crustáceos, equinodermos y moluscos (Glynn y Enochs, 2008) como a factores de tipo abiótico, e.g. cambios en la temperatura (que puede causar blanqueamiento), mareas bajas extremas (que generan mortalidad de corales y otros organismos asociados) y tormentas (que causan daño físico al andamiaje arrecifal). En adición, se pueden ver afectados por actividades humanas como pesca excesiva, contaminación de las aguas y extracción manual del coral, entre otros (White et al., 2000; Huges et al., 2003), actividades que pueden potenciar los efectos naturales. En los últimos años, se ha dado gran importancia al papel que cumplen los factores bióticos de estrés sobre los corales, ya que afectan directa y negativamente la aptitud física (fitness) del coral (Rotjan y Lewis, 2008), afectando todo el ecosistema. Uno de estos factores bióticos es la coralivoría; proceso que consiste en el consumo directo de los pólipos coralinos por parte de depredadores, ocasionando la muerte total o parcial de la colonia coralina o restringiendo su capacidad de recuperación por del daño infligido (Baums et al., 2003).
Existen diversos grupos animales que se han especializado como coralívoros, dentro de los cuales los moluscos tienen una importante representación. En la actualidad se conocen 20 especies coralívoras de este grupo, todas pertenecientes a la clase Gasteropoda (Rotjan y Lewis, 2008) dentro de los que se encuentra Jenneria pustulata, (Lightfoot, 1786), un caracol perteneciente a la Familia Ovulidae que se puede encontrar desde el norte del Golfo de California hasta el Perú y que se alimenta de corales escleractineos (Lorenz y Feshe 2009).
En Colombia, esta especie se ha registrado en los arrecifes coralinos de Isla Gorgona y la ensenada de Utría (Navas-Camacho et al., 2010); sin embargo, no se han realizado investigaciones conducentes a conocer sus efectos sobre estos arrecifes coralinos. De esta forma, la presente investigación pretende determinar la distribución de tallas y cuantificar la tasa de coralivoría de este depredador en los arrecifes de La Azufrada y Playa Blanca del Parque Nacional Natural Gorgona.
ÁREA DE ESTUDIO
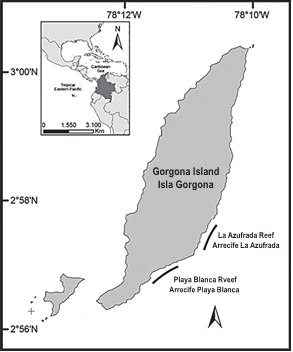
La isla Gorgona (2°58' N, 78°11' W; Figura 1), ubicada aproximadamente a 30 Km del continente (Guapi, Cauca), es el territorio insular más extenso (13,2 Km2) sobre la plataforma continental del Pacífico colombiano. La isla está influenciada por el Cinturón de Convergencia Intertropical, lo que determina su clima bi-estacional (Zapata et al., 2010). La temperatura superficial del mar varía entre 26º y 29ºC, pero en ocasiones puede descender a menos de 19ºC durante eventos de surgencia hacia comienzos del año (Díaz et al., 2001; Zapata, 2001). Las principales formaciones coralinas (La Azufrada y Playa Blanca) se encuentran sobre el margen oriental de la Isla. La Azufrada (LA) mide ~780 m de largo por ~80-180 m de ancho y cubre cerca de 0.094 km2. Playa Blanca (PB) está constituida por dos grandes parches arrecifales separados por un canal de ~60 m de ancho: el más pequeño, hacia el norte, tiene ~240 m de largo por ~40 m de ancho, mientras que el más grande, hacia el sur, tiene una longitud ~930 m y un ancho que varía entre ~60 m y 230m. En conjunto cubren un área aproximada de 0.098 km2. El principal constructor del andamio arrecifal es la especie de coral ramificado Pocillopora damicornis (familia Pocilloporidae) (Parada y Nikolaevna, 1990; Zapata, 2001; Zapata y Vargas- Ángel, 2003; Palacios et al., 2014)
MATERIALES Y MÉTODOS
Trabajo de campo y laboratorio
Se efectuaron dos salidas de campo: una al arrecife LA (abril de 2015) y otra al arrecife de PB (agosto de 2015). Como estos caracoles viven bajo las colonias de coral (Figura 2), se revisaron colonias sueltas de la matriz arrecifal, utilizando equipos SCUBA, en dos sesiones al día. Se colectaron y midieron (longitud de la concha en mm) todos los individuos de J. pustulata encontrados bajo las colonias; posterior a la medición fueron liberados. Con los datos se determinó la estructura de tallas de la especie en cada arrecife. En adición, en cada arrecife se colectaron 15 individuos (y un número idéntico de fragmentos de coral) con los cuales se realizó el experimento de consumo coralino bajo condiciones controladas en los laboratorios del PNN Gorgona.

Figura 2 Jenneria pustulata alimentándose de una colonia coralina. Fotografía por: Juan Felipe Lázarus.
Los individuos de J. pustulata y los fragmentos de coral previamente colectados durante la fase de campo, fueron utilizados para realizar el experimento de coralivoría.
En acuarios de 16 L de capacidad (n =15) se introdujeron de manera individual por un periodo de 24 h, individuos de J. pustulata y fragmentos vivos de coral (un individuo de J. pustulata y un fragmento de coral por acuario; el fragmento de coral asignado a cada caracol provenía de la colonia en la cual el caracol fue capturado). Durante el desarrollo de los experimentos, los acuarios fueron vigilados y el agua fue recambiada y oxigenada (con motores de ventilación) de manera constante. Posteriormente, las lesiones resultado de la depredación por parte del caracol (Figura 3), fueron fotografiadas utilizando una cámara Canon D30. Con el software de libre distribución ImageJ 64 (Abramoff et al., 2004) se calcularon las áreas (mm2) de las lesiones a partir de las fotografías.

Análisis estadístico
Después de confirmar los supuestos de las pruebas paramétricas, se evaluaron las tallas de los organismos mediante análisis de varianza simple con arrecife (2 niveles fijo) como factor explicativo. Además, se comparó la estructura de tallas de los organismos provenientes de cada arrecife con una prueba de Anderson Darling para dos muestras (Engmann et al., 2011). En adición, se realizó un análisis de varianza simple sobre los datos de consumo de tejido coralino obtenidos en cada uno de los arrecifes. Finalmente, el consumo de tejido coralino se evaluó con un análisis de covarianza, utilizando el arrecife de procedencia como variable explicativa y la talla de los caracoles como la covariable. Todos los análisis estadísticos fueron realizados utilizando el programa de libre distribución R (R Development Core Team, 2014) y siguiendo los supuestos estadísticos de Zar (Zar, 1999).
RESULTADOS
Talla y estructura de tallas
Se examinaron y midieron un total de 201 individuos de J. pustulata, de los cuales 74 (36.8%) fueron colectados en LA y 127 (63.2%) en PB. La media (±DE) del largo de la concha fue ligeramente mayor en PB (20.5±3.40 mm) que en LA (19.6±4.71 mm) (Figura 4). Aunque los anteriores valores son aparentemente similares, el análisis estadístico mostró que dicha diferencia es significativa (F (0,05(2)1;199) =11.24; P < 0.001). En adición, se observa que el rango de variación en los individuos de PB estuvo entre 16.5 y 25.0 mm, mientras que en LA fue más amplio y osciló entre 14.0 y 24.0 mm.
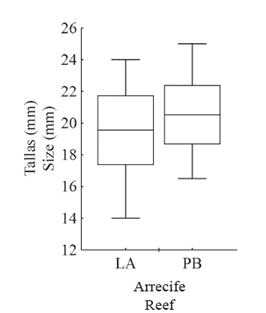
Figura 4 Talla media (Caja: DE, Bigotes: mínimo y máximo) de Jenneria pustulata en los arrecifes La Azufrada y Playa Blanca.
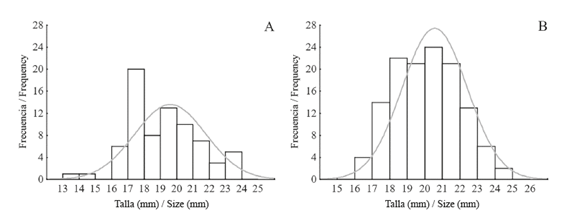
La mayoría (60.8%) de los individuos de J. pustulata en LA se encuentran distribuidos entre 17 y 21 mm de largo. La distribución de tallas de estos caracoles no se ajustó a la distribución Normal (A=0.92; P=0.019) (Figura 5A). Por su parte, la mayoría (78.4%) de los caracoles de PB se encuentran distribuidos principalmente entre 18 y 22 mm de largo; esta distribución de tallas también difirió significativamente de la distribución Normal (A=1.44; P<0.001) (Figura 5B). Finalmente, al comparar la estructura de tallas entre ambos arrecifes, se encontró que son diferentes entre sí (A=5.48; P=0.002).
Consumo de tejido coralino
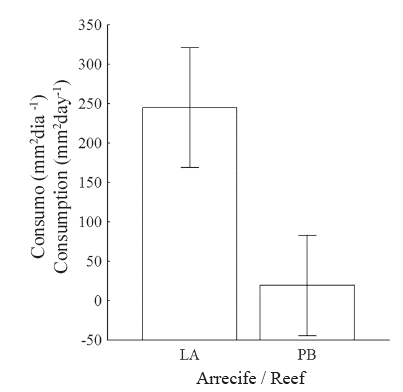
El experimento de coralivoría con los caracoles de LA tuvo un porcentaje de éxito del 44.4%, es decir, de los 18 caracoles iniciales, solo 8 consumieron tejido; mientras que de los caracoles provenientes de PB el 66.7% (10 de 15) consumió tejido coralino. Es claro que existe una relación directamente proporcional entre el tamaño de los caracoles y la cantidad de tejido coralino consumido tanto en LA (R2=0.81) como en PB (R2=0.53) (Figura 6). El consumo medio por caracol ±DE de tejido coralino, en mm2día-1, fue de 105.9±77.01 en LA y 130.5±105.50 en PB. El rango de consumo osciló entre (31.1 - 242.2) y (21.0 - 298.0) mm2día-1 en LA y PB respectivamente.
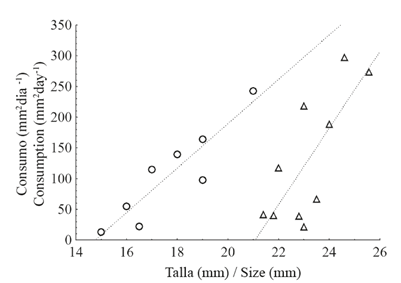
Figura 6 Relación entre el tamaño (longitud de la concha, en mm) y el tejido coralino consumido (mm2) por día por Jenneria pustulata en los arrecifes La Azufrada (P=0.002; círculos) y Playa Blanca (P=0.01; triángulos).
Aunque el consumo medio de tejido coralino, sin considerar la talla de los caracoles es superior en PB que en LA (130.5 vs. 105.9), el ANCOVA mostró que hay un efecto muy significativo del tamaño de los caracoles (P), lo cual muestra que un caracol de un tamaño dado, consumirá más tejido coralino en LA que en PB (Figuras 6 y 7).
DISCUSIÓN
Los arrecifes coralinos ubicados en el costado oriental de la isla Gorgona presentan un alto grado de similitud, tanto en tamaño como en estructura y en su marcada dominancia por corales del género Pocillopora (Prahl, 1986; Palacios et al., 2014). En adición, son espacialmente cercanos, por lo que se podría inferir que existe un alto grado de comunicación. Sin embargo, en el presente estudio se encontraron 1.7 veces más individuos de J. pustulata en PB que en LA, aunque el esfuerzo de muestreo (ca. 24 h en cada arrecife) fue similar en los dos arrecifes. Si bien, como se mencionó, los dos arrecifes son muy similares, la cantidad de sedimento que reciben por escorrentía varía significativamente entre los dos, siendo mayor en PB (293 g/m2 día-1) que en LA (95 g/m2 día-1) (Blanco, 2009). La cantidad de sedimentos que entran al arrecife tienen un efecto importante en la salud del mismo y podrían, eventualmente, hacerlo más vulnerable a enfermedades y reducir la capacidad del coral para competir por el espacio con otros organismos (e.g algas) (Granja y López, 2008; Erftenmeijer et al., 2012). Observaciones en campo, sugieren que J. pustulata tiene una marcada preferencia por colonias con algún grado de deterioro; es posible que esta preferencia se relacione con la capacidad del caracol de mimetizarse con este tipo de colonias, de tal forma que evite, de alguna manera, la depredación por peces como Arothron meleagris, Sufflamen verres y Pseudobalistes naufragium (Guzman 1988; Glynn, 2004). Por otro lado, el mayor número de individuos de J. pustulata encontrado en el arrecife de PB muestran una mayor tasa de depredación por parte del caracol en este arrecife al compararlo con el arrecife de LA; por lo tanto es factible plausible pensar que la sinergia de los efectos resultantes del influjo de sedimentos proveniente de las quebradas al interior de la isla, el consumo de tejido coralino por parte de J. pustulata y la subsecuente colonización del sustrato libre resultado de esta actividad por parte de las algas podrían afectar de forma significativa el estado de salud del arrecife de PB. Sin embargo, para corroborar estas afirmaciones, se necesitan más investigaciones que ahonden en el estado fisiológico de los corales en el arrecife de PB, pues según Tiliyanov y Tiliyanova (2005), la capacidad de recuperarse de una lesión en el tejido vivo depende de varios factores, entre ellos el tipo de lesión sufrida, el tipo de crecimiento (ramificada, masiva o sub-masiva) y el estado fisiológico en el que se encuentre la colonia.
Talla y estructura de tallas
La talla de los gasterópodos es una característica que puede verse afectada por factores internos como el estado de desarrollo del animal y la edad; y por factores externos como las condiciones ambientales (Wilbur y Owen, 1964). Según Bashevkin y Pechenik (2015) la temperatura tiene un efecto importante sobre el crecimiento de los moluscos, con temperaturas más altas ocasionando tasas de crecimiento más aceleradas que temperaturas bajas. Vale la pena mencionar que los arrecifes fueron visitados durante dos épocas diferentes: LA se visitó durante abril de 2015, mientras que PB se visitó durante agosto del mismo año. Las dos épocas presentan condiciones climáticas contrastantes, entre enero y abril hay enfriamiento de las aguas debido a un proceso de surgencia en la parte central del Golfo de Panamá que genera una lengua fría proveniente del norte con influencia sobre las aguas de Pacífico Tropical Oriental colombiano, mientras que durante mayo y diciembre, las temperaturas del agua son superiores (Giraldo et al., 2008). Es posible que esto haya ocasionado en parte las diferencias observadas en términos de largo de la concha (caracoles más grandes en promedio en agosto en PB [aguas cálidas] y más pequeños en abril en LA [aguas frías]) entre los caracoles en los dos arrecifes; sin embargo, es muy deseable probar esta hipótesis haciendo muestreos simultáneos. Por otro lado, existe una relación directamente proporcional entre el tamaño muestral y la densidad poblacional (Connor et al., 2000), lo cual también podría entenderse como que muestras con mayores abundancias podrían capturar un ámbito más amplio de características poblacionales. En PB se capturaron más individuos que en LA por lo cual se esperaría que se obtuvieran individuos en un rango de tamaños más amplio, lo que se vería reflejado en tamaños promedios mayores en dicho arrecife.
Finalmente, en LA existe evidencia de liberación de gases sulfurosos, aunque es una probabilidad muy baja, estos gases podrían estar disminuyendo el pH en las aguas circundantes, con el concomitante efecto sobre calcificación en las conchas y por lo tanto el registro de tamaños promedios inferiores.
Consumo de tejido coralino
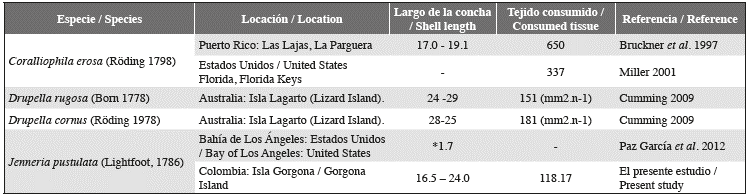
El papel ecológico de los coralívoros ha sido catalogado por algunos autores como el de modelar y estructurar las poblaciones y las dinámicas de la presa; este papel puede verse afectado por factores como la abundancia relativa de la presa y la talla y densidad del depredador en el arrecife (Bruckner et al., 1997). Jenneria pustulata es un depredador con un alto grado de especificidad, que se alimenta casi siempre de corales escleractinios con una marcada preferencia por el género Pocillopora (Bertsch, 1984; Glynn, 2004; Figura 2). Considerando que los dos arrecifes son relativamente similares tanto en composición de especies como en extensión total, se esperaría que los caracoles provenientes de estos, no presentaran variaciones significativas en la cantidad de coral consumido en los experimentos. Sin embargo, según los resultados del ANCOVA, caracoles comparables en tamaño consumirían en promedio más tejido coralino en LA que en PB; como los caracoles en LA son en promedio más pequeños, es posible que estos estén cumpliendo la misma función, en términos de consumo coralino, que caracoles grandes en PB. Aunque esta es una posible explicación a lo observado, es necesario evaluar la existencia de competencia por interferencia en esta especie, lo cual podría ayudar a establecer el hecho de que caracoles en promedio más pequeños en LA tengan el mismo consumo que caracoles más grandes en PB. Por otro lado, estudios realizados sobre otros coralívoros como Coralliophila erosa, Drupella cornus y D. rugosa (Bruckner et al., 1997; Miller, 2001; Cumming 2009) arrojan tasas más altas que las encontradas para J. pustulata en los arrecifes de Gorgona (Tabla 1). Esto podría indicar que aunque esta especie, en promedio es más grande que C. erosa (Del Mónaco et al., 2008) y de tamaño similar a las especies del género Drupela, causa un efecto sobre el ecosistema coralino que podría considerarse poco significativo con relación a las especies anteriormente nombradas. En adición, es importante recalcar que la rádula de J. pustulata presenta dos pares de dientes laterales con una base rectangular y cúspides fuertes y rígidas en forma de gancho (Simone, 2004). Es posible que esta disposición le permita alimentarse sin ocasionar daños permanentes a la coralita, permitiendo que la recuperación por parte del coral sea más rápida y energéticamente menos costosa.
Tabla 1 Tamaño de la concha (mm) y tejido coralino consumido (mm2. día-1) por diferentes especies de moluscos. (mm2.n-1): Registrado durante 12 h. * Medida en cm.
Estudios enfocados en la capacidad que tienen los corales ramificados del género Pocillopora para recuperarse de una lesión a nivel tisular (en donde no se daña el esqueleto calcáreo) demuestran que, aunque este proceso es energéticamente menos costoso, estas heridas tienden a ser colonizados por algas 4 o 5 días después de la aparición de la lesión en el tejido, y, aunque después de un tiempo el coral tiende a recuperarse casi completamente de la herida, esta recuperación implica un gasto energético importante, pues el coral no solo debe generar nuevos pólipos para cubrir de nuevo la zona lesionada sino que también debe regenerar los pólipos que han sido parcialmente dañados por el coralívoro (Titliyanov et al. 2005; Traylor-Knowles, 2016). Esto podría significar que, aunque el daño causado por la alimentación de J. pustulata es menor que el de otros moluscos coralívoros, esta especie puede estar afectando fuertemente la condición del coral debido a la importante inversión energética que ellos deben hacer, no solo para recuperarse del daño recibido sino también para sobrellevar la concomitante colonización del esqueleto calcáreo por parte de las algas. Es debido a esto que es necesario realizar más investigaciones con el propósito de ampliar el conocimiento sobre la estructura y dinámica poblacional de este caracol y corroborar la significancia de sus efectos sobre los arrecifes coralinos del PNN Gorgona.

