










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
versión impresa ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.47 no.2 Santa Marta jul./dic. 2018
https://doi.org/10.25268/bimc.invemar.2018.47.2.749
Research Articles
Evidence of sexually-produced coral recruitment at Gorgona Island, Eastern Tropical Pacific

1 Grupo de Ecología de Arrecifes Coralinos, Universidad del Valle. Calle 13 # 100-00. Cali, Colombia. carlos.g.munoz@correounivalle.edu.co
2 Instituto de Investigaciones Marinas y Costeras - Invemar [Marine and Coastal Research Institute]. Calle 25 No. 2-55, Playa Salguero. Rodadero, Santa Marta, Colombia.
We present evidence of coral sexual recruitment, which has typically been low in the Eastern Tropical Pacific, on coral reefs and rocky sites of Gorgona Island, Colombia, and for the first time in the Colombian Pacific. With data obtained by thoroughly examining natural substrates, coral juveniles of at least ten species were found at ten out of 19 sites surveyed during scuba diving explorations performed around the island between 2010-2017; however, no corals were found on settlement plates of five different materials. The two most abundant coral species found as juveniles were Porites panamensis and Pocillopora sp., occurring at mean densities of 5.7 ± 4.7 and 0.07 ± 0.11 colonies m-2 (± SD), respectively, and with mean colony-sizes of 2.3 ± 0.9 cm and 3.5 ± 1.0 cm (± SD), respectively. These results indicate that, even though settlement plates do not seem to be as useful to study coral recruitment in the region as they are elsewhere, coral recruitment derived from the sexual reproduction of a diverse set of species is an active process at Gorgona Island.
KEY WORDS: Coral settlement; Sexual recruitment; Juvenile corals; Pocillopora; Porites
Se presenta evidencia de reclutamiento coralino de origen sexual, el cual ha sido típicamente bajo en la región del Pacífico Tropical Oriental, en diferentes arrecifes coralinos y sitios rocosos de isla Gorgona, Colombia, y por primera vez para el Pacífico colombiano.
Mediante un examen minucioso de sustratos naturales, encontramos reclutas de coral de al menos diez especies, en diez de 19 sitios alrededor de la isla durante exploraciones de buceo entre los años 2010 y 2017, pero no se encontraron reclutas adheridos a placas de asentamiento de cinco materiales diferentes instaladas en uno de los arrecifes. Las dos especies de coral con juveniles más abundantes fueron Porites panamensis y Pocillopora sp., con densidades promedio de 5.7 ± 4.7 y 0.07 ± 0.11 colonias m-2 (± DE), respectivamente; y con tamaños promedio de 2.3 ± 0.9 cm y 3.5 ± 1.0 cm (± DE), respectivamente. Estos resultados indican que, aunque las placas de asentamiento no son tan útiles para estudiar el reclutamiento de coral en esta región como sí lo han sido en otros lugares del mundo, el reclutamiento coralino derivado de la reproducción sexual de un conjunto diverso de especies sí es un proceso activo en la isla.
PALABRAS CLAVE: Asentamiento coralino; Reclutamiento sexual; Corales juveniles; Pocillopora; Porites
INTRODUCTION
Recruitment, the incorporation of new individuals to a population, is a key demographic process involved in the maintenance of wild populations (Caley et al., 1996). In the case of organisms like trees and corals, key structural components of forests and coral reefs, recruitment also determines the resilience of these ecosystems (Hughes and Tanner, 2000). Understanding coral recruitment is critical given the evident global deterioration of coral reefs and their associated services in recent decades (Richmond, 1997; Babcock et al., 2003; Hughes et al., 2010).
Coral recruitment has been relatively well documented in the Indo-Pacific, Central Pacific and Caribbean regions, and less studied in the Eastern Tropical Pacific - ETP (Glynn et al., 2017a). Early studies in the ETP did not find coral recruitment on natural substrates, and sampling with settlement plates resulted in extremely low to nil recruitment (Birkeland, 1977; Wellington, 1982; Richmond, 1985; 1987). Such negative results, in addition to those from early histological analyses, even led some researchers to consider ETP corals as “an example of a sterile or expatriate population” (Richmond, 1985, 1987), whose recovery from perturbations was primarily driven by asexual reproduction such as that resulting from colony fragmentation (Highsmith, 1982; Muñoz et al., 2015). More recent studies found few - if any - recruits on settlement plates, for example: nine Porites recruits in the Mexican Pacific (Medina-Rosas et al., 2005), 291 recruits of Porites panamensis but only one Pocillopora recruit also in the Mexican Pacific (López- Pérez et al., 2007), and no recruits at all in Gorgona Island, Colombian Pacific (Lozano-Cortés and Zapata, 2014). Furthermore, new histological evidence clearly indicated that gonadal maturation was an active process (Glynn et al., 1991; 1994; 2017a and references therein), including pocilloporids at Gorgona Island (Castrillón et al., 2015).
Although Gorgona Island is located within a marine protected area so that direct anthropogenic impacts are virtually absent, these reefs are negatively affected by recurring natural disturbances such as El Niño warming events and subaerial exposures during extreme low tides (Zapata and Vargas-Ángel, 2003; Castrillón et al., 2017; Fiedler and Lavín, 2017). For example, mean live coral cover at La Azufrada reef declined 26% in the decade 1998- 2008, and critically so in shallow areas where it decreased from 61% to 15% (Zapata, 2017). Therefore, it has become urgent to know how resilient these reefs are and how much coral recruitment contributes to their resilience. However, one major difficulty to accomplish this has been the lack of evidence showing that sexually-produced coral recruitment actually occurs at Gorgona Island.
In this study, we present evidence of coral recruitment derived from larval settlement on natural substrates in coral reefs and rocky sites at Gorgona Island, but report the absence of coral recruits on settlement plates of different materials. We also report the population densities and size frequency distributions of juvenile colonies of Pocillopora sp. and Porites panamensis.
METHODS
Study area
Gorgona Island, located ~ 30 km off the coast (3° 00' 55''N, 78° 14' 30''W), constitutes the largest insular territory (13.2 km2) in the Colombian Pacific continental shelf. The Island and its surrounding waters are part of Gorgona National Natural Park, which harbors some of the largest, most diverse and developed coral reef formations in the ETP, along with extensive intertidal rocky shorelines and submerged rocky mounts (Zapata and Vargas-Ángel, 2003; Muñoz and Zapata, 2013). To date, around 25 species of hermatypic coral are known from Gorgona Island, but due to taxonomic difficulties within the genus Pocillopora (Schmidt-Roach et al., 2014), the true number remains uncertain (Glynn et al., 2017b). Pocilloporid corals dominate the shallow waters (< 5 m depth) and are the main builders of the coral reefs, while massive-growth species like Pavona varians, Porites lobata and Gardineroseris planulata are common in the fore- reef zones and in deeper rocky sites. Most coral species in Gorgona Island are considered broadcast spawners, except Porites panamensis that is a brooder, and the pocilloporid species with diverse reproductive strategies around the world (Glynn et al., 1991).
The average annual rainfall is high ranging between 4000 - 7000 mm with two contrasting climatological seasons, a rainy one from May to October and a dry season between December and March (Diaz et al., 2001; Blanco, 2009). Due to the heavy rainfall at Gorgona, freshwater streams are abundant, and turbidity is highly variable and partially related to rainfall. Tidal regime is semidiurnal, with a maximum tidal range of 5.7 m (IDEAM, 2017); ocean currents and wave action are stronger on the western, windward side of the main island (Giraldo et al., 2008).
Sampling
Three different methods to examine recruitment of sexually-produced corals at Gorgona Island were implemented: A) a wide search for coral juveniles on natural substrates around the island during exploratory dives, B) visual counts of coral juveniles on transects, and C) the use of settlement plates to sample coral recruits.
Exploratory dives
A total of 19 sites around the island including both coral reefs and rocky sites were inspected during exploratory dives searching for juvenile corals. Minimum three dives at each site were performed per year, 17 sites were surveyed during 2010 - 2015, and two additional sites were surveyed during 2017.
Different criteria (based on Richmond, 1985 and Glynn et al., 1996) were used to establish whether small coral colonies were derived from larval settlement, particularly in the branching pocilloporids that fragment frequently (Highsmith, 1982; Glynn et al., 1991; Muñoz et al., 2015). 1) The presence of a wide, flat, and nearly- symmetrical encrusting base, 2) attachment to steep or vertical substrates like rocky walls, where coral fragments would not be expected to occur, and 3) location of juveniles sufficiently far from adult colonies. Simultaneously fulfilling our criteria for each juvenile provided strong support against criticism by some authors that visual inspection of coral juveniles is insufficient to establish the sexual or asexual origin of a small coral colony (e.g., Miller et al., 2008).
Similar to Edmunds (2004), a criterion to define coral juveniles was a colony-size between 0.1 cm (1 polyp) and ≤ 5.0 cm diameter, hence including both recruits and juvenile corals. Considering that pocilloporid corals achieve sexual maturity at around 2-3 years old and grow ~ 2.7 cm y-1 (Palacios et al., 2014), a colony 5 cm in diameter would be 1.5 years old. Similarly, massive corals like Porites panamensis would be approximately 3.5 years old considering that they reach maturity at 4-7 years old (Richmond, 1997) and grow ~ 1.5 cm y-1 (Cabral- Tena et al., 2013). Finally, pocilloporid juveniles were difficult to identify to species, therefore, were classified as Pocillopora sp.
Counts on transects
To estimate the abundance and size (maximum diameter) of species of coral juveniles, during November 2010 ten visual transects (each transect 20 m2) were examined on each of two coral reefs (La Azufrada and Playa Blanca); during March 2011 nine similar transects were examined at El Laberinto rocky shore; and during June 2011 three smaller transects (10 m2) were examined at El Arrecifito coral reef. To test for abundance and size differences between pocilloporid juveniles from different sites, we used a one-way ANOVA, after testing for compliance with homoscedasticity and normality assumptions.
Settlement plates
In October 2010, 54 settlement plates measuring ~ 20 x 20 x 0.5 cm (~ 840 cm2 total surface per plate) were installed on the reef flat (~ 0.5 to 1 m depth relative to the Mean Lower Low Water datum) of La Azufrada reef (twelve plates of each of four materials: acrylic, ceramic, concrete and marble; and six additional plates of tarred twisted nylon twine interwoven on a polyvinyl pipe frame). The acrylic, ceramic, concrete, and marble plates were arranged in pairs, one on top of the other separated by approximately 2 cm (to provide the cryptic microhabitat often preferred by settling coral larvae (Harriott and Fisk, 1987) and held together by a steel bar driven into the substrate. To reduce potential effects on coral recruits by competition for space with algae and barnacles (Birkeland, 1977), these organisms were removed by scraping the surfaces approximately every 45 days from half of the plates, which were randomly chosen the first time. All plates were photographed, with a high-resolution digital camera, approximately every 45 days for a period of six months and the photographs were examined visually on a computer. In April 2011, the plates were recovered from the sea and examined in the laboratory with a dissecting microscope.
RESULTS
Exploratory dives
We found sexually produced coral juveniles at ten out of a total of 19 sites surveyed at Gorgona Island between 2010 and 2017. Coral juveniles were observed in all the coral reefs and in some of the rocky sites. Los Farallones and El Arrecifito were the sites with most species (6 and 5 species, respectively; Figure 1), while the main coral reefs (La Azufrada, Playa Blanca, La Ventana, El Muelle) had only two species. Coral juveniles belonged to at least ten different species and four families (Table 1; Figure 2).
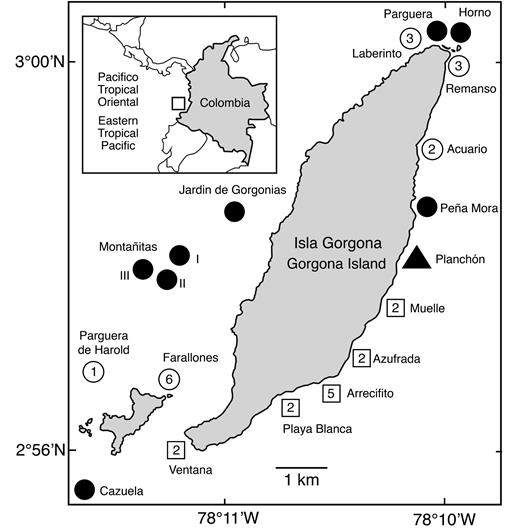
Figure 1 Number of species of juvenile corals per site. Square = coral reef; circle = rocky site; triangle = shipwreck on sandy bottom. Black = no juvenile corals found.
Table 1 Presence of juvenile corals observed at Gorgona National Natural Park (Colombia, Eastern Tropical Pacific) during 2010 - 2017.
| Agariciidae | Pocilloporidae | Poritidae | Psammocoridae | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sitio Site | Ámbito de profundidad (m) Depth Range (m) | Cobertura de coral (~ %) Coral cover (~ %) | Años del estudio Study years | Gardineroseris planulata | Pavona chiriquiensis | Pavona clavus | Pavona frondifera | Pavona gigantea | Pavona varians | Pocillopora sp. | Porites panamensis | Porites lobata | Psammocora stellata |
| Arrecifes de coral Coral Reefs | |||||||||||||
| Arrecifito | 0-10 | 70.0% | 2010-2012 | - | - | - | - | X | X | X | X | - | X |
| Azufrada | 0-10 | 50.0% | 2010-2015 | - | - | - | - | - | X | X | - | - | - |
| Muelle | 0-10 | 70.0% | 2010-2015 | - | - | - | - | X | X | - | - | - | - |
| Playa Blanca | 0-10 | 70.0% | 2010-2015 | - | - | - | - | - | - | X | X | - | - |
| Ventana | 2-10 | 85.0% | 2010-2015 | X | - | - | - | - | - | X | - | - | - |
| Montañas rocosas/peñascos | |||||||||||||
| Rocky mounts / boulders | |||||||||||||
| Cazuela | 16-30 | 0.5% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Farallones | 0-12 | 5.0% | 2010 | - | X | X | X | - | - | X | - | X | X |
| Horno | 0-35 | 0.5% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Jardín de Gorgonias | 10-25 | 1.0% | 2017 | - | - | - | - | - | - | - | - | - | - |
| Parguera de Harold | 10-20 | 3.5% | 2017 | - | - | - | - | - | - | X | - | - | - |
| Montañita I | 7-25 | 0.5% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Montañita II | 3-25 | 0.5% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Montañita III | 20-40 | 0.5% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Peña Mora | 7-20 | 5.0% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Costas rocosas | |||||||||||||
| Rocky shores | |||||||||||||
| Acuario | 0-5 | 40.0% | 2010-2014 | - | - | - | - | X | - | X | - | - | - |
| Laberinto | 0-15 | 25.0% | 2010-2014 | X | - | - | - | - | - | X | - | X | - |
| Parguera | 0-12 | 35.0% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |
| Remanso | 0-12 | 45.0% | 2010-2014 | - | - | - | - | X | - | X | - | X | - |
| Naufragio | |||||||||||||
| Shipwreck | |||||||||||||
| Planchón | 5-17 | 2.0% | 2013-2014 | - | - | - | - | - | - | - | - | - | - |

Figure 2 Coral juveniles derived from sexual reproduction attached to natural substrates at Gorgona Island (Eastern Tropical Pacific). Pocillopora sp. attached to coral rubble with calcareous algae (A, B); Pocillopora sp. attached to rocky wall with filamentous algae (C); Porites cf. panamensis attached to coral rubble with calcareous-algae (D); Pavona varians attached to coral rubble with calcareous-algae (E); Pavona gigantea attached to coral rubble with calcareous-algae (E); Pavona gigantea attached to rocky wall with filamentous algae (F). White arrows point to encrusting base in pocilloporid specimens.
Transect counts
Coral juveniles of at least two species were observed attached to the natural substrates within transects in Gorgona Island. Pocillopora sp. was found on the coral reefs of La Azufrada, and Playa Blanca, as well as at El Laberinto rocky shore (Figures 2A, 2B, 2C; Table 1), while Porites panamensis was found at El Arrecifito coral reef (Figure 2D; Table 1).
We found in total 41 pocilloporid juveniles, and estimated a mean density of 0.07 ± 0.11 colonies m-2 (mean ± SD). The highest coral recruitment was observed at La Azufrada (0.09 ± 0.12 colonies m-2), followed by El Laberinto (0.08 ± 0.11 colonies m-2) and Playa Blanca (0.05 ± 0.08 colonies m-2), but we found no significant differences in the density of pocilloporid juveniles between the three sites (F2, 26 = 0.28, p > 0.1). The mean colony- size of pocilloporid juveniles was 3.5 ± 1.0 cm, also with no significant differences between the three sites (F2, 38 = 0.31; p > 0.1; Figure 3A, 3B, 3C). The smallest pocilloporid juvenile had a diameter ~ 1.0 cm and was observed at El Laberinto attached to a vertical rocky surface (similar to Figure 2C).
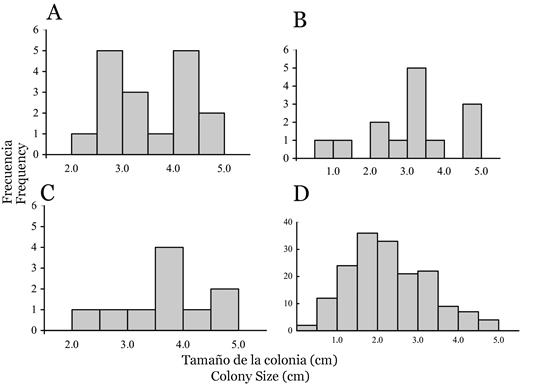
Figure 3 Size frequency distributions of coral juveniles (largest diameter of a colony observed from the top) at Gorgona Island (Eastern Tropical Pacific). The distributions are truncated at 5.0 cm, since juveniles are defined as colonies ≤ 5.0 cm. Distribution for pocilloporid juveniles (N = 41) at three sites: La Azufrada (A), El Laberinto (B), and Playa Blanca (C). Distribution for Porites panamensis juveniles (N = 170) at El Arrecifito (D).
At El Arrecifito, we found a total of 170 juveniles of Porites panamensis with a mean density of 5.7 ± 4.7 colonies m-2 and a mean colony size of 2.3 ± 0.9 cm. The smallest P. panamensis colony was ~ 0.5 cm in diameter, and had between 15 to 20 polyps each one about 0.1 cm in diameter (Figure 2D; 3D).
DISCUSSION
This study reports for the first time the occurrence of coral recruitment of sexual origin in the Colombian Pacific. However, we obtained two conflicting results similar to those reported by previous studies on coral recruitment in the ETP: despite low densities, coral juveniles were found on natural substrates on coral reefs and rocky sites, but the usual sampling method of settlement plates failed to detect coral recruitment (Birkeland, 1977; Wellington, 1982; Richmond, 1985; Medina-Rosas et al, 2005; López-Pérez et al., 2007; Lozano-Cortés and Zapata, 2014). In coral reefs, coral juveniles were found attached either to coral rubble or to the consolidated coral reef matrix covered by calcareous algae, while in rocky sites the juveniles were found attached to rocky walls, either bare or partially covered by mats of filamentous algae.
In addition to other reef locations in the ETP where juveniles of Pocillopora sp. and Porites panamensis derived from sexual reproduction have been already observed on natural substrates (Richmond, 1985; Medina- Rosas et al., 2005; Glynn et al., 1991; 1994; 2017a), this study reports the abundance and size distribution of coral juveniles for these species at the Colombian Pacific. The abundance of coral recruitment on natural substrates in the ETP reefs has been very low, indicating that the abundance reported here for P. panamensis is one of the highest in the region, second to that of Pavona clavus in Costa Rica (Glynn et al., 2017a), and the result of a recruitment pulse and a brooding reproductive mode (Glynn et al., 1994).
The implication of coral recruitment results obtained so far in the ETP, is twofold: first, corals are able of reproducing sexually in the region, previously suggested by several histological studies about coral reproduction throughout the ETP (Glynn et al., 2017a and references therein). These results refute the hypothesis that corals in the ETP are sterile (Richmond, 1985; 1987; Glynn et al., 1991), and at the same time challenge the idea that recruitment derives mostly from fragmentation of colonies (Birkeland, 1977; Highsmith, 1982; Wellington, 1982; Richmond, 1985).
Second, the scarcity of coral recruitment on artificial substrates in the ETP must then be due to some unknown factor that prevents coral larvae from settling or surviving on artificial sampling substrates. The negative results obtained with the settlement plates could be related to: 1) the use of inappropriate substrate materials, 2) insufficient time after deployment for adequate substrate conditioning (López-Pérez et al., 2007), and 3) competition, predation, or herbivory, on settlement plates (Birkeland, 1977; Sammarco, 1982; 1985; Richmond, 1987). However, in this study the first is an unlikely explanation because we used five different materials, including ceramic that has yielded the highest recruitment results in other studies, both in the Indo-Pacific (Harriott and Fisk, 1987) and the Caribbean (Tomascik, 1991). In the second case, it is well known that artificial substrates require a conditioning period during which an encrusting community develops before coral larvae will settle (Segal et al., 2012). While the duration of the conditioning period is not well established and may be highly variable from place to place, in southwestern Mexico coral recruitment occurred 5 - 10 months after plates had been submerged (López-Pérez et al., 2007). Since our plates were submerged for six months, it is plausible that the encrusting community that induces or enhances coral settlement had not fully developed. However, in a previous attempt to document recruitment on artificial substrates at La Azufrada reef, the settlement plates were submerged for a full year and yet no recruits were found (Lozano-Cortés and Zapata, 2014). In the third case, it is known that competition, predation, or herbivory on settlement plates may prevent coral spats from surviving and actually recruiting on these substrates (Birkeland, 1977; Sammarco, 1982; 1985; Díaz-Pulido et al., 2010).
Although we periodically removed algae and other sessile invertebrates from half of the plates to reduce potential competition with recently settled corals, organisms like sea urchins, ophiuroids, and gastropods were commonly seen on the plates and may have interfered with coral settlement or survival.
Additionally, there is a reasonable methodological argument related to the sampling size, whose solution can be impossible in practice: if the values of coral recruitment are naturally low, then the sampling area should be considerably larger than the one we used to successfully detect coral recruits. While there are several other potential explanations, none of them are general enough to explain the common failure to document coral recruitment on artificial settlement plates in the ETP, and therefore the reason for these results remains obscure.
In conclusion, the results presented here indicate that coral recruitment derived from sexual reproduction occurs at Gorgona Island. This, however, has been overlooked by the failure to observe coral settlement on artificial sampling substrates, and the difficulty to differentiate sexual juveniles from small surviving coral fragments. Future research requires examination of the relative contribution of sexual and asexual reproduction to the supply of coral recruitment and their role in the recovery and maintenance of coral populations and coral reefs in the Colombian Pacific and ETP.
ACKNOWLEDGEMENTS
We are thankful to National Natural Parks of Colombia for the constant support to our studies, this research would have been impossible without the help of the staff of Henry von Prahl scientific research station at PNN Gorgona. Main funding provided by COLCIENCIAS and Universidad del Valle (Project code: 1106-489-25135); CGM was beneficiary of the COLCIENCIAS doctorate program.
REFERENCES
Babcock, R., A. Baird, S. Piromvaragorn, D. Thomson and B. Willis. 2003. Identification of scleractinian coral recruits from Indo-Pacific reefs. Zool. Stud., 42: 211-226. [ Links ]
Birkeland, C. 1977. The importance of rate of biomass accumulation in early successional stages of benthic communities to the survival of coral recruits. Proceedings of the Third International Coral Reef Symposium, 1: 15-21. [ Links ]
Blanco, J.F. 2009. The hydroclimatology of Gorgona Island: Seasonal and ENSO-related patterns. Actual. Biol., 31: 111-121. [ Links ]
Cabral-Tena, R.A., H. Reyes-Bonilla, S. Lluch-Cota, D. Paz-García, L. Calderón-Aguilera, O. Norzagaray-Lopez and E. Balari. 2013. Different calcification rates in males and females of coral Porites panamensis in the Gulf of California. Mar. Ecol. Prog. Ser., 476: 1-8. [ Links ]
Caley, M., M. Carr, M. Hixon, T. Hughes, G. Jones and B. Menge. 1996. Recruitment and the local dynamics of open marine populations. An. Rev. Ecol. Syst., 27: 477-500. [ Links ]
Castrillón, A., C. Muñoz and F. Zapata. 2015. Reproductive patterns of the coral Pocillopora damicornis at Gorgona Island, Colombian Pacific Ocean. Mar. Biol. Res., 11: 1065-1075. [ Links ]
Castrillón-Cifuentes, A.L., D.F. Lozano-Cortés and F.A. Zapata. 2017. Effect of short-term subaerial exposure on the cauliflower coral, Pocillopora damicornis, during a simulated extreme low-tide event. Coral Reefs, 36(2): 401-414. [ Links ]
Díaz, J., J. Pinzón, A. Perdomo, L. Barrios y M. López-Victoria. 2001. Generalidades. 17-26. En: Barrios, L. y M. López-Victoria (Eds.). Gorgona marina, contribución al conocimiento de una isla única. INVEMAR, Serie de Publicaciones Especiales No. 7. Santa Marta, Colombia. 160 p. [ Links ]
Diaz-Pulido, G., S. Harii, L. McCook and O. Hoegh-Guldberg. 2010. The impact of benthic algae on the settlement of a reef-building coral. Coral Reefs, 29: 203-208. [ Links ]
Edmunds, P. J. 2004. Juvenile coral population dynamics track rising seawater temperature on a Caribbean reef. Mar. Ecol. Prog. Ser., 269: 111-119. [ Links ]
Fiedler, P. and M. Lavin. 2017. Oceanographic conditions of the Eastern Tropical Pacific. 59-83. In: Glynn P. W., D.P. Manzello and I.C. Enochs (Eds.). Coral Reefs of the Eastern Tropical Pacific: Persistence and Loss in a Dynamic Environment (Coral Reefs of the World, Volume 8). Springer, Dordrecht. 675 p. [ Links ]
Giraldo, A., E. Rodríguez-Rubio y F. Zapata. 2008. Condiciones oceanográficas de Isla Gorgona, Pacifico Oriental Tropical de Colombia. Lat. Am. J. Aquat. Res., 36: 121-128. [ Links ]
Glynn, P. W., J. S. Ault, G. M. Wellington, N. J. Gassman, C. M. Eakin, J. Cortés, D. B. Smith and H. M. Guzmán. 1991. Reef coral reproduction in the eastern Pacific: Costa Rica, Panama and Galápagos Islands (Ecuador). Part I. Pocilloporidae. Mar. Biol., 109: 355-368. [ Links ]
Glynn, P. W., S. B. Colley, C. M. Eakin, D. B. Smith, J. Cortés, N. J. Gassman and J.S. Feingold. 1994. Reef coral reproduction in the eastern Pacific: Costa Rica, Panamá, and Galápagos Islands (Ecuador). II. Poritidae. Mar. Biol., 118(2): 191-208. [ Links ]
Glynn, P., S. Colley, N. Gassman, K. Black, J. Cortés and J. Maté. 1996. Reef coral reproduction in the eastern Pacific: Costa Rica, Panama, and Galapagos Islands (Ecuador). III. Agariciidae (Pavona gigantea and Gardineroseris planulata). Mar. Biol., 125: 579-601. [ Links ]
Glynn, P., S. Colley, E. Carpizo-Ituarte and R. Richmond. 2017a. Coral reproduction in the Eastern Pacific. In: Glynn, P., D. Manzello and I. Enochs (Eds.). Coral Reefs of the Eastern Tropical Pacific: Persistence and loss in a dynamic environment (Coral Reefs of the world, Volume 8). Springer, Dordrecht, Germany. 657 p. [ Links ]
Glynn, P. W., J.J Alvarado, S. Banks, J. Cortés, J.S. Feingold, C. Jiménez… and S. Navarrete. 2017b. Eastern Pacific coral reef provinces, coral community structure and composition: an overview: 107-176. In: Glynn, P., D. Manzello and I. Enochs (Eds.). Coral Reefs of the Eastern Tropical Pacific: Persistence and loss in a dynamic environment (Coral Reefs of the world, Volume 8). Springer, Dordrecht, Germany. 657 p. [ Links ]
Harriott, V. and D. Fisk. 1987. A comparison of settlement plate types for experiments on the recruitment of scleractinian corals. Mar. Ecol. Prog. Ser., 37: 201-208. [ Links ]
Highsmith, R. 1982. Reproduction by fragmentation in corals. Mar. Ecol. Prog. Ser., 7: 207-226. [ Links ]
Hughes, T. and J. Tanner. 2000. Recruitment failure, life histories, and long-term decline of Caribbean corals. Ecology, 81: 2250-2263. [ Links ]
Hughes, T., N. Graham, J. Jackson, P. Mumby and R. Steneck. 2010. Rising to the challenge of sustaining coral reef resilience. Trends Ecol. Evol., 25: 633-642. [ Links ]
López-Pérez, R., M. Mora-Pérez and G. Leyte-Morales. 2007. Coral (Anthozoa: Scleractinia) recruitment at Bahías de Huatulco, Western México: Implications for coral community structure and dynamics. Pac. Sci., 61: 355-369. [ Links ]
Lozano-Cortés, D. and F. Zapata. 2014. Invertebrate colonization on artificial substrates in a coral reef at Gorgona Island, Colombian Pacific Ocean. Rev. Biol. Trop., 62 (Suplem. 1): 161-168. [ Links ]
Medina-Rosas, P., J. Carriquiry y A. Cupul-Magaña. 2005. Reclutamiento de Porites (Scleractinia) sobre sustrato artificial en arrecifes afectados por El Niño 1997-98, en Bahía de Banderas, Pacífico mexicano. Cienc. Mar., 31: 103-109. [ Links ]
Muñoz, C. G. y F. A. Zapata. 2013. Plan de manejo de los arrecifes coralinos del Parque Nacional Natural Gorgona. Parques Nacionales Naturales y WWF- Colombia. Cali, Colombia. 68 p. [ Links ]
Muñoz, C. G., C. Wild and F. A. Zapata. 2015. Drift logs are effective agents of physical coral fragmentation in a tropical eastern Pacific coral reef. B. Mar. Sci., 91(3): 375-376. [ Links ]
Palacios, M. M., C.G. Muñoz and F.A. Zapata. 2014. Fish corallivory on a pocilloporid reef and experimental coral responses to predation. Coral Reefs, 33: 625-636. [ Links ]
Richmond, R. 1985. Variations in the population biology of Pocillopora damicornis across the Pacific. Proceedings of the 5th International Coral Reef Congress, Tahiti, 6: 101-106. [ Links ]
Richmond, R.H. 1987. Energetic relationships and biogeographical differences among fecundity, growth and reproduction in the reefs: coral Pocillopora damicornis. Bull. Mar. Sci., 41(2): 594-604. [ Links ]
Richmond, R. 1997. Reproduction and recruitment in corals: Critical links in the persistence of reefs. 175-197. In: Birkeland, C. (Ed.). Life and Death of Coral Reefs. Chapman and Hall, New York. 536 p. [ Links ]
Sammarco, P.W. 1982. Effects of grazing by Diadema antillarum Philippi (Echinodermata: Echinoidea) on algal diversity and community structure. Journal of Exp. Mar. Biol. and Ecol., 65(1): 83-105. [ Links ]
Sammarco, P. 1985. The Great Barrier Reef vs. the Caribbean: Comparisons of grazers, coral recruitment patterns and reef recovery. Proceedings of the 5th International Coral Reef Congress, Tahiti. 4: 391-397. [ Links ]
Schmidt-Roach, S., K. J. Miller, P. Lundgren, and N. Andreakis. 2014. With eyes wide open: a revision of species within and closely related to the Pocillopora damicornis species complex (Scleractinia; Pocilloporidae) using morphology and genetics. Zool. J. Linnean Soc., 170(1): 1-33. [ Links ]
Segal, B., V. Berenguer and C. Castro. 2012. Experimental recruitment of the Brazilian endemic coral Mussismilia braziliensis and conditioning of settlement plates. Cienc. Mar., 38(1A): 1-10. [ Links ]
Tomascik, T. 1991. Settlement patterns of Caribbean scleractinian corals on artificial substrata along a eutrophication gradient, Barbados, West Indies. Mar. Ecol. Prog. Ser., 77: 261-269. [ Links ]
Wellington, G. 1982. Depth zonation of corals in the Gulf of Panama: Control and facilitation by resident reef fishes. Ecol. Monogr., 52: 224-241. [ Links ]
Williams, D.E., M.W. Miller and K.L. Kramer. 2008. Recruitment failure in Florida Keys Acropora palmata, a threatened Caribbean coral. Coral Reefs, 27(3): 697-705. [ Links ]
Zapata, F. 2017. Temporal dynamics of coral and algal cover and their drivers on a coral reef of Gorgona Island, Colombia (Eastern Tropical Pacific). Rev. Acad. Col. Cienc. Ex. Fís. Nat., 41(160): 298-310. [ Links ]
Zapata, F. and B. Vargas-Ángel. 2003. Corals and coral reefs of the Pacific coast of Colombia. 419-447. In: Cortés, J. (Ed.). Latin American Coral Reefs. Elsevier Science B.V., Amsterdam. 508 p. [ Links ]
Received: February 01, 2018; Accepted: August 29, 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
