











 texto en
texto en  Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
INTRODUCCIÓN
Gonostomatidae es una familia de peces marinos abisales, conocidos comúnmente como peces luminosos; una familia relativamente pequeña que contiene únicamente ocho géneros y 32 especies. Sin embargo, compensan su baja diversidad genérica con su abundancia: Cyclothone, con 12 especies, es probablemente junto con Vinciguerria, el género de vertebrados más abundante del planeta. Se encuentran ampliamente distribuidos en todos los mares cálidos y templados del mundo, Atlántico, Índico y Pacífico (Nelson, 2006). Once especies de gonostomátidos existen en el área de CalCOFI (California Cooperative Oceanic Fisheries Investigations), pero sólo cuatro (Cyclothone acclinidens, C. signata, Diplophos taenia y Gonostoma atlanticum), se encuentran con cierta regularidad en las muestras de ictioplancton de dicha área. Otras cuatro (Cyclothone pseudopallida, Diplophos proximus, Gonostoma ebelingi y G. elongatum), aparecen principalmente fuera de la costa al sur de las estaciones CalCOFI; sus distribuciones al parecer son en gran parte fuera del área CalCOFI (Watson, 1966). Aunque en ocasiones Diplophos proximus Parr (1931) aparece como sinonimia de D. taenia, ambas especies son consideradas válidas según Ozawa et al. (1990).
Los gonostomátidos adultos son de tamaño pequeño (la mayoría < 7 cm, máx. 36 cm, normalmente < 20 cm), poseen ojos sésiles redondos u ovoides sin tejido coroides, dentición pequeña y finos radios branquiostegales. La bioluminiscencia es debida al desarrollo de fotóforos en tallas mayores de 7.5 mm. Intestino de longitud variable, que termina en el origen de la aleta anal. Coloración generalmente negruzca, algunos son plateados. Son ovíparos; las larvas son alargadas y tienen notorios cambios durante el desarrollo, inicialmente ojos ovales, cuerpo poco pigmentado sin fotóforos y finalmente ojos redondeados, pigmentos dorsales y en transformación presentan fotóforos. Los adultos habitan principalmente profundidades medias entre los 300 y 800 m y algunas especies suben de noche a 50 m de la superficie. Se encuentran en las zonas epi (0- 200 m), meso (200-1000 m) y batipelágicas (>1000 m). Morfológicamente se parecen mucho a los phosíchthydos; el estado taxonómico de estas dos familias aún no ha sido resuelto del todo (Watson, 1996).
Según Ozawa et al. (1990), D. proximus, presenta diámetro orbital grande (en proporción al tamaño de la cabeza 21-28%), comparado con las otras especies D. taenia, D. orientalis y D. australis (15-23%). Sus características diagnósticas son: 83-86 vértebras totales (TV), 33-35 abdominales (AV)+ 49-52 vértebras caudales (CV) y 92- 100 fotóforos, suma de todas las series ventrales (IC).
El desove no se ha descrito para D. proximus, pero se ha documentado para otros gonostomátidos, los cuales son ovíparos, con huevos y larvas planctónicas. Las larvas de Diplophos presentan ojos no pigmentados, saco vitelino alargado y boca no funcional, el desarrollo es muy similar a otros gonostomátidos (Ahlstrom et al., 1984; Ozawa y Oda 1986).
Pocas larvas y pequeños juveniles pelágicos se han obtenido en recolecciones realizadas por CalCOFI desde el golfo de California y cerca del Cabo San Lucas. Sin embargo, son mucho más comunes al sur, frente a México y América Central. Para Colombia no se tienen registros de la captura ni de larvas ni de adultos de esta especie. Aparece citada en el artículo de Castellanos-Galindo et al. (2006) como una especie presente en aguas adyacentes al Pacífico colombiano.
Para esta región del Pacífico de Colombia se pueden encontrar las siguientes especies de gonostomátidos: Cyclothone acclinidens, C. microdon, C. signatha, Gonostoma sp. y Yarrella argenteola.Beltrán-León y Ríos (2000) realizaron los primeros registros en estadios larvales de Cyclothone acclinidens y C. signatha, citándolas para las subáreas III y IV y al oriente de la isla Malpelo, con abundancias bajas durante los cruceros de abril de 1993, junio-julio y diciembre de 1995 y noviembre de 1998. Por lo anterior, el presente trabajo contribuye con información de distribución y notas ecológicas de larvas de D. proximus, convirtiéndose en los primeros registros de capturas para el Pacífico colombiano, incluida la descripción de los estadios larvales encontrados.
ÁREA DE ESTUDIO
El Pacífico colombiano incluye las aguas comprendidas latitudinalmente entre las fronteras marítimas con Costa Rica, Panamá y Ecuador y longitudinalmente desde la línea de costa hasta el límite exterior de la zona económica exclusiva que genera la isla de Malpelo (740 km a partir de la costa continental), con 339500 km2 entre aguas estuarinas, costeras y oceánicas con profundidades hasta de 4000 m (Beltrán-León y Ríos, 2000).
Ubicado en el límite occidental del Pacífico Oriental, se caracteriza por la leve variación climática que se genera por su ubicación respecto al sol. La fisicoquímica de las aguas depende en buena parte de los fenómenos locales como el movimiento de la Zona de Convergencia Intertropical (ZCIT), los drenajes dulceacuícolas continentales y el afloramiento en Panamá, como también de un complejo patrón de corrientes superficiales. Las principales corrientes que afectan las aguas del Pacífico colombiano son la ciclónica de Panamá, la de Colombia y la contracorriente Ecuatorial.
En términos generales se presenta un contraste entre las aguas oceánicas y las costeras, incluidas las estuarinas, especialmente hacia el sur de Cabo Corrientes. Las primeras presentan en la superficie temperaturas medias entre 25 y 27°C y salinidades medias de 33.5. Los valores extremos máximos de temperatura (hasta 31°C) se han detectado con ocasión del fenómeno del Niño; mientras mínimos de hasta 18°C y salinidades cercanas a 35 se han registrado en los primeros meses del año y se deben al afloramiento del golfo de Panamá (Prahl et al., 1990). Para los estudios del Instituto Nacional de Pesca y Acuicultura -INPA, la zona fue estratificada por Zapata et al. (1998), en cuatro subáreas seleccionadas latitudinalmente de norte a sur según el tipo de costa, ancho de la plataforma, aporte de ríos y valores de temperatura y salinidad principalmente ((Figura 1).
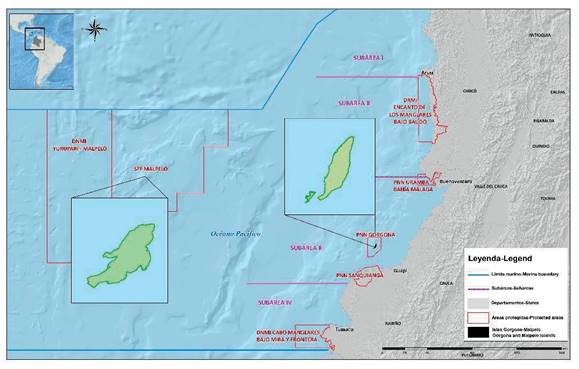
Subárea I: entre la frontera con Panamá y Cabo Corrientes (07°13' Lat. N - 05°30' Lat. N). Altamente influenciada por la surgencia de Panamá, zona de alta pluviosidad, con poca influencia de ríos, plataforma estrecha más o menos de 1.8 a 3.7 km y sustrato rocoso en el fondo.
Subárea II: entre Cabo Corrientes y bahía Málaga (05°30' Lat. N - 04°00' Lat. N). La plataforma, más ancha que la anterior, variando entre 9 y 19 km, recibe mayor influencia de ríos que en la subárea I, especialmente debido al gran aporte de agua dulce del río San Juan y posee generalmente sustrato areno fangoso en el fondo.
Subárea III: entre bahía Málaga y Pasacaballos (04°00' Lat. N - 02°30' Lat. N). Este sector tiene mayor aporte de agua dulce que las dos anteriores por la gran cantidad de ríos, su plataforma es bastante extensa llegando a 56 o 74 km y con sustrato areno fangoso.
Esta subárea se caracteriza por ser una zona de convergencia de flujos costeros, bien sea de origen oceánico, de mareas o fluviales. Por su amplia plataforma y la ubicación geográfica, las mezclas de agua son más evidentes y duraderas que en las otras subáreas del litoral.
 En general, la disposición superficial de los parámetrosvoceanográficos describe la época de final de año como de enfriamiento estacional por las temperaturas superficiales más bajas (
En general, la disposición superficial de los parámetrosvoceanográficos describe la época de final de año como de enfriamiento estacional por las temperaturas superficiales más bajas ( =26°C) (noviembre y diciembre); contrarias a las mayores temperaturas detectadas a mediados de año ( =28°C) (mayo y julio). La salinidad superficial es menor durante las épocas húmedas o de invierno ( =20-29) (mayo y noviembre) y el oxígeno es la variable más homogénea, presentando el valor promedio más alto durante julio (4 ml/l).
=26°C) (noviembre y diciembre); contrarias a las mayores temperaturas detectadas a mediados de año ( =28°C) (mayo y julio). La salinidad superficial es menor durante las épocas húmedas o de invierno ( =20-29) (mayo y noviembre) y el oxígeno es la variable más homogénea, presentando el valor promedio más alto durante julio (4 ml/l).
Subárea IV: localizada entre Pasacaballos y frontera con Ecuador (02°30' Lat. N - 01°30' Lat. N). Su gran aporte de agua dulce proviene de los ríos Patía y Mira, su plataforma es muy angosta, variando entre 9 y 19 km y tiene sustrato areno fangoso.
Se halla más influenciada por aguas oceánicas y ecuatoriales (propias de la contracorriente ecuatorial). Por este motivo la caracterización oceanográfica consiste en presentar aguas más frías (27.4-27.6°C) e hialinas (31-34) que las ubicadas al norte bajo una mayor influencia costera y pluviométrica. Esta situación es independiente de la época anual, y es la intensidad de los vientos alisios, lo que permite que dicha contracorriente tenga más o menos alcance en la costa colombiana.
Según Zapata et al. (1998), la importancia del Pacífico colombiano está representada por ser el punto de confluencia de variados elementos de la fauna submarina de interés zoogeográfico pertenecientes a diferentes provincias como la Indo-Pacífica, Caribe, Golfo de California y Peruano- Chilena, además de los de distribución circumtropical o cosmopolitas. Estas aguas reflejan en su composición faunística no solo la dinámica actual sino también la de un remoto pasado geológico, aún poco conocido.
MATERIALES Y MÉTODOS
El presente trabajo se realizó con 24 muestras positivas para larvas de la especie, obtenidas durante nueve cruceros: dos cruceros ERFEN (Estudio Regional Fenómeno de “El Niño”), los cuales se realizaron en toda el área del Pacífico colombiano durante los septiembres de 2003 y 2005 y otros siete cruceros realizados en la línea recta entre Buenaventura y el Santuario de Fauna y Flora Malpelo durante julio y diciembre de 2010, diciembre de 2013, febrero-marzo de 2014, junio de 2016, junio y septiembre de 2017, incluidas estaciones alrededor de la isla. El muestreo de ictioplancton y oceanografía se realizó a lo largo de 10 transectos perpendiculares a la costa, paralelos entre sí y separados 54 km, con 42 estaciones para los cruceros ERFEN de 2003 y 2005 (Figura 2) y 20 estaciones, ocho en línea recta entre Buenaventura y Malpelo y seis estaciones diurnas y nocturnas alrededor de la isla durante 2010 y 2017 (Figura 3).
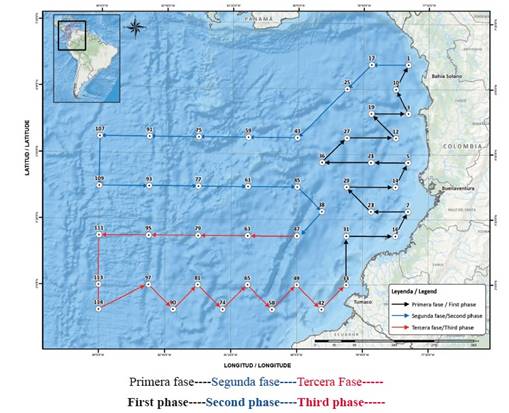
Figura 2 Estaciones y ruta de muestreo Cuenca del Pacífico Cruceros ERFEN 2003 y 2005 (Tomado de CCCP 2004-2007 Planes de ejecución Cruceros ERFEN).
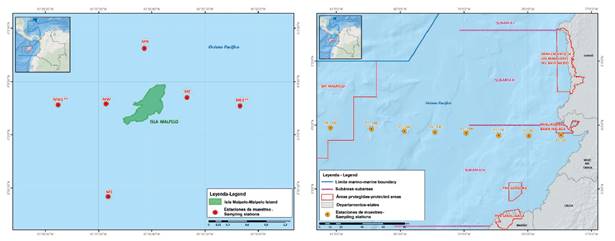
Figura 3 Estaciones de muestreo en el Santuario de Fauna y Flora Malpelo y el trayecto costa-Malpelo
Las muestras se obtuvieron mediante el empleo de una red bongo estándar de 60 cm de diámetro en cada una de las bocas, provista de mallas de 300 y 500 micrómetros, copos colectores blandos y contadores de flujo análogos Hydrobios (Figura 4), efectuando arrastres oblicuos de acuerdo con la metodología establecida por Smith y Richardson (1979), hasta una profundidad máxima de 200 m. Las muestras se fijaron con formol neutralizado con bórax, al 10% en agua de mar. En la Figura 5, se observan las estaciones donde fueron capturadas las larvas.

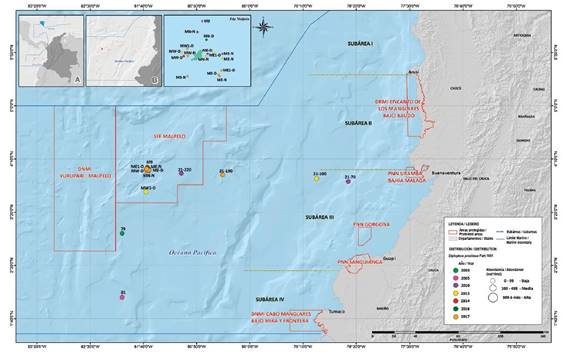
Figura 5 Distribución y abundancia de larvas de Diplophos proximus colectadas entre 2003 y 2017 en el Pacífico colombiano.
La identificación de las larvas de D. proximus se realizó tomando como base los caracteres determinantes de merística, morfometría y pigmentación, establecidos en el Atlas No. 33 de CALCOFI (Moser, 1996). Para determinar la abundancia se consideró el número de larvas y post larvas capturadas por familia, tipo, género, especie y se estandarizaron los resultados a número de individuos por 10 m2, para el análisis se siguió el rango establecido por Rueda y Beltrán (1992): Baja: 1 - 99 individuos/10m2; Media: 100 - 499 individuos/10m2; Alta: > 500 individuos/10m2. El análisis global de la información permitió elaborar dibujos con una cámara lúcida Wild, de cada uno de los estadios de la especie donde se presentan las diferentes características asociadas a una talla determinada.
RESULTADOS
Las colectas de las larvas de D. proximus se realizaron durante septiembre 2003, septiembre 2005, julio y diciembre 2010, diciembre 2013, febrero-marzo 2014, junio 2016, junio y septiembre 2017, en las subáreas III y IV (Figura 5).
Peces de Luz
Diplophos proximus Parr, 1931 (GONOSTOMATIDAE)
Morfología. PREFLEXIÓN: cuerpo muy alargado con longitud preanal (LpA) 68% de la longitud estándar (LE); cabeza pequeña (< 20% LE), ojo redondeado y de tamaño moderado (25 -33% longitud de la cabeza (LC)) y boca grande; membrana completa, aletas pectorales presentes, pero en formación. FLEXIÓN: LpA 64% LE; cabeza y ojos pequeños (< 25% LC); aletas dorsales, anal, pectorales y caudal en formación. POSTFLEXIÓN: LpA 62.7% LE; cabeza y ojo similar al anterior; con dentición en ambas mandíbulas, aleta anal formada. TRANSFORMACIÓN: LpA 56-58% LE; cabeza pequeña; solo aletas dorsal, anal y caudal formadas, pliegues transversales a lo largo del intestino grueso, 36 miómeros pre anales, 86 miómeros totales (Figura 6).
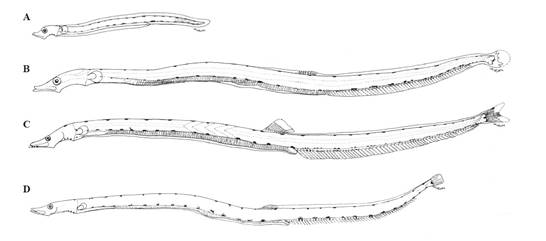
Figura 6 GONOSTOMATIDAE. Diplophos proximus. A. Preflexión 12.7 mm Estación MN-N (1012); B. Flexión 22.5 mm Estación 21-100 (1312); C. Postflexión 26.8 mm Estación 79 (0309); D. Transformación 45.8 mm Estación MW1-D (1312).
Pigmentación. PREFLEXIÓN: En el istmo 2 o 3 pigmentos, en el pedúnculo de la aleta pectoral y sobre el ano, series sobre los bordes dorsal y ventral del cuerpo. FLEXIÓN: En la cabeza, laterales al cerebro y en la punta de la mandíbula. Series de pigmentos post anales en parejas o parches en el borde ventral y base de radios. Abundantes sobre el pedúnculo caudal y radios. POSTFLEXIÓN: Similar a la anterior. Dos sobre el ano. TRANSFORMACIÓN: Similar a la anterior, además sobre el cerebro y maxila, serie grande de melanóforos o parches de pigmentos dorsales y ventrales, 20 dorsal, 13 ventral (en la cola), (Figura 6, Tabla 1).
Tabla 1 Merística y morfometría de larvas de Diplophos proximus capturadas en el Pacífico colombiano entre 2003 y 2017. N: Número de individuos revisados; V: Número de vértebras; M: Número de miómeros; D: Aleta dorsal; A: Aleta Anal; Pv: Aleta Pélvica; Pc: Aleta pectoral; C: Aleta caudal; Rb: Radios branquiostegales; LT: Longitud total; LE: Longitud estándar; *F: Aleta en formación Estación: 1007 (año, mes) ME-N: (Malpelo, Este, noche) MN-N: (Malpelo, Norte, noche); 21-100: (transecto, millas náuticas de la costa), ME1-D: (Malpelo, Este, 1 milla, día). 79: (estación ERFEN), MW-D: (Malpelo, Oeste, día), MW1-D: (Malpelo, Oeste, 1 milla, día).
| ESTADIO Stage | N | V | M | D | A | Pv | Pc | C | Rb | LT | LS | ESTACIÓN Station |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Preflexión / Preflexion | 3 | 86 | - | - | - | - | - | - | 8.3-12.7 | 8.2-12.6 | ME-N 1007; MN-N 1012 | |
| Flexión / Flexion | 2 | 86 | *F | *F | *F | *F | *F | - | 22.5-28.5 | 22.0-27.7 | 21-100 1213; ME1D 1312 | |
| Postflexión / Postflexion | 2 | 9-10 | 57 | *F | *F | *F | 26.8-28.5 | 26.2-27.2 | 79 0309; ME1-D 1312 | |||
| Transformación / Transformation | 2 | - | 10 | 57-59 | *F | *F | 10+9 | - | 41.3-45.8 | 39.5-44.5 | MW-D 1402; MW1-D 1312 | |
Especies similares. Las larvas de Diplophos taenia se pueden distinguir de las de D. proximus, por poseer mayor número de radios anales (59-72), miómeros (89-99) y pigmentos dorsales y ventrales.
Hábitat. Epi y mesopelágicos. Larvas presentes en las subáreas III y IV entre bahía Málaga y límites con Ecuador entre 130 y 463 km de la costa; y al norte, este y oeste del SFF Malpelo entre 0.80 y 1.61 km de la isla (Figura 5).
Condiciones de captura. Temperatura superficial: 25.3-28.1°C (X= 26.8°C). Salinidad superficial: 25-34 (X= 30.7). Profundidad de muestreo: 1-150 m (X= 27.7 m). Distancia de la costa: 130-463 km y 0.9 a 1.85 km de la isla Malpelo. Hora: (03:58 - 20:18) cualquiera, con preferencia por la noche y madrugada.
Abundancia. Baja (≤ 90 ind/m2) en el 100% de las estaciones donde se colectó, estación 79 (22 ind/10m2) septiembre de 2003, estaciones 81 (6 ind/10m2) y M9 (12 ind/10m2) septiembre de 2005, estaciones ME-D (2 ind/10m2) y ME-N (1 ind/10m2) julio de 2010, estaciones 21- 70 (2 ind/10m2) , 21-220 (1 ind/10m2) y MN-N (1 ind/10m2) diciembre de 2010, estaciones 21-100 (2 ind/10m2), ME-N (3 ind/10m2), MN-N (4 ind/10m2), ME1-D (2 ind/10m2), MW-D (4 ind/10m2) y MW1-D (2 ind/10m2) diciembre de 2013, estación MW-D (2 ind/10m2) febrero - marzo de 2014, estación MN-D (4 ind/10m2) junio de 2016, estaciones 21-190 (1 ind/10m2), MS-N (2 ind/10m2), ME-D (2 ind/10m2), Nevera D (2 ind/10m2) junio de 2017 y estaciones ME-N (3 ind/10m2), MW-N (4 ind/10m2), ME1-D (2 ind/10m2) y MW1-D (4 ind/10m2) septiembre de 2017. No se capturaron larvas de esta especie en otros cruceros realizados en fechas diferentes a junio, julio, septiembre, diciembre y febrero- marzo, ni en cruceros realizados a menos de 130 km de la costa. La mayor frecuencia y abundancia se encontró durante los meses de septiembre y diciembre (Figura 5).
Ámbito. Circumglobal en aguas tropicales y templadas, Pacífico oriental tropical al sur de California; siendo este el primer registro para el Pacífico de Colombia. La distribución encontrada para las larvas muestra un ámbito latitudinal entre bahía Málaga (04°00' Lat. N) y la frontera con Ecuador (01°30' Lat. N), subáreas III y IV y longitudinal desde 130 km de la costa (78º27'26,2 W) hasta las aguas circundantes al SFF Malpelo (Figura 5).
DISCUSIÓN
Las colectas de las larvas de D. proximus, se realizaron durante septiembre 2003, septiembre 2005, julio y diciembre 2010, diciembre 2013, febrero-marzo 2014, junio 2016, junio y septiembre 2017, años que coinciden con épocas después de eventos “El Niño” (junio 2002-marzo 2003, agosto 2004-febrero 2005, agosto 2009-mayo 2010, febrero 2015-junio 2016, diciembre 2016-marzo 2017) y durante “La Niña” (agosto 2010-mayo 2011, julio 2016-noviembre 2016, octubre 2017-febrero 2018) (Golden Gate Weather Services, 2015). Para Colombia la especie se encontró en épocas post niño, normales (dic/13 y feb-mar/14) y Niña y post Niña, patrón diferente al registrado por Funes-Rodríguez et al. (2006) para Baja California Sur, que plantea que D. proximus junto con Diaphus pacificus y Benthosema panamense hicieron parte de un grupo de especies tropicales útiles para distinguir el evento El Niño 1982-84. De igual manera, Avendaño-Ibarra et al. (2003) registran otros taxones demersales y mesopelágicos tropicales importantes como Benthosema panamense, Symphurus williamsi, Diplophos proximus y Cubiceps pauciradiatus, que aportaron 12.2% de la abundancia total durante el otoño de 2003 para Baja California Sur, coincidiendo con informes previos de éstos durante periodos cálidos y con anomalías positivas de la temperatura superficial del mar (TSM) (Funes- Rodríguez et al., 1995, 2006).
Diplophos proximus es válida de acuerdo con Ozawa et al. (1990) como endémica en el Pacífico oriental tropical; Fourriére et al. (2016) la registran en los arrecifes del archipiélago de Revillagigedo y áreas cercanas de México. Sin embargo, en muchos casos aún D. proximus es considerado una sinonimia de D. taenia Günther y aunque es una especie diferente, descrita en su estadio larvario en el Atlas No. 33 de CALCOFI (Moser, 1996), esta información no ha sido incluida en la base de peces FishBase.
CONCLUSIONES
Dado que para Colombia la especie Diplophos proximus se encontró durante épocas climáticas normales, ‘El Niño’ y ‘La Niña’ y que para Baja California Sur se le registra durante periodos cálidos y con anomalías positivas, se podría concluir que efectivamente la especie hace parte de un grupo de especies tropicales útiles para distinguir el evento “El Niño” en otras latitudes.
El encontrar estadios larvales de Diplophos proximus desde 2003 hasta la fecha con una distribución relativamente amplia en el centro sur del Pacífico colombiano, permite asumir que en esa región debe existir una población de adultos potencialmente reproductivos de la especie.
El hecho de no tener capturas o registros de adultos de las especies de la familia Gonostomatidae y de muchas otras especies de otras familias de hábitats meso y batipelágicos, motiva a realizar estudios más completos y detallados en aguas profundas colombianas.

